Bioprocess
Vol.4 No.04(2014), Article
ID:14492,6
pages
DOI:10.12677/BP.2014.44009
Comparative Study of Digestive Tract Morphology of Apodemus chevrieri in Chenggong and Jianchuan of Yunnan Province
Key Laboratory of Adaptive Evolution and Ecological Conservation on Plants and Animals in Southwest Mountain Ecosystem of Yunnan Higher Education Institutes, School of Life Sciences, Yunnan Normal University, Kunming
Email: *zwl_8307@163.com
Copyright © 2014 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received: Aug. 21st, 2014; revised: Sep. 4th, 2014; accepted: Oct. 10th, 2014
ABSTRACT
In order to investigate whether there are differences on digestive tract morphology in A. chevrieri from different areas, we measured weight and lengths of total digestive tract morphology including stomach, small intestine, large intestine and cecum of A. chevrieri from Jianchuan and Chenggong. The results showed that the length and weight without contents of the small intestine and cecum in A. chevrieri at Jianchuan were significantly greater than that of Chenggong; body mass, the stomach, the weight with content of stomach and cecal and lengths of cecum at Jianchuan were significantly less than that of Chenggong. The results indicated that A. chevrieri at Jianchuan might adjust the body mass and weight of digestive tract to maintain the normal physiological function in stressed condition, such as at cold environment and lack of food. This suggests that the adjustments in capacity and size of the gastrointestinal tract play an important role in their digestive strategies of A. chevrieri in natural environment.
Keywords:Apodemus chevrieri, Different Regions, Digestive Tract Morphology

云南呈贡和剑川地区
高山姬鼠消化道形态的比较
郑 佳,朱万龙*
云南省高校西南山地生态系统动植物生态适应进化及保护重点实验室,云南师范大学生命科学学院,昆明
Email: *zwl_8307@163.com
收稿日期:2014年8月21日;修回日期:2014年9月4日;录用日期:2014年10月10日

摘 要
为探讨不同地区的高山姬鼠(Apodemus chevrieri)消化道形态是否存在差异,本研究于2012年6月分别对捕自云南剑川和呈贡地区高山姬鼠的体重、消化道(胃、小肠、大肠和盲肠)的长度以及含内容物重、去内容物重进行了测量。结果显示:剑川地区高山姬鼠的小肠和盲肠去内容物重以及长度均显著大于呈贡地区;体重、胃和盲肠含内容物重以及盲肠长均显著小于呈贡地区。结果表明:剑川地区的高山姬鼠具有较小的体重以减少对能量的绝对需求来应对该地较低的气温和食物资源短缺,暗示消化道形态的大小和消化能力的调整在自然环境高山姬鼠消化对策中起着重要的作用。
关键词
高山姬鼠,不同地区,消化道形态

1. 引言
小型脊椎动物通过自然选择,使其消化能力和吸收能力与环境相适应[1] 。动物消化道作为食物消化和吸收的重要场所,其形态结构与食物的质量及食物的可获得性密切相关[2] -[4] 。表型可塑性(phenotypic plasticity)是指某种基因型对外界不同环境条件产生不同表型的特征[5] 。消化器官表型变化在动物适应能量消耗增加或实物质量下降等方面占有重要地位[6] 。研究表明,小型动物主要通过改变消化道容量和消化道壁组织结构来调节其形态结构,改变消化道容量主要是通过改变消化道长度或管径,而消化道壁组织结构的变化主要包括微绒毛数量和管壁厚度的变化[7] -[9] 。
横断山脉地区是我国特有的高山峡谷地区,不仅哺乳动物种类丰富,而且特有种和古老种类比例高,被誉为“第四纪冰区动物的避难所”;同时,该地区特殊的动物地理条件使其成为举世瞩目的南北动物迁移和扩散通道[10] 。高山姬鼠(Apodemus chevrieri)属于鼠亚科,又名齐氏姬鼠,是典型的古北界种类[11] 。主要分布于我国西部高原,云南省主要分布于昭通、昆明、大理、丽江、澜沧江和怒江流域[12] 。关于高山姬鼠的相关研究已经很多,主要有光周期对高山姬鼠能量收支的影响[13] ,冷诱导最大代谢率的季节性差异[14] ,幼仔的生长发育和产热特征[15] ,冷驯化过程中的能量收支[16] 的研究等。本文以高山姬鼠为研究对象,对不同地区体重和消化道形态进行了比较,假设高山姬鼠能通过适当调节消化道形态来应对外界不同的环境,从而观察其在面对环境温度和食物质量不同时消化道形态的变化规律。
2. 材料与方法
2.1. 动物来源
实验动物于2012年6月分别捕自云南省剑川县石龙村(北纬26˚15'~26˚45',东经99˚40'~99˚55',海拔2590 m)的农田和云南省呈贡地区(北纬24˚32'~24˚55',东经102˚35'~102˚51',海拔1963 m)的灌木丛中。剑川县位于云岭山脉的中部(属横断山),年平均温度为−4℃。该地区干湿季节明显,变现出明显的温带季风气候特征。而呈贡地区属于热带半湿润季风气候,年平均气温14.5℃。两地相距137 km左右。剑川地区、呈贡地区两地分别捕获高山姬鼠10只,10只。
2.2. 动物处理
实验动物捕回后选择非繁殖成年个体作为实验样本,带回云南师范大学生命科学学院,称量体重、记录性别等,断颈处死,然后进行解剖。仔细将胃肠器官完全取出,分离出胃、小肠、大肠及盲肠,将各器官小心剔除肠系膜级其他组织,平展为最大长度,不要拉伸,然后测量各部分长度(精度1 mm)。用分析天平称量各器官的重量(精度为0.1 mg)。参照王德华等(2000)的方法,每个器官在滤纸上干燥后,放于锡箔纸上,称量其含内容物器官重;然后用解剖剪将器官纵切,用生理盐水充分冲洗内容物,用滤纸干燥,称量其去内容物器官重,即含内容物重。
2.3. 统计分析
采用SPSS 16.0进行实验数据的统计分析,数据经过正态分布和方差齐次性检验,符合参数检验条件。不同地区高山姬鼠消化道形态的差异采用独立样本t检验分析(Independent samples t-test),P < 0.05被认为差异显著,P < 0.01为差异极显著,文内数据均以平均值±标准差(Mean ± SE)表示。
3. 结果
3.1. 体重和总消化道
体重:剑川地区高山姬鼠体重显著小于呈贡地区(t = −3.465, P < 0.05)(表1)。总消化道:剑川地区高山姬鼠总消化道含内容物重(t = −5.275, P < 0.01),去内容物重(t = 2.944, P < 0.01)和长度(t = 8.290, P < 0.01)极显著大于呈贡地区(表1)。
3.2. 消化道各部分
胃:剑川地区高山姬鼠胃含内容物重显著小于呈贡地区(t = −4.022, P < 0.05),胃长度极显著小于呈贡地区(t = −5.742, P < 0.01)(图1、图2、图3)。小肠:剑川地区高山姬鼠小肠去内容物重显著性大于呈贡地区(t = 3.566, P < 0.05),小肠长度极显著大于呈贡地区(t = 6.314, P < 0.01)(图1、图2、图3)。大肠:剑川地区高山姬鼠大肠去内容物重显著大于呈贡地区(t = 2.811, P < 0.05)(图1、图2、图3)。盲肠:剑川地区高山姬鼠盲肠含内容物重极显著小于呈贡地区(t = −6.303, P < 0.01),盲肠长差显著大于呈贡地区(t = 2.456, P < 0.05)(图1、图2、图3)。
4. 讨论
4.1. 体重的变化
很多哺乳动物会通过降低体重来适应冷胁迫[17] [18] 。如草原田鼠[19] 和林姬鼠[20] 等。然而,有些小型哺乳动物在低温条件下会使其体重增加,如中缅树鼩[21] 。高山姬鼠的体重在剑川地区显著性的低于呈贡地区,这可能是因为剑川地区年平均气温较低,使得当外界环境温度较低时,高山姬鼠为了维持能

Table 1. Differences of the total digestive tract morphology of Apodemus chevrieri at different areas
表1. 不同地区高山姬鼠总消化道形态的差异
用独立样本t-检验的方法分析不同地区总消化道形态的差异,数值为平均值±标准误差,P < 0.05:差异显著,P < 0.01:差异极显著。
Note: Using independent samples t-test to analyze the significant variations of total digestive tract morphology at different areas, Data are means ± SE, P <0.05: significant differences。
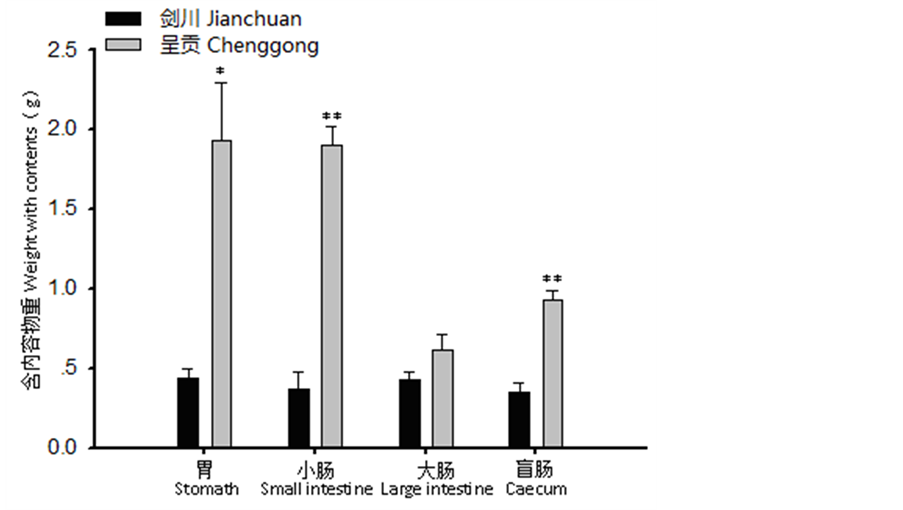
Figure 1. Comparison of the weight with contents of the digestive tract morphology in Apodemus chevrieri from different areas *P < 0.05, **P < 0.01
图1. 不同地区高山姬鼠各消化道含内容物重比较 *P < 0.05,**P < 0.01
量平衡,降低体重以减少能量的绝对需求。这一研究结果也与高山姬鼠冷诱导最大代谢率的季节性差异[14] 中结论一致。
4.2. 消化道总长度和重量及其适应意义
动物为了维持正常的消化生理活动,需要一定的能量支出,而消化系统的可塑性是许多小型哺乳动物在应对食物质量和数量变化时的适应策略之一[22] 。不同地区食物的丰富度和食物的构成均有很大的差异,对于高山姬鼠,这些外界压力对其消化道各器官提出了不同的要求,使其消化道形态发生相应的调整以适应能量需求的变化。本研究中剑川地区相对于呈贡地区温度较低,食物质量较差,面临的食物压
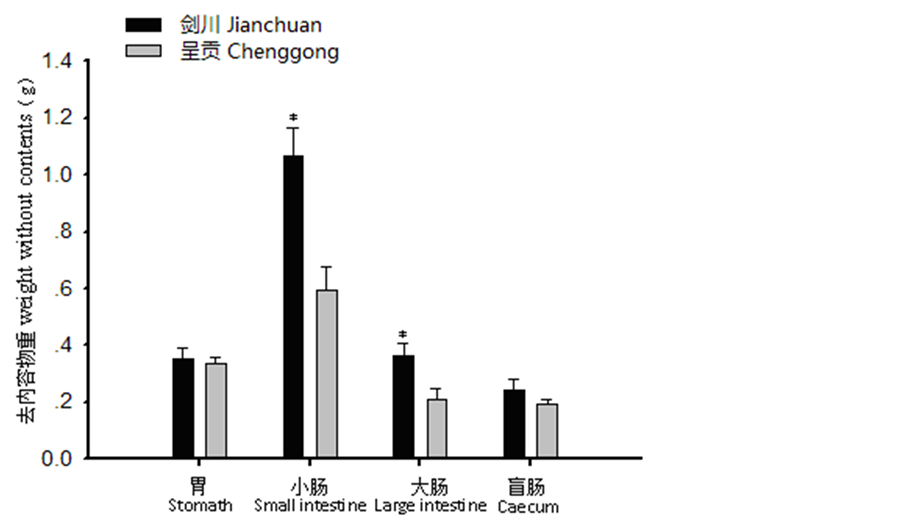
Figure 2. Comparison of the weight without contents of the digestive tract morphology in Apodemus chevrieri from different areas *P < 0.05, **P < 0.01
图2. 不同地区高山姬鼠各消化道去内容物重比较 *P < 0.05,**P < 0.01

Figure 3. Comparison of the length of the digestive tract morphology in Apodemus chevrieri from different areas *P < 0.05, **P < 0.01
图3. 不同地区高山姬鼠各消化道长度比较*P < 0.05,**P < 0.01
力增加,其通过增加消化道总长度和各消化道总去内容物重来增加能量摄入,从而抵御寒冷。
胃作为动物暂时储存食物并对食物进行初步消化吸收的产所[23] 。剑川地区高山姬鼠胃含内容物重显著小于呈贡地区,而去内容物重无显著差异,可能是因为呈贡地区温度较高,有较丰富的食物。小肠是食物消化和营养吸收的主要场所[24] 。在本研究发现,剑川地区高山姬鼠小肠含内容物重显著性小于呈贡地区,而长度和去内容物重剑川地区显著性大于呈贡地区,这可能是因为在剑川地区食物相对少的情况下,高山姬鼠通过增加小肠重量和长度以增加食物消化和营养吸收效率,以应对食物缺乏带来的影响。大肠是水分和离子重吸收的重要部位[25] 。剑川地区和呈贡地区大肠含内容物重和长度差异不显著,而去内容物重剑川地区显著性高于呈贡地区,表明了剑川地区食物质量,纤维素含量与呈贡地区相比存在一定差异性。盲肠作为纤维素的发酵场所[7] [8] [24] 。剑川地区高山姬鼠盲肠长度相对呈贡地区较大,含内容物重呈贡地区显著性大于剑川地区,表明剑川地区食物较少,且食物质量较差。
总之,高山姬鼠在云南呈贡地区和剑川地区消化道表型的变化特征,在一定程度上反映由于剑川地区纬度较高,气温较低,食物资源短缺,导致高山姬鼠通过增加消化道长度和重量来维持能量平衡。
基金项目
国家国际科技合作项目(2014DFR31040);十二五规划支撑项目(2014BAI01B00);国家自然科学基金项目(No. 31360096);云南省应用基础研究计划重点项目(No. 2013FA014)。
参考文献 (References)
- [1] Diamand, J.M. (1991) Evolutionary design of intestinal nutrient absorption enough but not too much. News Physiological Science, 6, 92-96.
- [2] Gross, J.M., Wang, Z. and Wunder, B.A. (1985) Effects of food quality and energy need Changes in gut morphology and capacities of Microtus chrogaster. Journal of Mammalogy, 66, 661-667.
- [3] Derting, T.L. and Bogue, B.A. (1993) Responses of the gut to moderate energy demands in a small herbivore (Microtus pennsylvanicus). Journal of Mammalogy, 74, 59-68.
- [4] 王德华, 王祖望 (2001) 高寒地区高原鼠兔消化道形态的季节动态. 动物学报, 47, 495-501.
- [5] Pigliucci, M. (2001) Phenotypic plasticity: Beyond nature and nurture. Johns Hopkins University Press, Baltimore, 125-128.
- [6] Derting, T.L. and Bogue, B.A. (1993) Responses of the gut to moderate energy demands in a small herbivore (Microtus pennsylvanicus). Canadian Journal of Zoology, 73, 243-252.
- [7] 沈丽, 王勇, 等 (2005) 洞庭湖不同生态类型区黑线姬鼠消化道重量和长度的季节性变化. 四川动物, 24, 132-137.
- [8] 柳劲松, 孙儒泳, 王德华 (2007) 三种锯齿动物的消化道形态特征. 动物学杂志, 42, 8-13.
- [9] 张美文, 王勇, 李波, 等 (2007) 洞庭湖区社鼠消化道长度和质量的季节变化. 生态学杂志, 26, 61-66.
- [10] 王政昆, 刘璐, 梁子卿, 等 (1999) 大绒鼠体温调节和产热特征. 兽类学报, 19, 276-286.
- [11] Corbet, G.B. (1978) The mammals of the Palaearctic region: A taxonomic review. British Museum (Natural History), London, 200-314.
- [12] 黄文几, 陈延熹, 温业新 (1995) 中国锯齿类. 复旦大学出版社, 上海, 126-127.
- [13] 李晓婷, 王蓓, 王政昆 (2011) 光周期对高山姬鼠能量收支的影响. 中国科技论文在线, 17, 1562-1565.
- [14] 朱万龙, 王蓓, 王政昆, 等 (2010) 高山姬鼠冷诱导最大代谢率的季节性差异. 师大学报, 2, 10-13.
- [15] 柳鹏飞, 张麟, 王政昆, 王睿, 朱万龙, 刘春燕, 孟丽华 (2010) 高山姬鼠幼仔的生长发育和产热特征. 兽类学报, 1, 45-50.
- [16] 王蓓, 徐伟江, 王政昆, 姜文秀, 练硝, 蔡金红, 尹锡全 (2007) 高山姬鼠冷驯化过程中的能量收支. 兽类学报, 4, 395-402.
- [17] Heldmaier, G., Steinlecher, S. and Ruf, T. (1989) Photoperiod and thermoregulation in vertebrate: Body temperature rhyme and thermgenic acclimation. Journal of Biological Rhythms, 4, 251-265.
- [18] Lovegrove, B.G. (2005) Seasonal thermoregulatory responses in mammals. Journal of Comparative Physiology B, 175, 231-247.
- [19] 王德华, 王祖望 (1990) 小型哺乳动物在高寒环境中的生存对策:Ⅱ高原鼠兔和根田鼠非颤抖性产热(NST). 兽类学报, 1, 40-53.
- [20] Corp, N., Gorman, M.L. and Speakman, J.R. (1999) Daily energy expenditure of free-living male wood mice in different habitats and seasons. Function Ecology, 13, 1365-2435.
- [21] 张武先, 王政昆 (2002) 冷驯化对中缅树鼩能量代谢的影响. 兽类学报, 2, 123-129.
- [22] 高文荣, 朱万龙, 王政昆 (2013) 禁食和重喂食对大绒鼠消化道长度与重量的影响. 动物学杂志, 4, 626-633.
- [23] Green, D.A. and Millarm, J.S. (1987) Changes in gut dimensions and capacity of Peromurscus maniculatus relative to diet quality and energy needs. Canadian Journal of Zoology, 65, 2159-2162.
- [24] 徐兴军, 吕建伟, 谢振丽 (2008) 嫩江下游人工林中黑线姬鼠与黑线仓鼠的消化道形态及其取食策略. 动物学杂志, 6, 131-136.
- [25] Green, D.A. and Millar, J.S. (1987) Changes in gut dimensions and capacity of Peromyscus maniculatus relative to diet quality and energy needs. Canadian Journal of Zoology, 65, 2159-2162.

NOTES
*通讯作者。