Medical Diagnosis
Vol.2 No.2(2012), Article ID:656,5 pages DOI:10.12677/md.2012.22002
The Molecular Cloning and Bioactivity Characterization of Calmodulin from Medicinal Fungus Agrocybe aegerita
1Department of Clinical Immunology, Guangdong Medical College, Dongguan
2The College of Life Sciences, Wuhan University, Wuhan
Email: liangyi@gdmc.edu.cn
Received: Mar. 29th, 2012; revised: Apr. 9th, 2012; accepted: Apr. 24th, 2012
ABSTRACT:
Objective: The antitumor-associated gene of Agrocybe aegerita was cloned to identify the bioactivity. Method: cDNA library was constructed and the gene calmodulin was obtained by random sequencing. The eukaryotic expression vector was constructed to detect cell apoptosis and senescence activity. Result: The calmodulin gene was cloned with 588 bps code for 150 amino acid residues, and the amino acid sequence was very conserved in calmodulin family. The expression of cloned gene in HeLa cells did not induce the cell apoptosis or senescence. Conclusion: The sequence of calmodulin contained conserved motif and belonged to calmodulin family.
Keywords: Medicinal Fungus; CaM; cDNA Sequence
药用真菌杨树菇钙调素基因及活性初步鉴定
梁 一1,孙 慧2
1广东医学院检验学院,临床免疫教研室,东莞
2武汉大学生命科学学院,武汉
Email: liangyi@gdmc.edu.cn
摘 要:
目的:克隆杨树菇抗肿瘤相关蛋白基因,初步鉴定其活性。方法:构建cDNA文库,随机测序得到杨树菇钙调素(calmodulin,CaM)基因,并对其进行生物信息学分析,通过检测其在HeLa细胞中表达引起的细胞凋亡及衰老的变化,初步鉴定其抗肿瘤活性。结果:得到全长为588 bp的杨树菇钙调素cDNA序列,编码150个氨基酸,其氨基酸序列在整个钙调素家族中十分保守。钙调素基因在真核细胞中表达没有引起肿瘤细胞的凋亡和衰老。结论:杨树菇钙调素序列具有CaM保守序列模式,是CaM家族的成员。
收稿日期:2012年3月29日;修回日期:2012年4月9日;录用日期:2012年4月24日
关键词:药用真菌;CaM;cDNA序列
1. 引言
药用真菌因其独特的药理活性被越来越多的研究者关注[1-3]。其中蛋白质组分占子实体干重约10%~ 40%[4],但长期以来,仅有上百种蛋白/多肽被分离,且多集中在对其氨基酸组成、活性分析等初步研究。已有十多种药用真菌蛋白被报道具很强的抗肿瘤活性(nM级)[5,6],提示除多糖外,蛋白质是药用真菌中另一类重要的抗肿瘤活性组分[7]。
然而由于药用真菌基因组信息缺乏,不易克隆蛋白基因序列,药用真菌的抗肿瘤活性蛋白研究十分不足。目前随着高通量测序技术的成熟,国内外学者也都致力于对药用真菌基因组的测序工作,极大地推动了药用真菌活性蛋白的研究。在之前的研究中,杨树菇Agrocybe aegerita活性蛋白在体内和体外试验中均表现出显著的抗肿瘤活性[7],本文通过构建cDNA文库测序的方法,克隆药用真菌杨树菇抗肿瘤活性相关基因并对其活性做初步筛选,为研究抗肿瘤蛋白提供理论基础。
2. 材料与方法
2.1. 材料
杨树菇Agrocybe aegerita子实体由华中农业大学食用菌研究所培育,品种为杨树菇3号,采集之后迅速冷冻于液氮罐中保存。RNA提取试剂盒是Qiagen公司的RNeasy Plant Mini Kit,SMART cDNA Library Construction Kit购于Clontech公司。真核表达载体pGEFP-C1系本实验室保存,HeLa细胞购于中国典型培养物保藏中心(CCTCC),转染试剂为invitrogen公司的Lipofectamine 2000。衰老相关β-galactosidase检测试剂盒购自上海杰美生物公司,碘化丙锭PI购于武汉大风生物技术有限公司。
2.2. cDNA文库及基因测序
使用RNeasy Plant Mini Kit提取杨树菇子实体总RNA,按照SMART cDNA Library Construction Kit说明书构建cDNA文库。即在合成cDNA的反应中事先加入的3’末端带Oligo(dG)的SMART引物,由于逆转录酶以mRNA为模板合成cDNA,在到达mRNA的5’末端时碰到真核mRNA特有的“帽子结构”,即甲基化的G时会连续在合成的cDNA末端加上几个(dC),SMART引物的Oligo(dG)与合成cDNA末端突出的几个C配对后形成cDNA的延伸模板,逆转录酶会自动转换模板,以SMART引物作为延伸模板继续延伸cDNA单链直到引物的末端,这样得到的所有cDNA单链的一端有含Oligo(dT)的起始引物序列,另一端有已知的SMART引物序列,合成第二链后可以利用通用引物进行扩增。由于有5’帽子结构的mRNA才能利用这个反应得到能扩增的cDNA,因此扩增得到的cDNA就是全长cDNA。
对文库进行随机测序,得到的序列用CodonCode Aligner软件去掉低质量序列、载体序列、重复序列等,用Phrap拼接同源EST,并与NCBI非冗余蛋白库比对(BLASTX)。
2.3. 真核表达质粒构建及转染
通过引物Forward: cccaagcttggatggctgaccaactg, reverse: cggggtaccttactttgacaacatcatc,将CaM基因克隆到真核表达载体pGEFP-C1上。HeLa细胞按每孔约3 × 104个细胞接种于6孔细胞培养板,37℃培养过夜,待细胞密度为80%~90%使用Lipofectamine 2000转染真核表达质粒。
2.4. PI染色及流式细胞仪分析
按照上述方法转染待质粒,继续培养48 h。使用荧光显微镜检测转染效率。细胞用PBS重悬洗涤1次,75%乙醇重悬后,加RNase A (终浓度0.25 mg/mL) 37℃孵育1 h,终浓度50 mg/mL的碘化丙啶4℃避光染色30 min,流式细胞计数仪检测荧光强度分布(激发波长488 nm)。
2.5. 衰老相关β-galactosidase检测
按照上述方法转染质粒,使用β-galactosidase衰老相关试剂盒里对细胞进行染色,细胞呈现蓝色部位为衰老细胞特异性酶(β-半乳糖苷酶)活性位点。
2.6. 统计学方法
应用统计软件SPSS13.0进行数据分析,组间比较采用单因素方差分析,两两比较采用LSD-t检验,P < 0.05有统计学意义。
3. 结果
3.1. 钙调素基因序列及数据分析
杨树菇cDNA文库构建成功,随机测序获得杨树菇Agrocybe aegerita钙调素序列(如图1所示),长588 bp。用NCBI的ORF Finder程序分析其开放阅读框,5’-非翻译区为52 bp,3’-非翻译区86 bp,其中末尾有31 bp的poly-A。含有长450 bp的开放阅读框(包括一个终止密码子),编码150个氨基酸。
杨树菇钙调素氨基酸序列用BLASTP搜索找出其相似度较高的氨基酸序列,然后用Clustal W2比对序列,如图2所示。Agrocybe aegerita CaM与Schizophyllum commune CaM(登录号XP_003025993)、Ganoderma lucidum CaM(登录号ADV17372)、Postia placenta CaM(登录号XP_002473838)和Homo sapiens

Figure 1. The gene of calmodulin from Agrocybe aegerita and the deduced amino acid sequence
图1. 杨树菇Agrocybe aegerita calmodulin基因序列及氨基酸序列

Figure 2. The amino acid sequence comparison of calmodulin. “*” meant identical residues, “:” meant conserved substitutions, “.” meant semiconserved substitutions
图2. 钙调素氨基酸序列比对结果。“*”表示完全相同的氨基酸残基,“:”表示保守替换,“.”表示非保守替换
CaM(登录号NP_001734)的相似性分别为96.7%、96.7%、94.7%和91.3%,这一结果表明钙调素氨基酸序列不仅在真菌中非常保守,在整个钙调素家族中都十分保守。
通过SWISS-MODEL[8]对杨树菇钙调素进行结构预测,同源建模的模板为Gallus gallus的钙调素分子(PDB号:2o5g),如图3所示,杨树菇钙调素分子由8个α螺旋和3个β折叠组成,属典型的α + β类型结构。结合钙离子的4个结构片段与模板相比没有显著变化,进一步说明该钙调素分子的结构保守性。

Figure 3. In silico model of calmodulin. (a) The experimentally determined structure of Agrocybe aegerita calmodulin; (b) Schematic representation of the molecular models of Gallus gallus calmodulin (PDB code: 2o5g)
图3. 钙调素基因同源建模:(a) 同源建模得到的杨树菇钙调素蛋白结构;(b) SWISS-MODEL中同源建模的模板,为Gallus gallus钙调素分子(PDB号:2o5g),灰色小球代表钙离子
3.2. 真核表达质粒构建及转染
PCR扩增连在cDNA文库构建质粒载体pDNRlib上的CaM基因,扩增结果如图4(a),将基因克隆到pEGFP-C1载体上,酶切鉴定结果如图4(b)。真核表达载体构建完成后,用pEGFP-C1空载体作为对照,以相同的量转染入6孔板的HeLa细胞,48小时后通过标签GFP在荧光显微镜下检测转染效率,图4(c)表明转染效率可达80%以上。
3.3. 钙调素表达对HeLa细胞的影响
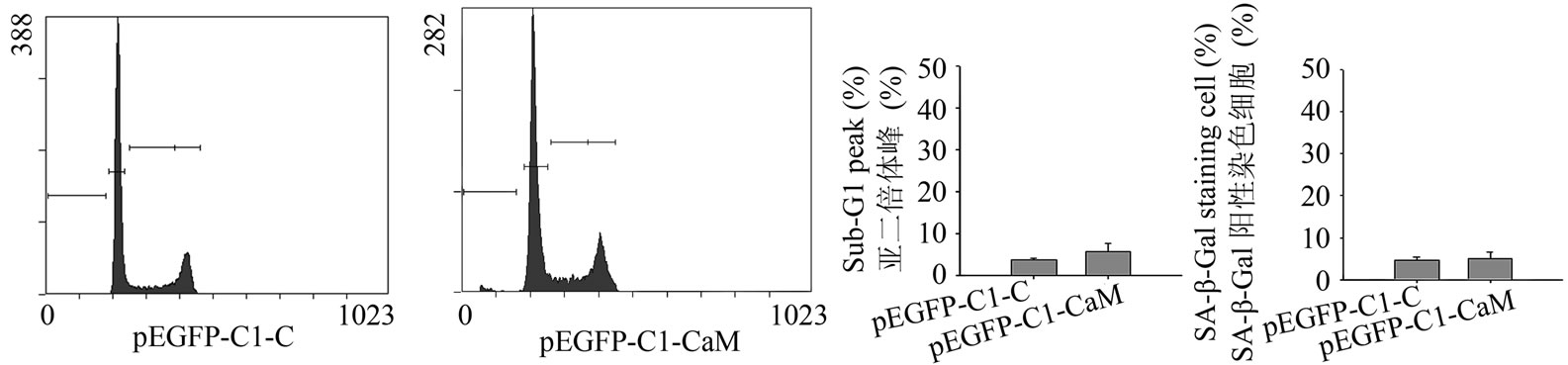
使用碘化丙啶染色以及流式细胞仪分析发现,CaM基因表达后,HeLa细胞亚二倍体峰细胞群也就是凋亡细胞数目并没有增多,如图5(a)所示。转染空质粒凋亡峰为3.86% ± 0.32%,CaM基因表达诱导细胞出现5.8% ± 1.9%的亚二倍体峰,与对照组相比没有显著性差异(图5(b))。
转染质粒48小时后,使用衰老相关β-galactosidase检测试剂盒对细胞进行染色,每个样品随机选取4个视野,计算SA-β-Gal呈阳性反应的细胞比例,统计结果如图5(c)所示,转染CaM基因与对照组相比没有显著性差异。
4. 讨论
钙调素是细胞中重要的功能分子,参与介导许多生理活动,包括炎症反应、代谢、细胞凋亡、神经发育、免疫反应等[9]。本研究得到杨树菇Agrocybe aegerita钙调素基因,不仅与真菌钙调素同源性高,与人类钙调素序列比对同源性也很高,这说明钙调素分子序列在整个钙调素家族中都十分保守,也间接说明该分子在细胞功能的重要性。
 (a) (b) (c)
(a) (b) (c)
Figure 4. The construction and the transformation of the plasmid pEGFP-C1-CaM. (a) The PCR result of calmodulin, M: DL2000 Marker, 1: Amplification of calmodulin’s ORF; (b) M: DL2000 Marker, 1: the result of enzyme digestion of the plasmid pEGFP-C1-calmodulin; (c) The transformation of the plasmid pEGFP-C1-calmodulin. The plasmid with GFP-tag was transformed into HeLa cells, and after 48h the result was observed and pictured by fluorescence microscope (pEGFP-C1-C was the control)
图4. pEGFP-C1-CaM真核质粒构建及转染:(a) 钙调素基因扩增,M:DL2000 Marker,1为钙调素ORF扩增结果;(b) M:DL2000 Marker,1为pEGFP-C1-calmodulin质粒酶切鉴定结果;(c) pEGFP-C1-CaM质粒转染,带有GFP标签的质粒转染HeLa细胞,48小时后荧光显微镜观察并照相(pEGFP-C1-C为空载体对照)
 (a) (b) (c)
(a) (b) (c)
Figure 5. The effect of the expression of calmodulin on HeLa cells. (a) The plasmids were transformed into HeLa cells, and after 48 h, the cells were detected by PI staining and flow cytometry. pEGFP-C1-C was used as blank control, and the line on the left indicated sub-G1 peak; (b) The statistical data of three experiments performed in A; (c) The cells was stained for 16 h by the senescence associated β-galactosidase detection kit, and the percentage of positive cells was statistically collected in the four microscopic fields random selected
图5. 检测钙调素基因表达对HeLa细胞的影响:(a) 转染质粒48小时后,PI染色结合流式细胞仪检测细胞。pEGFP-C1-C为转入空质粒对照,图中左侧横线表示亚二倍体峰;(b) 三次流式试验的统计学数据;(c) 使用衰老相关β-galactosidase检测试剂盒染色16小时,随机挑选4个视野计数阳性反应即衰老细胞比例的统计数据
药用真菌中蛋白组分的研究十分不足,重要原因之一在于其基因信息的缺乏。由于其繁殖特点(有性或无性繁殖,菌丝的异宗结合,不同遗传背景的菌丝纽结成子实体等特点),使得基因克隆非常困难,因此,构建cDNA文库结合表达序列标签分析将有助于获得大量基因信息,从而推动对药用真菌抗肿瘤蛋白的研究。本研究通过该方法结合构建真核表达质粒,转染入HeLa细胞,初步筛选到多个具有抗肿瘤活性的基因,包括泛素蛋白酶系统相关分子,及部分未知基因。真菌钙调素功能及作用机制各有不同,例如病原真菌的钙调素分子可能参与帮助病原体存活[10]。药用真菌的钙调素分子研究不多,基于药用真菌独特的抗肿瘤作用,本文检测了真核表达杨树菇钙调素的活性,发现并没有引起肿瘤细胞的凋亡或衰老,该结果可能与钙调素基因在整个家族中高度同源有关,至于杨树菇钙调素的其他生物学功能需要在进一步的研究中做一阐明。
5. 致谢
感谢国家自然科学基金(No. 81102850),广东省医学科研基金(A2011434),东莞市高等院校科研项目基金(No. 2011108102049)及广东医学院博士启动基金项目(XB1106)给予本研究的支持。
参考文献 (References)
[1] A. T. Borchers, C. L. Keen and M. E. Gershwin. Mushrooms, tumors, and immunity: An update. Experimental Biology and Medicine, 2004, 229(5): 393-406.
[2] A. T. Borchers, A. Krishnamurthy, C. L. Keen, et al. The immunobiology of mushrooms. Experimental Biology and Medicine, 2008, 233(3): 259-276.
[3] A. T. Borchers, J. S. Stern, R. M. Hackman, et al. Mushrooms, tumors, and immunity. Proceedings of the Society for Experimental Biology and Medicine, 1999, 221(4): 281-293.
[4] W. Breene. Nutritional and medicinal value of specialty mushrooms. Journal of Food Protection, 1990, 53(10): 883-894.
[5] T. B. Ng. Peptides and proteins from fungi. Peptides, 2004, 25 (6): 1055-1073.
[6] N. T. Wang, H. X. Ool. V.E.C, Lectins from mushrooms. Mycological Research, 1998, 102(8): 897-906.
[7] C. Y. Liang, H. H. Liu, R. Luan, T. Che, S. Jiang, D. Y. Xie and H. Sun. The tumor rejection effect of protein components from medicinal fungus. Biomedicine & Preventive Nutrition, 2011, 1 (4): 245-254.
[8] K. Arnold, L. Bordoli, J. Kopp, et al. The SWISS-MODEL workspace: A web-based environment for protein structure homology modeling. Bioinformatics, 2006, 22(2): 195-201.
[9] M. J. de Carvalho, R. S. A. Jesuino, B. S. Daher, et al. Functional and genetic characterization of calmodulin from the dimorphic and pathogenic fungus Paracoccidioides brasiliensis. Fungal Genetics Biology, 2003, 39(3): 204-210.
[10] P. R. Kraus, J. Heitman. Coping with stress: Calmodulin and calcineurin in model and pathogenic fungi. Biochemical and Biophysical Research Communications, 2003, 311(4): 1151-1157.