Hans Journal of Agricultural Sciences
Vol.1 No.2(2011), Article ID:398,7 pages DOI:10.4236/hjas.2011.12002
Genetic Differentiation of Hybrids from Cultivated Strain and Wild Strain in Oyster Mushroom Pleurotus ostreatus*
Institute of Applied Mycology, Huazhong Agricultural University, Wuhan
Email: bianyb.123@163.com
Received Sep. 15th, 2011; revised Oct. 20th, 2011; accepted Oct. 24th, 2011.
ABSTRACT:
Cultivated strain HP and wild strain ZJJ of Pleurotus ostreatus were used in this study. 38 monokaryons from strain HP and 55 monokaryons from strain ZJJ were obtained respectively by single basidiospore isolation. 10 monokaryons from each parental strain were selected randomly for hybridization and 14 dikaryons were obtained. According to the different antagonist reaction between hybrid and parents, the 14 hybrids were divided into 4 types. The mycelia growth rate on different media showed dominant differrentiation. The result of cultivation test showed that one hybrid could not fruit and yield of the others were lower than that of the cultivated strain HP, while the yields of 5 hybrids were higher than that of the wild strain ZJJ. Among the hybrids, most pilei showed similar color with that of parent HP (white), 3 hybrids formed fruitbodies without pilei. The analysis of hybrids agronomic charateristics showed that the coefficient of variation (CV) of yield was the largest (21.9%), the others in the order were primordial initiation days (25.4%), stipe length (21.8%),full colonization time (21.0%),average weight of single fruitbody (16.2%), pileus thickness (14.5%), pileus diameter (13.5%)and stipe diameter (9.4%).
Keywords: Hybridization; Trait; Genetic Differentiation
平菇栽培菌株与野生菌株杂交子农艺性状的遗传分化*
张玉霞,徐章逸,蔡英丽,边银丙
华中农业大学应用真菌研究所,武汉
Email: bianyb.123@163.com
摘 要:
以平菇(Pleurotus ostreatus)栽培菌株HP和野生菌株ZJJ为杂交亲本材料,采用担孢子稀释分离法分别获得HP、ZJJ的单孢菌株38个和55个。两个亲本各随机挑选10个单孢菌株在PDA培养基上进行配对杂交,获得14个杂交子。根据杂交子与两个亲本菌株的拮抗反应不同,可将14个杂交子划分为4种类型。在不同的培养基质中,菌丝生长速度出现明显分化。杂交后代中除1个未出菇之外,其它杂交子的平均产量都低于栽培菌株HP,但有5个杂交子产量高于野生菌株ZJJ。大部分杂交子菌盖颜色接近亲本HP的颜色,其中3个杂交子长出无菌盖的子实体。杂交子农艺性状分析表明,产量的变异系数最大(37.2%),其它依次为原基形成期(25.4%)、菌柄长度(21.8%)、菌丝长满袋的天数(21.0%)、单菇重量(16.2%)、菌盖厚度(14.5%)、菌盖直径(13.5%)和菌柄直径(9.4%)。
收稿日期:2011年9月15日;修回日期:2011年10月20日;录用日期:2011年10月24日
关键词:杂交;性状;遗传分化
1. 引言
狭义上平菇是糙皮侧耳(Pleurotus ostreatus)的商品名,广义上的平菇则包括糙皮侧耳(Pleurotus ostreatus)、肺形侧耳(P. pulmonarius)、紫孢侧耳(P. sapidus)、黄白侧耳(P. cornucopiae)等生产中广泛栽培的侧耳属(Pleurotus)多个种类,它们隶属于担子菌门(Basidiomycota)、层菌纲(Hymenomycetes)、伞菌目(Agaricales)、侧耳科(Pleurotaceae)。平菇是广泛栽培的食用菌之一,适应性强,分布广泛,是具有重要食用和药用价值的木腐真菌(Thorn et al., 2000),在木质纤维素的转化以及环境修复方面也有很重要的作用(Hadar et al., 1993),其产生大量的孢子会严重影响菇农的身体健康(Helbling A. et al.,1998),选育具有优良农艺性状以及少孢或无孢品种是育种工作的主要目标。本研究以平菇栽培菌株HP和野生菌株ZJJ为亲本,分析了F1代杂交子农艺性状的遗传分化,为平菇优良杂交子系统筛选奠定了基础。
2. 材料和方法
2.1. 供试菌株及来源
华平特白HP系平菇生产中广泛使用的栽培菌株,由华中农业大学菌种实验中心提供;野生菌株ZJJ由浙江省丽水林业科学院应国华提供,系从湖南张家界国家自然保护区野外采集的子实体经组织分离获得(图1)。
2.2. 供试培养基
PDA培养基:用于菌丝培养(戚元成等,2010)。
三级菌种培养料配方:棉籽壳80%,麸皮18%,石膏1%,石灰1%。
栽培料配方:棉籽壳90%,麸皮8%,石膏1%,石灰1%。
2.3. 杂交子的获得
亲本菌株HP和ZJJ的栽培、单孢菌株的获得以及杂交配对均参考文献(Jatinder Kaur et al., 2008)进行,以锁状联合的有无来判定是否形成杂交子。

Figure 1. Fruit bodies of crossing parental strains
图1. 杂交亲本菌株的子实体
2.4. 拮抗线试验与菌丝生长速度
使用直径90 mm的平皿,每皿倒约25 mL PDA培养基。将杂交菌株和两亲菌株的接种块等距离接种于同一平皿,接种点之间间距约4 cm,于25℃培养箱倒置培养20 d,观察拮抗线形成情况。
参考文献(李红梅等,2009)中的方法测定菌丝在不同培养基上的生长速度,每个处理设3次重复。采用软件spss17.0对生长速度数据进行分析。
2.5. 栽培试验与农艺性状观察
试验于2010年9月初制备栽培种,10月1日在武汉市洪山区野芷湖村进行接种栽培,每个菌株接种15袋,随机区组试验设计,在子实体菌盖伸展但边缘内卷(约8分成熟)时采收。对亲本及其杂交后代的菌丝满袋时间、原基形成期、子实体形态、产量等进行了观察记录。每个菌株随机选取达到采收标准的15个子实体,测量单菇重量、菌盖直径、菌柄长、菌柄直径和菌盖厚度。采用数据处理软件spss17.0对农艺性状相关的数据进行统计分析。
3. 结果与分析
3.1. 单核体杂交配对结果
以锁状联合为标记,从HP、ZJJ两个亲本菌株子实体上经过单孢分离培养,分别获得38个和55个单孢菌株。每个亲本随机取10个单孢,分别编号为H1, H2, ... H10和Z1, Z2, ... Z10,进行10 × 10配对杂交。在100个配对杂交组合中,形成了14个杂交子,杂交亲和率为14%。
3.2. 杂交子与亲本的拮抗试验
将14个杂交子与两个亲本进行对峙培养,PDA培养基中培养20d左右,可观察到杂交子与两亲本间都出现拮抗线。根据拮抗线区域菌丝的密度、有无色素沉着等特征,可将拮抗线分为a、b、c三种类型。a型是在两菌落接触区出现菌丝集结隆起,并有明显的色素沉着的分界线;b型既无明显的菌丝集结隆起,又无色素产生;c型出现菌丝集结隆起,但拮抗线颜色较浅。
根据拮抗线类型的不同,可将杂交子分为HA、HB、HC和HD四种类型(图2)。HA型杂交子与两亲本均形成a型拮抗线,包括杂交子H2Z1、H2Z2、H2Z4 、H2Z5、H2Z6、H2Z8和H2Z10;HB 型杂交子与亲本HP形成a型拮抗线,与亲本ZJJ形成b型拮抗线,包括杂交子H5Z1、H5Z7和H5Z10;HC型杂交子与两亲本均形成b型拮抗线,仅包括杂交子H5Z2;HD型杂交子与亲本HP形成a型拮抗线,与亲本ZJJ形成c型拮抗线,包括杂交子H2Z3、H2Z7和H2Z9。
3.3. 杂交子的菌落形态和菌丝生长速度
杂交子在培养皿上呈现8种不同的菌落形态,气生菌丝形态、色泽、浓密程度、环形轮纹有无等方面存在明显不同(图3)。与亲本菌落特征相比较,杂交群体出现显著的性状分离,表现出不同于亲本菌株的特征。
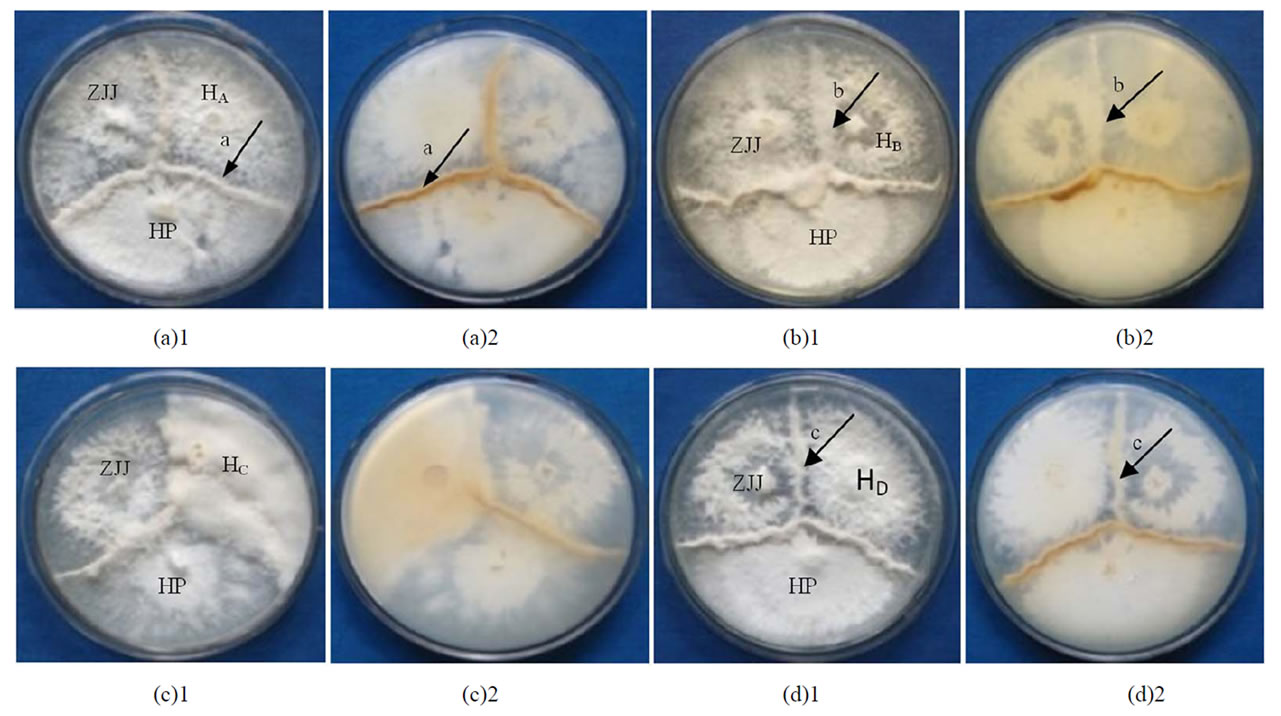
HA、HB、HC and HD represent respectively the hybrids of four antagonist reaction types.
HA、HB、HC和HD 代表4种拮抗反应类型的杂交子。
Figure 2. Antagonist reaction types between hybrid dikaryons and parent strains
图2. 平菇杂交子与亲本的拮抗反应类型

Figure 3. Morphological characteristics of the Colony of hybrids and its parents HP and Zjj in Pleurotus ostreatus
图3. 平菇HP × ZJJ杂交子及亲本的菌落形态
所有杂交子在PDA培养基上菌丝生长速度均显著慢于亲本HP,12个杂交子生长速度介于两亲本之间,H5Z1与H5Z2两个杂交子生长速度慢于亲本ZJJ。4个杂交子在三级菌种培养基中菌丝生长速度超过两个亲本,3个杂交子介于两个亲本之间,7个杂交子生长速度慢于两个亲本。杂交子在PDA培养基与三级菌种培养基中菌丝生长速度表现并不一致(图4)。
3.4. 杂交子的农艺性状
对出菇期子实体形态进行观察,8个杂交子菌盖颜色为白色,与栽培菌株HP相似, 但H5Z2、H5Z7、
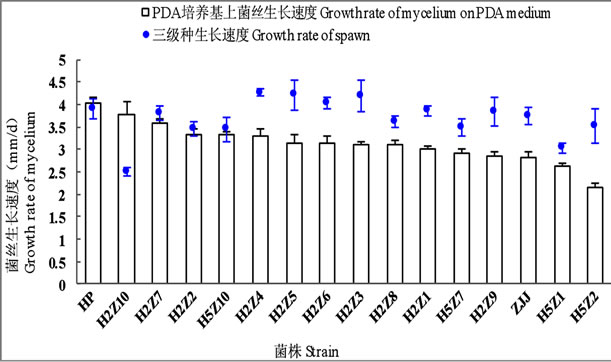
Figure 4. Mycelial growth rate of hybrids on PDA medium and spawn medium in Pleurotus ostreatus
图4. 平菇杂交子在PDA培养基和三级菌种培养料中的菌丝生长速度
H5Z10等3个菌株形成无菌盖的子实体, H2Z2菌盖表面有密集的瘤状突起(图5)。
在菌丝满袋时间上,6个杂交子菌丝满袋时间较两亲本时间短,6个杂交子较两个亲本菌株时间长。9个菌株原基形成时间短于两亲本(H2Z1~H2Z9),3个菌株原基形成时间较两个亲本菌株长,H5Z1未形成原基。在产量性状上,H2Z5、H2Z6、H2Z9等3个杂交子产量介于两亲本之间,H5Z2、H5Z7、H5Z10等3个菌株产量极低。H2Z5、H2Z10两个杂交子单菇重量高于两亲本,且差异显著,其它杂交子的单菇重量均低于两个亲本。H2Z4、H2Z6、H2Z8等3个杂交子菌柄较亲本HP短,且H2Z4和H2Z8菌柄长度与亲本差异显著。5个杂交子菌柄直径介于两亲本之间,3个杂交子菌柄直径高于亲本HP。在菌盖直径上,1个杂交子超过2个亲本,4个杂交子介于两个亲本之间,5个杂交子菌盖直径小于两亲本,3个杂交子未形成菌盖;3个杂交子菌盖厚度大于两亲本,3个杂交子小于两个亲本,4个杂交子介于两个亲本之间(表1)。
对杂交子农艺性状特征指标进行分析表明(表2),单袋产量的变异系数最大(37.2%),其它依次为原基形成期(25.4%)、菌柄长度(21.8%)、满袋天数(21.0%)、单菇重量(16.2%)、菌盖厚度(14.5%)、菌盖直径(13.5%)
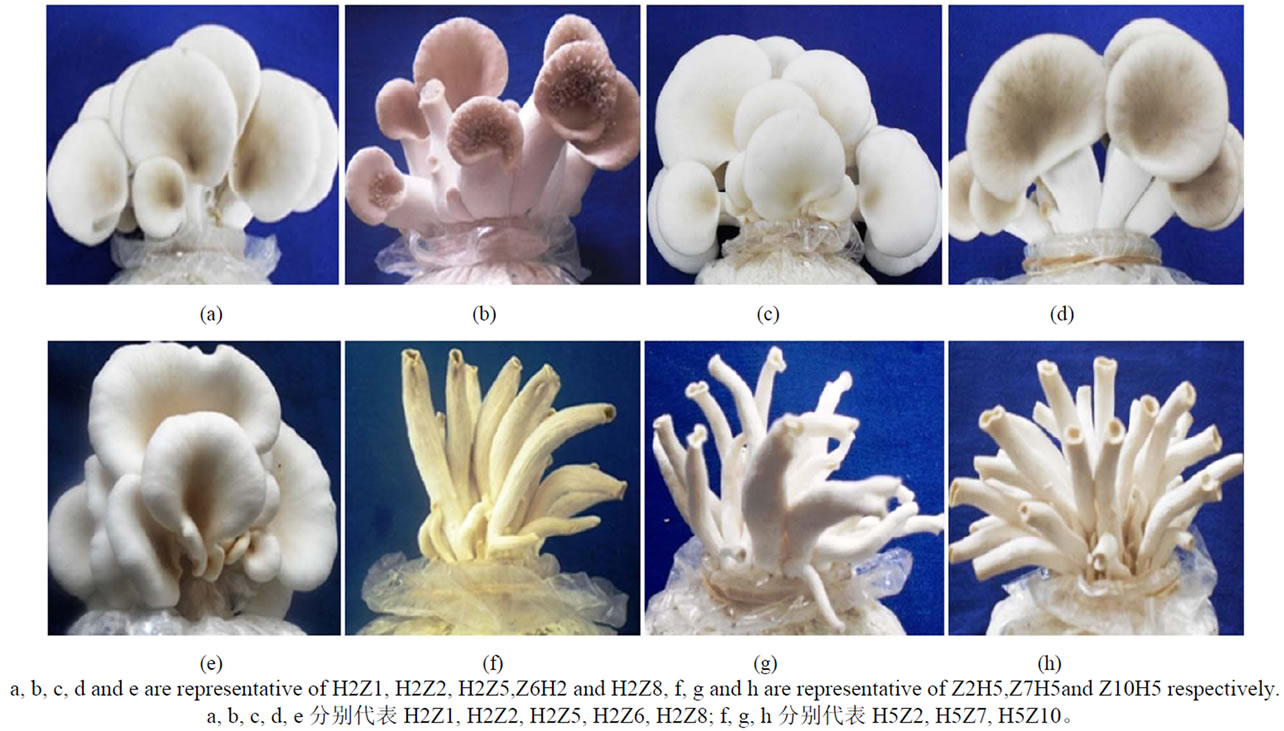
Figure 5. Different fruitbodies morphologicals of some hybrids in Pleurotus ostreatus
图5.平菇部分杂交子的子实体形态

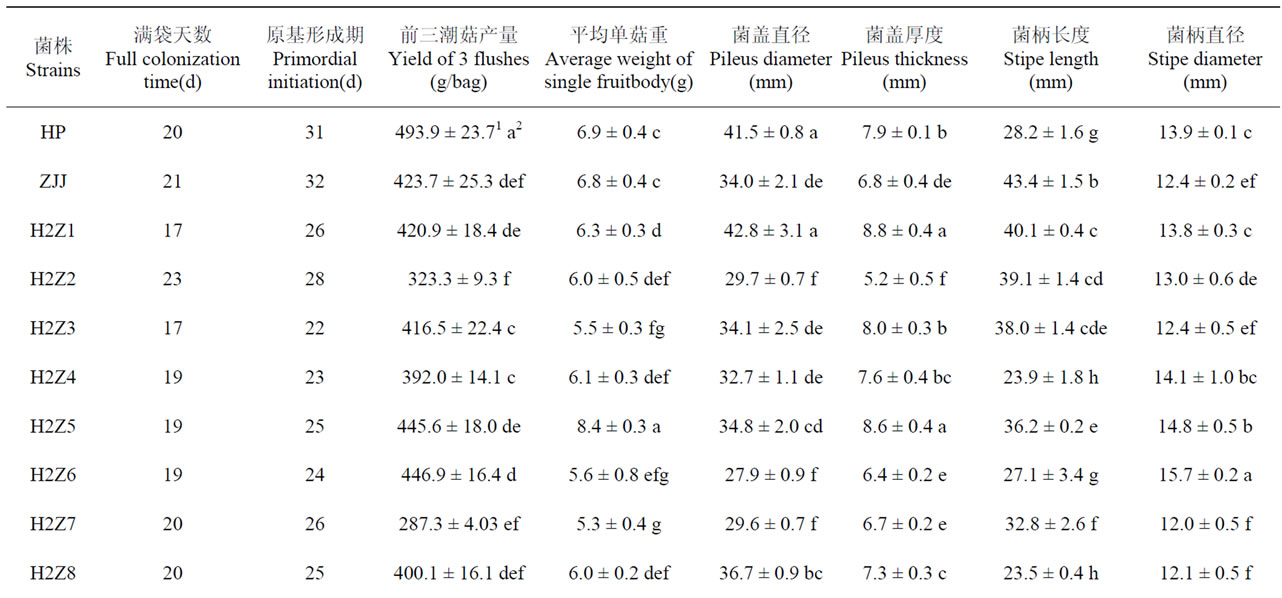
Table 1. The statistical results of quantitative traits of hybrids and its’ parents in Pleurotus ostreatus
表1. 平菇杂交子和亲本数量性状统计结果

Table 2. The statistical results of agnomic chracteristics index of hybrids in Pleurotus ostreatus
表2. 平菇杂交子农艺性状指标的统计结果
和菌柄直径(9.4%)。杂交子满袋天数、原基形成期、单袋产量、单菇重量、菌盖直径、菌盖厚度、菌柄长度和菌柄直径的变异幅度分别为17 d~35 d、22 d~45 d、96.5 g~469.1 g、5.34 g~9.38 g、27.9 mm~42.8 mm、5.2 mm~8.8 mm、23.5 mm~47.5 mm和12.0 mm~15.7 mm。由杂交子各个性状变异程度的差异可见,杂交F1代性状分离和遗传重组极具多样性,不同的基因重组以及对环境的适应性表现为子代表型特征的广泛变异,变异系数大的农艺性状在后代的遗传变化较大,可根据不同的育种目标,对杂交子代进行选择。
4. 讨论
本研究中对野生菌株ZJJ进行ITS序列测定,结果发现ZJJ与黄晨阳等所构建的系统发育树中肺形侧耳(P. pulmonarius)聚为一类(黄晨阳等,2010),但与糙皮侧耳(Pleurotus ostreatus)亲缘关系也较近。真菌物种的鉴定仅靠ITS序列分析是不够的,还需要结合野生子实体形态、解剖特征、菌丝结构、孢子大小等微观特征等进行。
近年来关于侧耳属内种的分类仍然众说纷纭,关于侧耳属真菌系统分类的最合理有效的方法尚无定论。Bao等通过性亲和试验鉴定糙皮侧耳和肺形侧耳二者杂交不亲和,为不同的生物学种(Bao D.P. et al., 2004),但菌株之间的杂交不育或可育是在大量测试的种群内才能发生的,真正的杂交不育应是全世界范围内的(王呈玉等,2006)。28S rDNA序列分析表明糙皮侧耳与肺形侧耳具有很近的亲缘关系(Guzmán, 2000;Vilgalys et al., 1994),Petersen等发现一个来自新西兰的肺形侧耳菌株与糙皮侧耳弱亲和(Petersen R H et al., 1996),Bao等通过性亲和试验证明肺形侧耳与佛罗里达侧耳(Pleurotus florida)可亲和,Guzman发现凤尾菇(Pleurotus sajor-caju)与糙皮侧耳交配亲和,并产生了典型的糙皮侧耳子实体(Guzman et al., 1994)。而Gonzalez等对线粒体小亚基rDNA的可变区V4,V6, V9区域研究表明,佛罗里达侧耳和糙皮侧耳,肺形侧耳和凤尾菇应该是同一个物种的不同名称(Gonzalez et al.,2000)。显然,糙皮侧耳可能是一个包含肺形侧耳与佛罗里达侧耳在内的复合种。基于杂交育种的目的,本研究只做了10 × 10配对杂交,还不能完全反映两个菌株之间遗传距离和性亲和规律,也不能确定两个供试菌株是否属于不同的物种。
杂交子与亲本的拮抗反应类型与子实体形态分类具有一定的关联性。H5Z1、H5Z7和H5Z10三个菌株均只能形成无菌盖的子实体,它们与亲本HP均形成菌丝集结隆起并有明显的色素沉着的拮抗线,与亲本ZJJ均形成无明显的菌丝集结隆起又无色素产生的拮抗线;H5Z2未形成原基,它与亲本的拮抗反应表现与其它13个杂交子均不相同。
菌丝体在PDA、三级菌种培养基、栽培料培养基中的生长速度并未呈现一致性。有研究表明菌丝生长速度是由基因、培养基组成共同控制的,降解木质素和纤维素是由多种酶谢协同完成的,有的酶是组成型表达,有的是诱导表达,而PDA培养基、栽培种培养基以及栽培料的营养成分是有很大不同的(Silva EM et al., 2005b; Lakhtar H et al., 2007; Elisashvili V et al., 2008)。
参考文献 (References)
[1] D. P. Bao, S. Kinugasa and Y. Kitamoto. The biological species of oyster mushrooms (Pleurotus spp.) from Asia based on mating compatibility tests. Journal of Wood Science, 2004, 50(2): 162-168.
[2] V. Elisashvili, M. Penninckx, E. Kachlishvili, N. Tsiklauri, E. Metreveli, T. Kharziani, G. Kvesitadze. Lentinus edodes and Pleurotus species lignocellulolytic enzymes activity in submerged and solid state fermentation of lignocellulosic wastes of different composition. Bioresource Technology, 2008, 99(3): 457-462.
[3] G. Guzman, L. Montoya, G. Mata and D. Salmones. Studies in the genus Pleurotus Ⅲ. The varieties of P. ostreatus complex based in interbreeding strains and in the study of basidiomata obtained in culture. Mycotaxon, 1994, 50(1): 365-378.
[4] P. Gonzalez, J. Labarère. Phylogenetic relationships of pleurotus species according to the sequence and secondary structure of the mitochondrial small-subunit rDNA V4, V6 and V9 domains. Microbiology, 2000, 146(1): 209-221.
[5] G. Guzmán. Genus Pleurotus (Jacq.: Fr.) P . Kumm. (Agaricomycetideae): Diversity, taxonomy problems, and cultural and traditional medicinal uses. International Journal of Medicinal Mushrooms, 2000, 2(1): 95-123.
[6] Y. Hadar, Z. Kerem, B. Gorodecki. Biodegradation of lignocellulosic agricultural wastes by Pleurotus ostreatus. Journal of Biotechnology, 1993, 30(1): 133-139.
[7] A. Helbling, F. Gayer, W. J. Pichler, et al. Mushroom (Basidiomycete) allergy: Diagnosis established by skin test and nasal challenge. Journal of Allergy and Clinical Immunology, 1998, 102(5): 853-858.
[8] 黄晨阳, 陈强, 高山等. 侧耳属主要种类ITS 序列分析[J]. 菌物学报, 2010, 29(3): 365-372.
[9] K. Jatinder, S. S. Harpreet and K. Shammi. Breeding of Pleurotu florida (oyster mushroom) for phenotypic pigmentation and high yield potential. Journal of the Science of Food and Agriculture, 2008, 88(15): 2676-2681.
[10] 李红梅, 尚晓冬, 谭琦等. 刺芹侧耳自交S1代若干性状的遗传分化[J]. 菌物学报, 2009, 28(4): 541-547.
[11] H. Lakhtar, M. Ismaili-Alaoui, A. Philippoussis, H. Macarie and S. Roussos. Preliminary studies on Lentinula edodes strains able to degrade polyphenols in olive wastewater. XVI National Bioprocess Symposium—SINAFERM, Curitiba, 2007.
[12] R. H. Petersen, G. S. Ridley. A New Zealand Pleurotus with multiple-species sexual compatibility. Mycologia, 1996, 88(2): 198-207.
[13] 戚元成, 张小强, 高玉千等.中国栽培糙皮侧耳品种体细胞不亲和性实验与RAPD分析结果的比较[J]. 菌物学报, 2010, 29(3): 379-388.
[14] E. M. Silva, A. Machuca, A. M. F. Milagres. Evaluating the growth and enzyme production from Lentinula edodes strains. Process Biochemistry, 2005, 40(1): 161-164.
[15] R. G. Thorn, J. M. Moncalvo, C. A. Reddy, et al. Phylogenetic analyses and the distribution of nematophagy support a monophyletic Pleurotaceae within the polyphyletic pleurotoid-lentinoid fungi. Mycologia, 2000, 92(2): 241-252.
[16] R. Vilgalys, B. L. Sun. Ancient and recent patterns of geographic speciation in the oyster mushroom Pleurotus revealed by phylogenetic analysis of ribosomal DNA sequences. Proceedings of the National Academy of Sciences, 1994, 91(10): 4599-4603.
[17] 王呈玉, 图力古尔, 李玉. 侧耳属真菌系统分类研究概况[J].吉林农业大学学报, 2006, 28(1): 158-163.
NOTES
*基金项目:公益性行业(农业)科技专项“食用菌菌种质量评价与菌种信息系统研究与建立”和湖北省自然科学基金重点项目。