Bioprocess
Vol.3 No.1(2013), Article ID:9623,6 pages DOI:10.12677/BP.2013.31002
Isolation and Characterization of a Novel Medium-Long Alkane-Degrading Strain*
College of Life Sciences, China West Normal University, Nanchong
Email: #tangyun_502@yahoo.com.cn
Received: Dec. 3rd, 2012; revised: Dec. 8th, 2012; accepted: Feb. 1st, 2013
ABSTRACT:
A strain of medium-long chain n-alkanes degrading strain TY12 was screened from petroleum-contaminated soils of Nanchong oil refinery. We adopted several phenotypic and genotypical methods, such as morphological, physical and biochemical characteristics, antibiotic susceptivities, 16S rRNA gene sequences based phylogenetic analysis, to outline the basic biological characteristics and determinate the phylogenetic position. The Gram-negative isolate TY12 was a member of the genus Acinetobacter, short rods, no capsules, no endospores, catalase-positive, oxidase-negative, 0.9 - 1.6 μm in diameter and 1.5 - 2.5 μm in length; TY12 was susceptible to 14 kinds of tested antibiotics but resistant to Tetracycline, Amoxicillin, Ampicillin; The similarity between its 16S rRNA gene and that of its most closely related type strain in GenBank Acinetobacter beijerinckii CCM7266 (AJ626712), Acinetobacter beijerinckii LΜH6214 (AJ303013) were 100%. The optimal temperature and pH for the strain utilizing industrial ethanol were 30˚C and 7.0, and the optimal concentration of ethanol and concentration of yeast extract were 0.7% and 0.025 g/L, respectively. The n-dodecane degradation was 92% after the strain was growing on in hydrocarbon degradation medium with 1% (W/V) of n-dodecane at 30˚C and 180 r/min for 36 h. The strain could degrade a large range of n-alkanes with chain length C12 - C32. It has potential in bioremediation of oil contaminated environment.
Keywords: Petroleum Pollution; Hydrocarbon-Degrading Bacteria; Isolation and Identification; Acinetobacter
一株高效降解中长链烷烃菌株的分离鉴定及其降解特性研究*
唐 赟#, 谭 洪
西华师范大学生命科学学院,南充
Email: #tangyun_502@yahoo.com.cn
摘 要:
从南充市炼油厂被石油污染土壤中分离到一株中长链烷烃降解菌TY12。采用形态学观察、生理生化指标测定、抗生素抗性实验、16S rRNA基因序列同源性分析等多种方法对该菌株进行鉴定。菌株为Acinetobacter beijerinckii,革兰氏阴性,短杆状,无荚膜,无芽孢,接触酶阳性,氧化酶阴性,菌体大小为0.9~1.6 μm × 1.5~2.5 μm;对新霉素、氯霉素、卡拉霉素等14种抗生素敏感,而对四环素、阿莫西林、氨苄西林耐药;在GenBank中与其16S rRNA基因序列相似度最高的模式株分别为Acinetobacter beijerinckii CCM7266 (AJ626712)、Acinetobacter beijerinckii LMH6214 (AJ303013),相似性为100%。研究表明,以工业乙醇为唯一碳源的无机盐培养基培养种子最适宜;其最适温度和pH分别为30℃和7.0,无水乙醇最适加入量为0.7%,最适酵母粉浓度为0.025 g/L。在初始正十二烷浓度为1%(W/V)培养基中接入1%种子液,于30℃、180 r/min震荡培养36 h,正十二烷降解率可达92%;菌株对正构烷烃的降解范围较宽,能以C12~C32的正构烷烃为唯一碳源和能源生长。该菌是一株能够降解中长链烷烃的不动杆菌新种,具有应用到石油污染修复的潜力。
收稿日期:2012年12月3日;修回日期:2012年12月8日;录用日期:2013年2月1日
关键词:石油污染;烷烃降解;分离鉴定;不动杆菌
1. 引言
石油是人类最重要的能源之一,随着科技进步和日益加快的工业化步伐,世界各国都加快了对石油资源的开发和利用。在石油开采、加工和运输过程中,石油及其产品进入环境引起环境的污染和破坏。因此,对石油污染的治理成为近年来世界各国环境保护的焦点,人们不断研究发展了多种修复方法如:物理修复法、化学修复法和生物修复法等。其中,生物修复技术由于具有成本费用低、效果好、对环境污染小、无二次污染等优点,受到越来越多国家的欢迎[1]。
石油中含量最高的成分是各种链长的烷烃,占总质量的20%~50%,因此在众多的石油污染中,烷烃是最常见的污染源,其生物降解一直以来都是研究的热点。目前已分离到多种烷烃和芳香烃降解菌[2-4]。其中,不动杆菌属是主要的种属之一,该属目前已报道有五个种具有降解烷烃的能力,第一个种是由Kleber HP等[5]人分离到的Acinetobacter calcoaceticus EB104;而最具代表性的是由Sakai Y等[6]人分离到的Acinetobacter sp. M-1,它能利用C13~C44为唯一的碳源和能源,在石油污染的修复过程中起到非常重要的作用。
我们研究小组从南充市炼油厂被石油污染土壤中分离筛选到一株中长链烷烃降解菌Acinetobacter beijerinckii TY12。采用形态学观察、生理生化指标测定、抗生素抗性实验、16S rRNA基因序列同源性分析等多种方法对其进行了分类学鉴定,并对其正构烷烃利用情况进行了初步分析,表明菌株Acinetobacter beijerinckii TY12一株能够降解中长链烷烃的不动杆菌新种。
2. 实验材料与方法
2.1. 样品来源
样品取自南充市炼油厂被石油污染的土壤,南充炼油厂位于南充市主城区北半部的中心位置,处于城区上风向。占地面积50.88万平方米,年加工原油能力100万吨。采集的样品立即放入无菌塑料袋中送至实验室富集培养。
2.2. 材料与试剂
正己烷,正辛烷,正十烷,正十二烷,正十四烷,正十六烷,正十八烷,正二十二烷,正二十四烷,正二十八烷,正三十二烷,所有烷烃的纯度>98%,均购自天津化学试剂研究所。蛋白胨,酵母粉,琼脂粉,NaNH4HPO4·4H2O,K2HPO4,MgSO4·7H2O,KH2PO4,NaCl等均为国产分析纯,均购自Sigma公司。TIANGEN细菌基因组DNA提取试剂盒,TIANGEN纯化回收试剂盒,TIANGEN普通质粒小提试剂盒均购自成都博瑞克生物技术有限公司。生化鉴定管购自青岛海博生物技术有限公司;药敏试剂盒购自杭州天和微生物试剂有限公司。
2.3. 培养基
2.3.1. E2无机盐培养基[7]
NaNH4HPO4.4H2O 3.5 g,K2HPO4 7.5 g,KH2PO4 3.6 g加水至1000 mL,调节pH值7.0,于121℃灭菌20 min。固体培养基在此基础上加15 g琼脂粉,于121℃灭菌20 min后加入1 mol/L MgSO4 1.0 mL和微量元素溶液II 1.0 mL。
微量元素溶液II(250 mL):FeSO4.7H2O 0.695 g,MnCl2.4H2O 0.5 g, CoSO4.7H2O 0.7 g,CaCl2.2H2O 0.278 g,CuCl2.2H2O 0.043 g,ZnSO4.7H2O 0.073 g,溶于125 mL 1mol/L的HCl中,再用250 mL的容量瓶定容,过滤除菌。
2.3.2. 乙醇过渡培养基[8]
无机盐培养基,加0.7%的工业乙醇。固体培养基在此基础上加1.5%的琼脂粉。
2.3.3. 正构烷烃培养基
无机盐培养基,加1%(W/V)的正构烷烃。固体培养基在此基础上加1.5%的琼脂粉。
2.3.4. LB培养基[9]
2.4. 分析测定方法
2.4.1. 细菌生长量的测定
以未接种的培养基为对照,用722型分光光度计在600 nm波长下测定菌悬液的OD600nm。
2.4.2. 烷烃的定量测定
在50 mL烷烃培养液中加入5 mL色谱纯正己烷及等量正十四烷(内标),振荡2 min后迅速倒入离心管中,在4℃、10,000 r/min条件下离心5 min,抽取上清液,再分别加入5 mL正己烷萃取两次,合并三次萃取液,并作适当稀释后按以下方法进行气相色谱分析。
用HP4890气相色谱仪(GC-FID)进行分析,色谱柱为美国KB-1(0.32 mm × 30 m × 0.25 μm)。运行条件:柱温先在150℃维持1 min,然后以15℃/min速率升至250℃;检测器温度为280℃;注射室温度250℃;进样量1 mL。通过测定与正十四烷的峰面积比值,定量分析各种烷烃的质量浓度。
2.5. 烷烃高效降解菌株的富集和分离
将采集的土壤样品用玻璃珠打碎后静置,取上清液加入到十二烷培养基中,于30℃、180r/min振荡恒温培养箱中培养7 d,将培养后的菌液经梯度稀释后涂布在倒置的LB固体烷烃平板上,于30℃生化培养箱中培养2 d;挑取平板上长出的单菌落进行重复的分离纯化,直到得到单菌株。挑取单菌落接种到烷烃培养基中,测试各菌种在30℃、180 r/min培养72 h时对正十二烷的利用情况,筛选出对烷烃降解高效菌株。
2.6. 菌株的鉴定
2.6.1. 菌株形态鉴定
采用扫描电子显微镜观察菌体形态,电镜照片样品制备参照文献[10]所述方法。
2.6.2. 菌株生理实验
最适温度、最适pH、最适NaCl浓度等生长实验采用LB培养基。药敏实验采用K-B法,所用的药敏纸片由杭州微生物试剂有限公司提供。
2.6.3. 菌株生化实验
革兰氏染色镜检形态、氧化酶、接触酶实验及微量生化反应管生化鉴定等,参照相关细菌鉴定手册进行[11,12]。
2.6.4. 菌株16S rRNA序列测定
1) 菌体总DNA的提取和纯化取过夜培养菌液,离心收集菌体,用细菌基因组提取试剂盒提取并纯化基因组DNA。
2) 16S rRNA基因序列的PCR扩增及克隆[13]以TY12总DNA为模板,采用通用fD1.rD1引物(fD1: 5-AGAGTTTGATCCTGGCTCAG-3,rD1: 5-AAGCAGGTGATCCAGCC-3)进行16S rRNA的PCR扩增[14]。100 μL的PCR反应体系组成如下:100倍稀释DNA模板4 μL,10 μmol/L引物fD1 10 μL,10 μmol/L引物rD1 10 μL,2 × MasterMix 50 μL,加dd H2O至总体积为100 μL。PCR反应条件为:94℃预变性5 min;94℃变性1 min,55℃退火45 s,72℃延伸1 min,总计25个循环;最后延伸10 min。扩增得到的DNA片段并与pMD19-T载体连接,转化大肠杆菌DH5α。对正确的重组子[15],送北京泰和生物技术有限公司进行DNA测序。将测序结果与GenBank上的相关16S rRNA序列在Clustal X2程序包中进行多重序列匹配排列分析,用MEGA(5.0)软件建系统进化树[16]。
2.7. 烷烃降解菌降解特性的研究
2.7.1. 种子液的制备
在滩涂石油污染的生物修复中,种子液将制成一定剂型使用,这就要求其成分不能有烃类物质,避免加重污染。其碳源选择只有糖类和醇类,考虑到菌株能够很好的利用工业乙醇,且制备的种子液不易变质,便于保存,故选用工业乙醇为碳源。
实验不同温度、pH值、碳源浓度及生长因子浓度对菌株生长的影响,并在最佳的培养条件下测定菌株的生长曲线。
2.7.2. 菌株TY12在正十二烷中生长情况
取过渡培养基中生长12 h的菌液培养物0.5 mL接入50 mL的以十二烷为唯一碳源和能源的十二烷液体培养基中(十二烷含量为1%W/V),30℃,180 r/min振荡培养。观察观察菌体生长情况并测定正烷烃的总降解率。
2.7.3. 纯烃底物降解实验
将TY12菌液按1%接入装有50 mL液体培养基的250 mL三角瓶中培养36 h,摇瓶转速180 r/min,温度为30℃,每种底物做3个重复。以不加任何碳源的接菌培养液做空白对照,测定培养液OD600nm,观察菌体生长情况。
3. 结果和讨论
3.1. 烷烃降解菌的分离与筛选
从南充炼油厂经常被含油废水浸泡的土壤上采集的土壤,经过富集、分离纯化,共获得13株单菌,分别取代号为TY6-TY18。将分离的单菌接种到十二烷培养基中,根据菌株在30℃的生长曲线和烷烃降解曲线,最后选择生长速度快,对正十二烷降解率最大的TY12用于进一步的实验。
3.2. 菌株鉴定
3.2.1. 菌株的培养特征
菌株TY12在LB固体培养基上培养12 h,生长良好,菌落直径大约1.0~2.0 mm,呈淡黄色,不透明,圆形,边缘平整,中间略隆起,表面略湿,有微弱光泽,不粘稠,无臭味。在半固体培养基中,在琼脂表面生长旺盛,穿刺线不明显。在液体LB培养基中呈均匀浑浊,静止时液体呈及微薄的菌膜,轻摇则散。菌体革兰氏染色阴性,抗酸性染色阴性,短杆状,两端略钝,直或略弯,成双排列、也少量不同程度的链状,无荚膜,无芽孢,大小为0.9~1.6 μm × 1.5~2.5 μm,如图1。
3.2.2. 生理特征
菌株生长温度为25℃~37℃,最适温度为30℃;NaCl浓度小于4%能生长,最适浓度为0.5%~1.5%;由pH实验得出,菌株TY12生长pH为5.0~10.0,最适pH为6.0~8.0。
抗生素实验结果表明,菌株TY12对链霉素、卡拉霉素、大观霉素、氯霉素、氧氟沙星、庆大霉素、恩诺沙星、新霉素、复方新诺明、环丙沙星高度敏感;

Figure 1. Microphotograph of the strain TY12 (1000×)
图1. 菌株TY12的显微照片(1000×)
对青霉素G、克林霉素、阿米卡星、乙酰螺旋霉素等中度敏感;对阿莫西林、氨苄西林、四环素不敏感。
3.2.3. 生化特征
接触性酶阳性,氧化酶阴性,不能水解明胶、淀粉和酪蛋白;不能利用葡萄糖、鼠李糖、蔗糖、蜜二糖、苦杏仁苷、阿拉伯糖、水杨苷、木糖、海藻糖、纤维二糖、苯甲酸、己二酸、酒石酸盐、壬二酸盐、腐胺、精氨酸、赖氨酸、鸟氨酸、苯丙氨酸作为碳源,微弱利用乳糖、肌醇、山梨醇、麦芽糖、乳糖、棉籽糖、七叶苷。能利用乙醇、苹果酸盐、醋酸盐、硝酸盐、柠檬酸盐、丙二酸盐作为碳源。由表1生理生化特征对比可以看出,菌株TY12具有与A. beijerinckii特别相近生理生化特征,但TY12发酵DL-乳糖产酸微弱;而与A. venetianus和A. junii的生化特性相差较大,其中后三者的生理生化特征参照文献[17]所描述。因此,可以初步推断TY12属于A. beijerinckii。
3.2.4. 16S rRNA基因序列的系统进化分析
通过对菌株TY12的16S rRNA进行PCR扩增、测序,获得1541 bp大小的基因序列,此序列在GenBank上的登录号为:JF742664。将16S rRNA序列与GenBank + EMBL + DDBJ + PDB数据库中的近缘种进行BLAST(basic local alignment search tool)比对分析。图2是采用邻位相连法(Neighbor-joining method)

Table 1. Comparison of selected phenotypic characteristics of TY12 and reference strains
表1. TY12菌株与相近菌株的主要特征比较
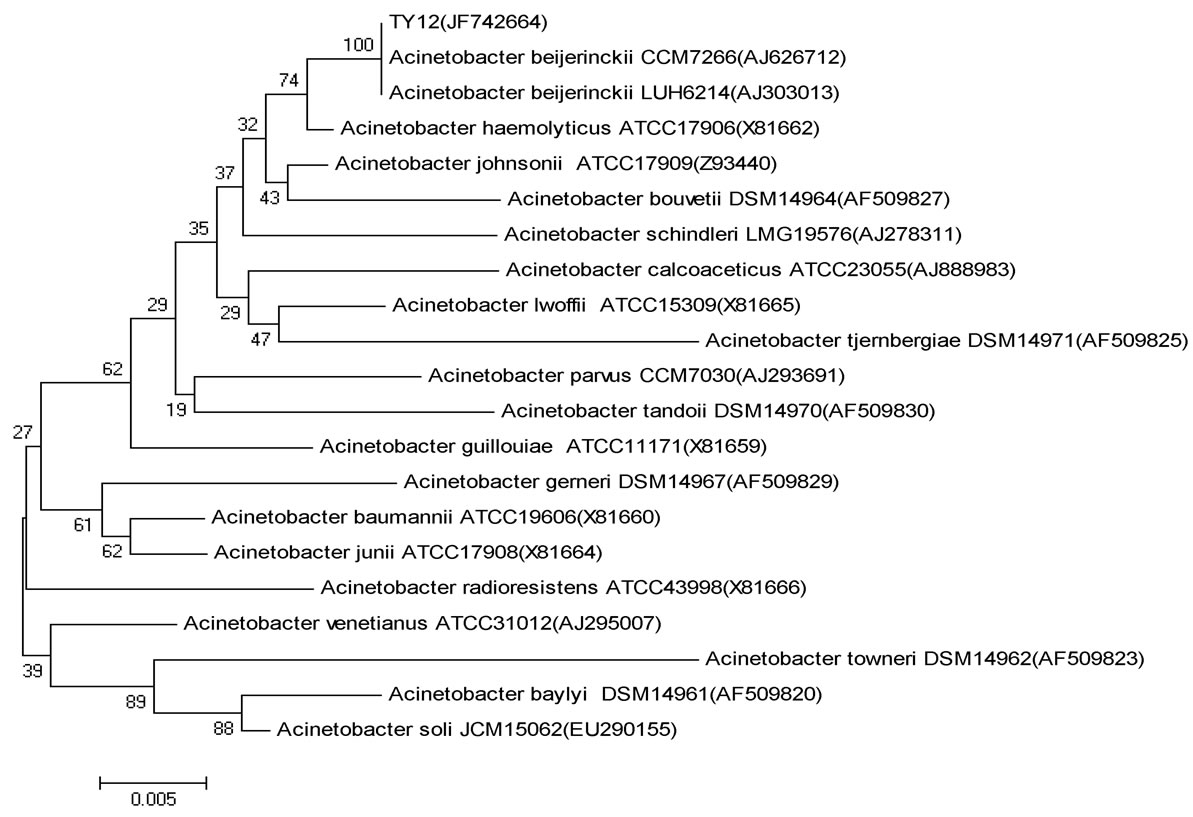
Figure 2. Phylogenetic tree demonstrating the relationship of the 16 S rRNA of the strain TY12 closest related Acinetobacter. Scale bar indicates evolutionary distance
图2. 基于菌株TY12的16 S rRNA序列同源性菌株系统发育树
构建的菌株TY12与Acinetobacter属的20株模式种的16S rRNA系统进化树。由图2所示,菌株TY12和模式菌株Acinetobacter beijerinckii CCM7266 (AJ626712)、Acinetobacter beijerinckii LΜH6214 (AJ303013)的进化距离最小,相似性为100%。结合传统的生理生化特性分析,确定菌株TY12为A. beijerinckii。
3.3. 烷烃降解菌降解特性的研究
3.3.1. 种子液的制备
对种子液培养的温度、pH、培养基碳源浓度、生长因子浓度进行了选择性实验。如图3~6所示,TY12在30℃时生长良好;在pH值为7.0时生物量达到最高;当加入工业乙醇浓度0.7%、生长因子浓度0.025 g/L时,菌体浓度最高。
在最佳的条件下,震荡培养20 h,每隔1 h取样,以培养时间为横坐标,OD600nm为纵坐标,测菌株的生长曲线。如图7所示,菌株培养2 h开始进入对数生长期,到14 h左右进入生长稳定期,确定菌株处于对数生长后期的培养时间。选择培养12 h菌液接种烃降解培养基,此时种子液菌体浓度可以达到108 cells/ ml。
3.3.2. 菌株TY12在正十二烷中生长情况
取新鲜的种子液按1%接种量接入50 mL正十二烷浓度为1%(W/V)无机盐培养基中,于30℃、180
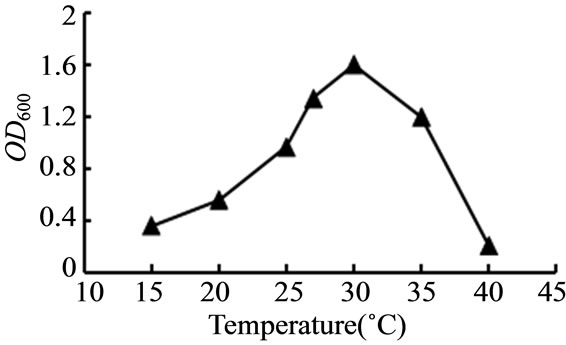
Figure 3. Effect on the growth of the strain TY12 by temperature
图3. 温度对菌株TY12生长的影响
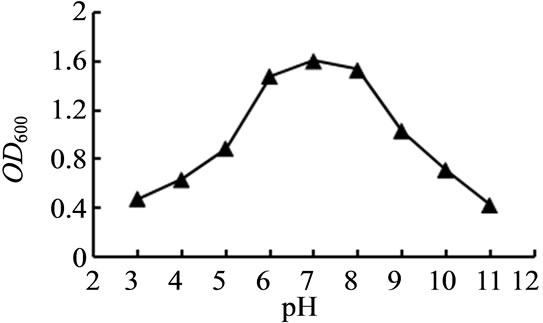
Figure 4. Effect on the growth of the strain TY12 by pH
图4. pH对菌株TY12生长的影响
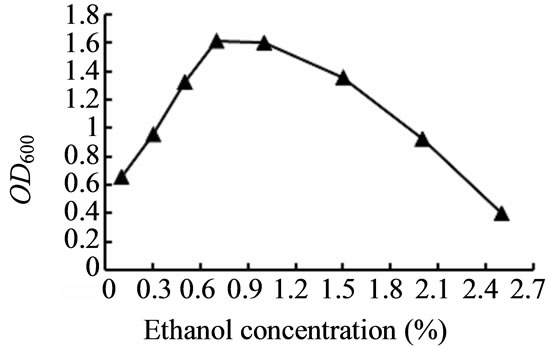
Figure 5. Effect on the growth of the strain TY12 by Industrial ethanol
图5. 工业乙醇对菌株TY12生长的影响

Figure 6. Effect on the growth of the strain TY12 by yeast extract
图6. 酵母粉对菌株TY12生长的影响
Figure 7. Growth curve of the stain TY12
图7. 菌株TY12种子液生长曲线
r/min振荡恒温培养箱中培养,每隔4 h取样,测定培养液OD600 nm及残留正十二烷量,以确定菌株TY12生长量与正十二烷降解量之间的关系,结果见图8。由图8所示,TY12在正十二烷培养基中,延迟期约为3 h,随后生长迅速,到32 h左右生长量最大。细菌生长量和正十二烷的降解率是相平行的。培养到14 h时,正十二烷的降解率达到约45%,OD600nm为0.938;其后,菌株出现了一次比较明显的生长现象,OD600nm在26 h达到1.753,而降解率升高到74%,可能是中链烷烃分解成的短链烷烃为菌株的进一步生长提供了碳源;培养到36 h时,正十二烷的降解率达到约92%,OD600nm为1.909。
3.3.3. 烷烃降解范围
按方法[1.7.3],以不同的烷烃作为唯一碳源,接入1%的TY12种子液,培养36 h观察,结果如图9所示。菌株TY12在C6中生长十分的缓慢,C8、C10中几乎不生长,推测可能是因为该菌对这些短链烷烃的毒性作用比较敏感或缺乏利用机制。该菌在C12~C28中生长良好,在C32中生长稍微缓慢些,这可能是长

Figure 8. Growth and degrading curve of the strain TY12 with n-dodecane as sole carbon resource
图8. 菌株TY12以正十二烷为唯一碳源生长和降解曲线
Figure 9. Growth of the strain TY12 with different length of n-alkanes as sole carbon resource
图9. 菌株TY12以不同长度碳链烷烃为唯一碳源的生长情况
链烷烃在水中的溶解度非常小,从而菌株对其可利用性降低。不动杆菌是目前发现的革兰氏阴性烷烃降解菌中降解范围较广的降解菌,之所以在石油污染的治理中能起着重要的作用,与它们宽域的降解谱是密切相关的。
4. 结论
由于细菌的16S rRNA具有很高的保守性,因而近十年来细菌分类学家们广泛采用16S rRNA序列分析法进行发育分类学研究。通过将某一未知细菌的16S rRNA编码基因(16S rRNA)的序列与基因库的16S rRNA序列进行同源性比较,可以初步确定该细菌的种属地位,有些细菌甚至可以鉴定到种的水平。通过16S rRNA序列分析,结合形态学观察、生理生化指标测定、抗生素抗性实验等多种方法进行鉴定,将TY12归属为Acinetobacter beijerinckii。不动杆菌属细菌是条件致病菌,可引起呼吸道感染、败血症、脑膜炎、心内膜炎、伤口及皮肤感染、泌尿生殖道感染等。抗生素实验也表明该菌对多种抗生素不敏感,这将严重影响其实际应用。随着分子生物学的发展,可以使用基因敲除方法对菌株进行基因改造,获得去除毒力因子的目的菌株。
目前人们已经研究了Alcanivorax sp. A-11-3A[18]、Gordonia sp. S14-10[19]、Geobacillus thermodenitrificans NG80[20]等菌株的烷烃降解酶系和相关基因。已报道的AlkBs和CYP153s都是负责中等链长烷烃(C5~C12)的降解,对于长链烷烃羟化酶AlmA和LadA的报道却不多见。Maier T等人[21]从Acinetobacter calcoaceticus EB104克隆到了参与烷烃羟化的P450基因。Mimmi Throne-Holst等人[22,23]从Acinetobacter venetianus 6A2中克隆到了参与烷烃羟化的alkMa、alkMb、almA基因。菌株Acinetobacter beijerinckii TY12具有降解中长链烷烃能力、也是未曾报道的新种,烷烃降解酶基因的研究可以作为继续研究的方向。
参考文献 (References)
[1] 陆健, 黄潇, 伍贤明等. 高效烷烃降解菌xcz的分离鉴定及降解特性[J]. 土壤, 2008, 40(3): 460-464.
[2] V. Mishra, R. Lal and Srinivasan. Enzymes and operons mediating xenobiotic degradation in bacteria. Critical Reviews in Microbiology, 2001, 27(2): 133-166.
[3] N. Das, P. Chandran. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnology Research International, 2011, 2011: Article ID 941810.
[4] S. Harayama, Y. Kasai and A. Hara. Microbial communities in oil contaminated seawater. Environmental Biotechnology, 2004, 15(3): 205-214.
[5] H. P. Kleber, R. Claus and O. Asperger. Enzymologie der n-alkanoxidation bei acinetobacter. Acta Biotechnologica, 1983, 3(3): 251-260.
[6] Y. Sakai, J. H. Maeng, Y. Tani and N. Kato. Use of long-chain n-alkanes (C13-C44) by an isolate, Acinetobacter sp. M-1. Bioscience, Biotechnology, and Biochemistry, 1994, 58(11): 2128- 2130.
[7] R. G. Lageveen, G. W. Huisman, H. Preusting, et al. Formation of polyesters by Pseudomonas oleovarans: Effect of substrates on formation and Composition of Poly-(R)-3-Hydroyalkanoates and Poly-(R)-3-Hydroxyalkenoates. Applied and Environmental Microbiology, 1988, 54(12): 2924-2932.
[8] 黄磊, 李丹, 孙丹, 谢玉娟等. 一株低温石油烃降解菌的分类鉴定及降解特性研究[J]. 环境科学, 2007, 28(9): 2101-2102.
[9] F. C. Neidhardt, P. L. Bloch and D. F. Smith. Culture medium for enterobacteria. J. Bacterial, 1974, 119(3): 736-747.
[10] 仇天雷, 承磊, 邓宇等. 一株近海沉积物中产甲烷菌的分离及鉴定[J]. 中国沼气, 2007, 25(2): 3-6.
[11] 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001: 353-398.
[12] R. E. 布坎南, N. E. 吉本斯, 著, 中国科学院微生物研究所《伯杰细菌鉴定手册》翻译组, 译. 伯杰鉴定手册(第八版) [M]. 北京: 科学出版社, 1995: 591-611.
[13] J. 萨姆布鲁克, D. W. 拉赛尔, 著, 黄培堂等, 译. 分子克隆实验指南(第三版). 北京: 科学出版社, 2002.
[14] W. G. Weisburg, S. M. Barna, D. A. Pelletier, et al. 16S ribosomal DNA amplification for phylogenetic study. Journal of Bacteriology, 1991, 173(2): 697-703.
[15] 陈书霞, 王晓武, 房玉林. 单菌落PCR法直接快速鉴定重组克隆[J]. 微生物学通报, 2006, 33(3): 52-56.
[16] N. Saitou, M. Nei. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 1987, 4(4): 406-425.
[17] A. Nemec, M. Musilek, M. Maixnerova, et al. Acinetobacter beijerinckii sp. nov. and Acinetobacter gyllenbergii sp. nov., haemolytic organisms isolated from humans. Journal of Systematic and Evolutionary Microbiology, 2009, 59(1): 118-124.
[18] 吴业辉, 邵宗泽. 海洋烷烃降解菌Alcanivorax sp. A-11-3的分离鉴定及其降解酶基因研究[J]. 台湾海峡, 2008, 27(4): 427-434.
[19] 王丽萍, 刘昱慧, 邵宗泽. 一株来自大西洋表层海水的烷烃降解菌Gordonia sp. S14-10的分离鉴定及其降解相关特性的分析[J]. 微生物学报, 2009, 49(12): 1634-1642.
[20] L. Li, X. Liu, W. Yang, et al. Crystal structure of long-chain alkane monooxygenase (LadA) in complex with coenzyme FMN: Unveiling the long-chain alkane hydroxylase. Journal of Molecular Biology, 2008, 376(2): 453-465.
[21] T. Maier, H. H. Foerster, O. Asperger, et al. Molecular characterization of the 56-kDa CYP153 from Acinetobacter sp. EB104. Biochemical and Biophysical Research Communications, 2001, 286(3): 652-658.
[22] M. Throne-Holst, A. Wentzel, T. E. Ellingsen, et al. Identification of novel genes involved in long-chain n-alkane degradation by Acinetobacter sp. strain DSM 17874. Applied and Environmental Microbiology, 2007, 73(10): 3327-3332.
[23] M. Throne-Holst, S. Markussen, A. Winnberg, et al. Utilization of n-alkanes by a newly isolated strain of Acinetobacter venetianus: The role of two AlkB-type alkane hydroxylases. Applied Microbiology and Biotechnology, 2006, 72(2): 353-360.
NOTES
*基金项目:四川省教育厅自然科学重点项目(09ZA123)和南充市科技局重点项目([2009]1067)资助。
#通讯作者。