Advances in Applied Mathematics
Vol.05 No.02(2016), Article ID:17556,13
pages
10.12677/AAM.2016.52031
Modeling the Transmission of Densovirus (HaDNV-1) in the Helicoverpa armigera Population
Shuting Chen, Zhiming Guo
School of Mathematics and Information Science, Guangzhou University, Guangzhou Guangdong
Received: Apr. 23rd, 2016; accepted: May 10th, 2016; published: May 13th, 2016
Copyright © 2016 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



ABSTRACT
Densovirus (HaDNV-1) is a mutualistic symbiont of Helicoverpa armigera, and the infected females occupy advantage in vertical transmission. A model of species competition was posed to understand the influence of HaDNV-1. When the damage of HaDNV-1 is neglected, the stability of equilibria was analysed and a conclusion was drawn that the infected is dominant in Helicoverpa armigera population. Considering the harm of HaDNV-1, the model was modified by raising the mortality of the infected adult and larva respectively to indicate the cost; thereby the mortality was evaluated to lead to the infected pests extinct.
Keywords:Species Competition, Mutualistic Symbiont, Female Transmission Advantage, Stability

浓核病毒HaDNV-1在棉铃虫种群中传播的 数学模型
陈淑庭,郭志明
广州大学数学与信息科学学院,广东 广州
收稿日期:2016年4月23日;录用日期:2016年5月10日;发布日期:2016年5月13日

摘 要
HaDNV-1是一种与棉铃虫互利共生的浓核病毒,垂直传播时具有“雌性传播优势”机制。通过建立棉铃虫种群竞争模型,研究HaDNV-1在其种群内的传播情形。在不考虑HaDNV-1对棉铃虫的危害时,通过分析模型平衡点的稳定性得出感染型棉铃虫在竞争中占优势的结论。在考虑HaDNV-1对棉铃虫危害时,分别建立了棉铃虫造成成虫和幼虫死亡率增加的模型,并得到了使得感染型棉铃虫最终灭绝的成虫死亡率及幼虫死亡率的范围。
关键词 :种群竞争,互利共生,雌性传播优势,稳定性

1. 引言
棉铃虫是一种棉花蕾铃期的重要钻蛀性害虫,广泛分布在中国及世界各地。棉区和蔬菜种植区均有发生。棉铃虫主要蛀食蕾,花和铃,也取食嫩叶,危害十分严重。据相关统计,我国每年因棉铃虫造成的棉田损失达15%~20%,有些地区严重年份则高达50%以上,甚至绝收。因此,准确了解棉铃虫的生长规律和主要爆发期,掌握棉铃虫的种群动态性质,能够为农业生产部门的防治提供一定的理论指导。关于棉铃虫种群预测模型,许多学者运用统计学方法,通过收集某一地区历年来棉铃虫的爆发数据,结合软件分析影响种群的因素,得到种群预测模型 [1] - [4] 。例如,周志香 [1] 等在2000年建立了数据库,使用了灰色代数曲线,多项式回归和逐步回归的统计方法建立了棉铃虫的中、短期预报模型。刘硕 [2] 在2014年探讨了棉铃虫的危害因素,基于Logistic回归模型建立了棉铃虫的危害概率模型。
中国农业科学院吴孔明科研团队和英国兰开斯特大学(Lancaster University)威尔逊教授(Kenneth Wilson)科研团队合作研究发现了一种对寄主棉铃虫有利的浓核病毒(HaDNV-1)。该病毒不但使棉铃虫幼虫更健康,还可提高其对生物杀虫剂的抗性水平。相关研究成果于2014年发表于著名国际病理学专业期刊《公共科学图书馆:病原体(PLoS Pathogens)》上 [5] 。研究结果表明,棉铃虫感染该病毒后,幼虫和蛹的发育进度加快,成虫繁殖能力增强,对棉铃虫核型多角体病毒和Bt毒素的抗性提高。2008~2012年对不同地区棉铃虫自然种群的取样检测结果显示,野生棉铃虫的HaDNV-1平均感染率接近80% [6] 。
转Bt基因抗虫作物和生物农药(棉铃虫核型多角体病毒等)是控制棉铃虫发生危害的主要手段。HaDNV-1帮助棉铃虫发育得更健康,并提升其对Bt作物与生物杀虫剂的抗性,表明了自然生态系统物种关系的复杂性和生物防治方法遇到的挑战。该项研究成果是科学界对昆虫-病毒关系的新认知,对深入揭示农业生态系统物种关系协同进化机制,发展害虫防治新理论和新方法有重要科学意义 [6] 。
文献 [5] 的实验数据表明,病毒HaDNV-1进行垂直传播时,存在一种特殊的机制-雌性传播优势:即感染了病毒的雌性棉铃虫,后代全都是与生俱来携带病毒的棉铃虫。假设用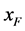 和
和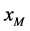 分别代表感染病毒的雌性棉铃虫和雄性棉铃虫,
分别代表感染病毒的雌性棉铃虫和雄性棉铃虫,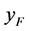 和
和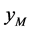 分别代表正常(无感染病毒)的雌性棉铃虫和雄性棉铃虫,那么HaDNV-1在棉铃虫种群中的垂直传播机制如表1所示。这种“雌性传播优势”有可能会使得HaDNV-1短时期内在棉铃虫种群内大面积爆发,从而使得对于感染病毒棉铃虫的防治越来越困难。
分别代表正常(无感染病毒)的雌性棉铃虫和雄性棉铃虫,那么HaDNV-1在棉铃虫种群中的垂直传播机制如表1所示。这种“雌性传播优势”有可能会使得HaDNV-1短时期内在棉铃虫种群内大面积爆发,从而使得对于感染病毒棉铃虫的防治越来越困难。
关于病毒在棉铃虫种群中传播的相关研究还可参考 [7] - [10] 等。例如,2005年Zhou [9] 等通过实验和野外收集数据,分析了HaSNPV-WT和HaSNPV-AaIT在棉铃虫种群中的水平和垂直传播动态,得到了影响病毒传播速率的因素。2006年Sun [10] 等则建立一个综合模型模拟棉铃虫种群某一阶段的病毒流行

Table 1. The vertical transmission of HaDNV-1
表1. HaDNV-1垂直传播
动态并分析其生物防治方案。
本文主要研究浓核病毒HaDNV-1在棉铃虫种群中的传播情况。在第一节中根据感染病毒的棉铃虫和正常棉铃虫的生长,发育以及繁殖特性,建立棉铃虫种群竞争的基础模型。在第二节中将研究第一节所建立的基础模型,应用常微分方程稳定性理论,通过分析基础模型平衡解的性态,得出HaDNV-1在棉铃虫种群中大范围传播而使得正常棉铃虫最终灭绝。第三节将根据文献 [5] 中观察得到的实际情况,考虑当感染HaDNV-1对棉铃虫造成一定危害时,对基础模型进行适当修正,得到使感染棉铃虫最终灭绝的条件以及两种棉铃虫稳定共存的条件。
2. 基础模型
文献 [5] 指出HaDNV-1水平传播时需要接触浓度较高的HaDNV-1溶液,即在棉铃虫种群内HaDNV-1实现水平传播的几率很小,因此可以忽略水平传播的影响,只考虑垂直传播的情形。令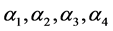 分别表示
分别表示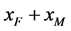 ,
,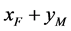 ,
,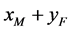 ,
,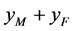 这四种交配方式后代的出生率,
这四种交配方式后代的出生率,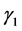 表示
表示 交配生出的后代为
交配生出的后代为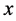 型(遗传了HaDNV-1病毒)的比例,则
型(遗传了HaDNV-1病毒)的比例,则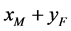 交配生出的后代为
交配生出的后代为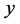 型的比例为
型的比例为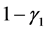 。令
。令 分别表示交配方式
分别表示交配方式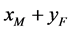 下,
下,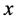 型后代雌性(
型后代雌性( )所占的比例和
)所占的比例和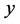 型后代雌性(
型后代雌性(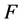 )所占的比例。
)所占的比例。 分别为
分别为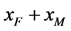 ,
,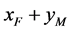 ,
,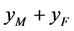 交配生出的后代中雌性(
交配生出的后代中雌性(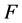 )所占的比例。根据随机交配的原则 [11] ,单个感染型雌性棉铃虫
)所占的比例。根据随机交配的原则 [11] ,单个感染型雌性棉铃虫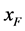 与感染型雄性棉铃虫
与感染型雄性棉铃虫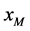 交配的概率等于单个正常雌性棉铃虫
交配的概率等于单个正常雌性棉铃虫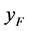 与感染型雄性棉铃虫
与感染型雄性棉铃虫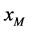 交配的概率,都是
交配的概率,都是 ;单个感染型雌性棉铃虫
;单个感染型雌性棉铃虫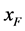 与正常雄性棉铃虫
与正常雄性棉铃虫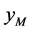 交配的概率等于单个正常雌性棉铃虫
交配的概率等于单个正常雌性棉铃虫 与正常雄性棉铃虫
与正常雄性棉铃虫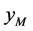 交配的概率,均为
交配的概率,均为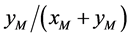 。记
。记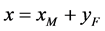 ,
,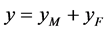 ,我们建立如下棉铃虫种群模型:
,我们建立如下棉铃虫种群模型:
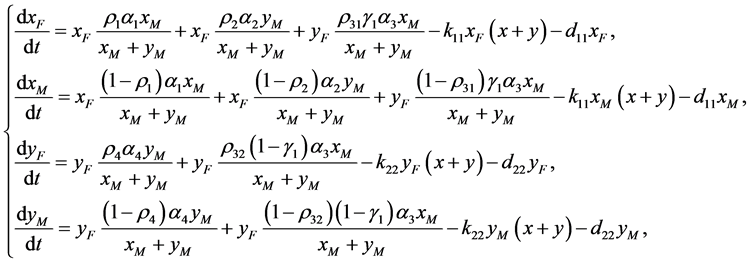 (2.1)
(2.1)
其中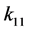 和
和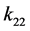 表示由密度制约和竞争引起的死亡率,
表示由密度制约和竞争引起的死亡率,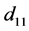 和
和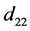 代表自然死亡率。由于性别染色体的随机搭配,不妨设雌性和雄性的比例为1:1,即
代表自然死亡率。由于性别染色体的随机搭配,不妨设雌性和雄性的比例为1:1,即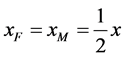 ,
,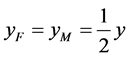 ,并且
,并且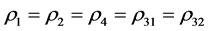 。因此把系统(2.1)的第一个方程和第二个方程相加,第三个方程和第四个方程相加则变为:
。因此把系统(2.1)的第一个方程和第二个方程相加,第三个方程和第四个方程相加则变为:
 (2.2)
(2.2)
系统(2.2)可改写成如下形式:
 (2.3)
(2.3)
其中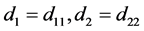
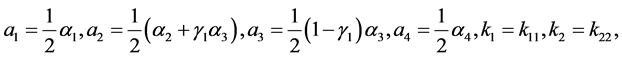 。
。
一般来说,棉铃虫的出生率与母体的体质相关。因此,在四种出生率中可假设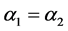 ,
,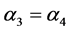 。由于感染病毒的棉铃虫比正常型棉铃虫具有更强的繁殖能力,从而有
。由于感染病毒的棉铃虫比正常型棉铃虫具有更强的繁殖能力,从而有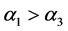 。实验表明
。实验表明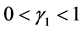 [5] 。因此有
[5] 。因此有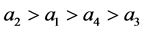 。因为感染棉铃虫比正常棉铃虫寿命更长,即在同等环境下,感染棉铃虫的自然死亡率更小,即
。因为感染棉铃虫比正常棉铃虫寿命更长,即在同等环境下,感染棉铃虫的自然死亡率更小,即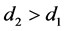 。此外,HaDNV-1帮助棉铃虫发育得更健康,并提升其对Bt作物与生物杀虫剂的抗性,说明感染棉铃虫对环境的适应度更大,即环境对于感染棉铃虫的威胁更小,故有
。此外,HaDNV-1帮助棉铃虫发育得更健康,并提升其对Bt作物与生物杀虫剂的抗性,说明感染棉铃虫对环境的适应度更大,即环境对于感染棉铃虫的威胁更小,故有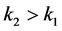 。由此建立了关于感染棉铃虫和正常棉铃虫的种群竞争基础模型。
。由此建立了关于感染棉铃虫和正常棉铃虫的种群竞争基础模型。
3. 系统(2.3)平衡点的稳定性
由于系统(2.3)在原点处没有定义,为了解原点及其附近邻域内解的性质,将系统(2.3)修正为:
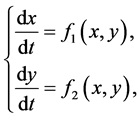 (3.1)
(3.1)
其中
显然, 是系统(3.1)的一个平衡点。由生物学意义,只需要考虑
是系统(3.1)的一个平衡点。由生物学意义,只需要考虑 的情况。由
的情况。由 ,有
,有
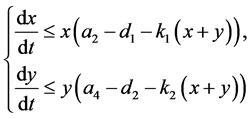
显然,如果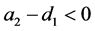 且
且 ,那么
,那么 ,易得如下结论:
,易得如下结论:
定理3.1:若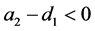 ,
,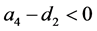 ,则系统(3.1)的平衡点
,则系统(3.1)的平衡点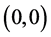 在
在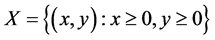 是全局稳定的。
是全局稳定的。
由系统(3.1),可以得到两个边界平衡点 和
和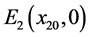 。如果
。如果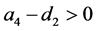 ,则系统(3.1)有边界平衡点
,则系统(3.1)有边界平衡点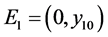 ,其中
,其中
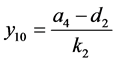 。
。
如果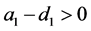 ,则系统(3.1)有边界平衡点
,则系统(3.1)有边界平衡点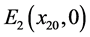 ,其中
,其中
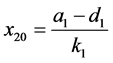 。
。
下面讨论 和
和 的稳定性。为此计算系统(3.1)的雅可比矩阵,
的稳定性。为此计算系统(3.1)的雅可比矩阵,
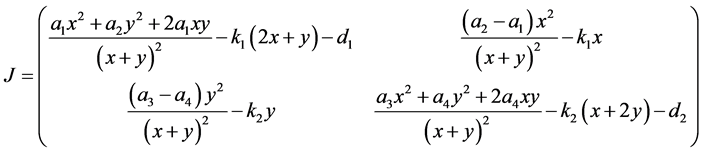 。
。
因此,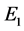 的特征方程为
的特征方程为
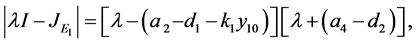
这时 。考虑上述特征方程的另一个根
。考虑上述特征方程的另一个根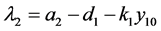 。由模型系数的关系知
。由模型系数的关系知
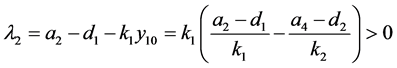 。
。
因此,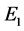 是一个鞍点。同样,可以得到
是一个鞍点。同样,可以得到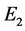 的特征方程如下:
的特征方程如下:
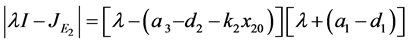 。
。
该方程的一个特征根为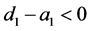 ,其另一个根为
,其另一个根为
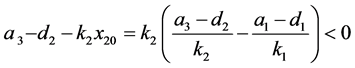 。
。
故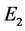 是稳定的结点。
是稳定的结点。
接下来考虑正平衡点的存在性。令
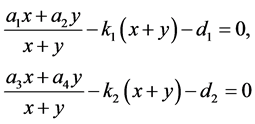
于是有
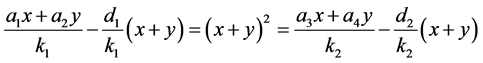 。
。
把上面等式两端进行化简可以得到,
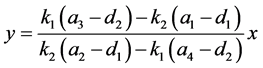 。
。
由于
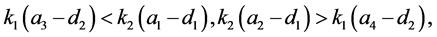
故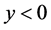 。即系统(3.1)不存在正平衡点。
。即系统(3.1)不存在正平衡点。
由上面的讨论我们得到如下定理:
定理3.2:(i) 当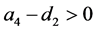 时,系统(3.1)存在边界平衡点
时,系统(3.1)存在边界平衡点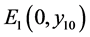 ,且
,且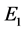 是一个鞍点;
是一个鞍点;
(ii) 当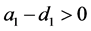 时,系统(3.1)存在边界平衡点
时,系统(3.1)存在边界平衡点 ,且
,且 是一个局部渐近稳定的结点;
是一个局部渐近稳定的结点;
(iii) 系统(3.1)不存在正平衡点。
注:由定理3.2可知,感染型棉铃虫在竞争中是占优势的,极容易使得病毒HaDNV-1在棉铃虫种群中快速地传播开来。这显然得益于它良好的生长发育能力,对Bt作物和杀虫剂的抗性以及病毒HaDNV-1的雌性传播优势机制。2008~2012年对不同地区棉铃虫自然种群的取样检测结果显示,野生棉铃虫的HaDNV-1平均感染率接近80% [6] ,说明HaDNV-1在棉铃虫种群内大范围传播,这与我们所建立的模型是相吻合的。
接下来证明系统(3.1)的有界性。
定理3.3:系统(3.1)带非负初值的解最终一致有界。
证明:显然,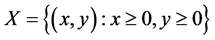 是系统(3.1)的正不变集。令
是系统(3.1)的正不变集。令
 。
。
则
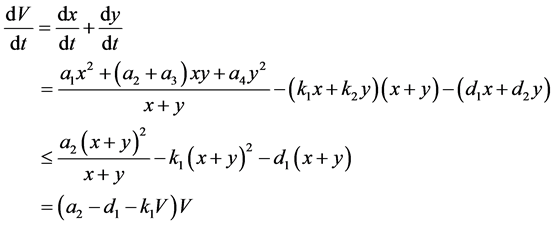
因此,对足够大的 ,有
,有
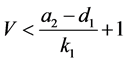
故系统(3.1)带非负初值的解最终一致有界,证毕。
从定理3.2和定理3.3可知,当 和
和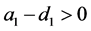 同时满足,系统(3.1)只存在三个平衡点
同时满足,系统(3.1)只存在三个平衡点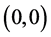 ,
,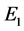 和
和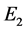 ,且
,且 是不稳定的结点,
是不稳定的结点,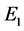 为鞍点,
为鞍点,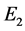 为稳定的结点。又由系统(3.1)最终一致有界可知,在
为稳定的结点。又由系统(3.1)最终一致有界可知,在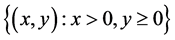 的解最终都趋于
的解最终都趋于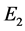 。
。
如果仅大量地采用转Bt基因抗虫作物和生物农药(棉铃虫核型多角体病毒)进行棉铃虫防治,由于感染型棉铃虫对Bt毒素和棉铃虫核型多角体病毒具有抗性而不会造成数量的大幅度减少,正常棉铃虫则在这两种防治措施下使得出生率减小而死亡率增大,最终会导致整个棉铃虫种群都携带HaDNV-1,从而这两种防治措施都会失效。
4. 考虑HaDNV-1危害的模型
文献 [5] 指出,2008-2012年研究人员对若干个地区的野外棉铃虫种群进行了抽样检测,得到连续五年HaDNV-1的感染率:2008年为87.1%,2009年为81.2%,2010年为76.8%,2011年为68%,2012年为67%。数据显示尽管HaDNV-1感染率较高,但呈稳定下降的趋势,这说明HaDNV-1感染对棉铃虫可能存在一些不确定的代价。一些未知的竞争因素,种群密度,微生物干扰等都有可能成为这种不确定代价的原因。一种可能的情况是感染HaDNV-1的棉铃虫一开始并不会出现不利于自身生长发育的情况,但在垂直传播了若干代后病毒的危害逐渐显现,而使后代感染HaDNV-1的棉铃虫付出一定的代价。
考虑HaDNV-1的危害在棉铃虫成虫期出现,造成棉铃虫成虫的死亡率增加。假设感染型棉铃虫成虫死亡率增加了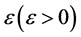 ,那么系统(2.3)变为
,那么系统(2.3)变为
 (4.1)
(4.1)
显然,当 时
时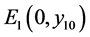 仍是系统(4.1)的一个平衡点。当
仍是系统(4.1)的一个平衡点。当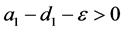 时,存在平衡点
时,存在平衡点 ,其中
,其中
 。
。
 的特征方程为
的特征方程为
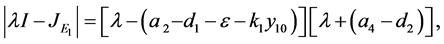
因此,
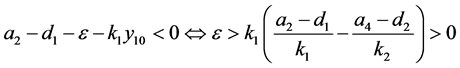
同样,可以算出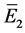 的特征方程为
的特征方程为
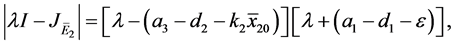
则
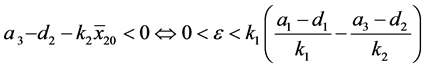
定理4.1:(i) 假设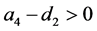 。若
。若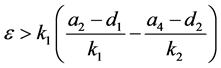 ,则
,则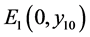 是局部渐近稳定的;
是局部渐近稳定的;
(ii) 假设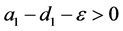 。若
。若 ,则
,则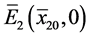 是局部渐近稳定的。
是局部渐近稳定的。
下面讨论正平衡点的存在性和稳定性。类似于系统(3.1)计算正平衡点的方法,可以得到
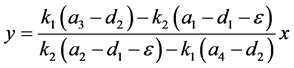
记

解得
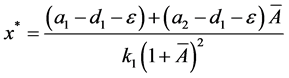
为保持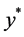 的正性,要求
的正性,要求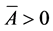 ,即
,即
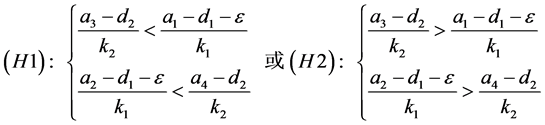
(H1)成立等价于

而(H2)成立等价于
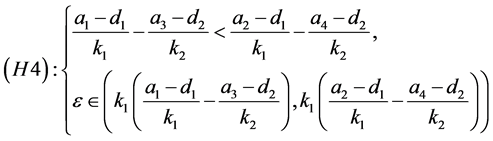
由于
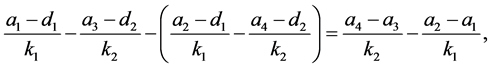
由模型的系数关系知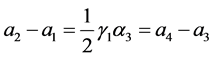 。又
。又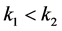 ,故
,故
 (4.2)
(4.2)
因此(H3)不成立。由(H4)可知,当
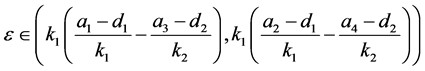
时,系统(4.1)存在唯一的正平衡点 ,其中
,其中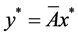 。
。
下面讨论 的稳定性。根据雅可比矩阵得到
的稳定性。根据雅可比矩阵得到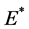 的特征方程为
的特征方程为

其中
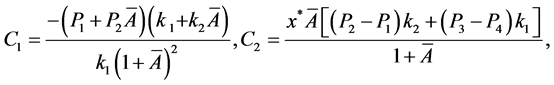
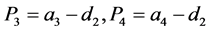
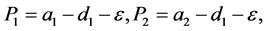 。
。
显然, ;又由(4.2)知
;又由(4.2)知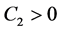 。因此特征方程的根均具有负实部。故可得以下结论:
。因此特征方程的根均具有负实部。故可得以下结论:
定理4.2:若
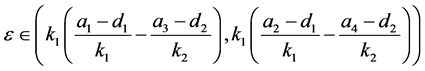
满足,则系统(4.1)存在唯一的正平衡点 ,且
,且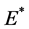 是稳定的结点。
是稳定的结点。
根据定理4.1和定理4.2,系统(4.1)平衡解的性态如表2所示。
从表2可以看出,若HaDNV-1的危害使得感染型棉铃虫成虫的死亡率增加,会使得HaDNV-1在棉铃虫种群中的传播情况发生变化。当感染棉铃虫成虫的死亡率增加量属于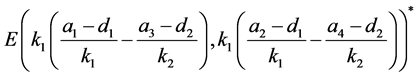 时,感染HaDNV-1的棉铃虫数量比例会逐渐下降,使得两种棉铃虫达到一个平衡。当感染棉铃虫成虫的死亡率增加量属于
时,感染HaDNV-1的棉铃虫数量比例会逐渐下降,使得两种棉铃虫达到一个平衡。当感染棉铃虫成虫的死亡率增加量属于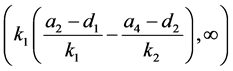 时,感染棉铃虫会最终灭绝。
时,感染棉铃虫会最终灭绝。
考虑HaDNV-1的危害在棉铃虫幼虫期出现,造成感染HaDNV-1棉铃虫幼虫死亡率增加。假设感染棉铃虫幼虫和正常棉铃虫幼虫的死亡率分别为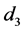 和
和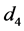 。由于HaDNV-1的危害,有
。由于HaDNV-1的危害,有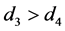 。令
。令 表示棉铃虫从出生成长到成虫所需要的时间,那么感染棉铃虫和正常棉铃虫从幼虫成长到成虫的存活率分别为
表示棉铃虫从出生成长到成虫所需要的时间,那么感染棉铃虫和正常棉铃虫从幼虫成长到成虫的存活率分别为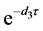 和
和 。因此,考虑如下棉铃虫成虫增长模型:
。因此,考虑如下棉铃虫成虫增长模型:
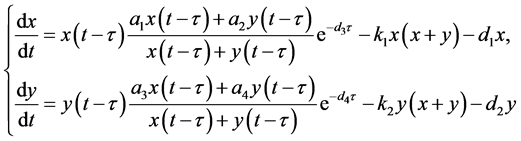 (4.3)
(4.3)
类似于系统(3.1)在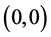 处补充定义,显然
处补充定义,显然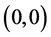 始终是平衡点。容易算得系统(4.3)的平衡点。当
始终是平衡点。容易算得系统(4.3)的平衡点。当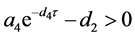 时,存在平衡点
时,存在平衡点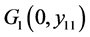 ,其中
,其中
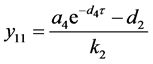 。
。
当 时,存在平衡点
时,存在平衡点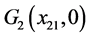 ,其中
,其中
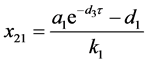 。
。
下面讨论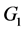 和
和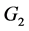 的稳定性。在
的稳定性。在 处的特征方程为
处的特征方程为
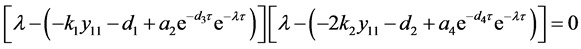 。
。
Table 2. The type of equilibria of system (4.1)
表2. 系统(4.1)平衡点的类型
根据文献 [12] 的定理4.7,可知

的根均具有负实部。考虑方程
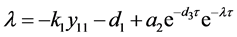
把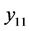 代入可知,上式的根均具有负实部的充分条件是
代入可知,上式的根均具有负实部的充分条件是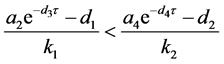 。
。
在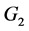 处的特征方程为
处的特征方程为

上式的根均具有负实部的充分条件是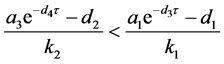 。
。
定理4.3:(i) 假设 。若
。若
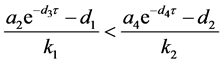 ,
,
则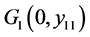 局部渐近稳定;
局部渐近稳定;
(ii) 假设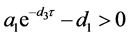 。若
。若
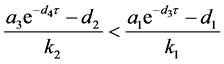 ,
,
则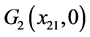 局部渐近稳定。
局部渐近稳定。
下面分析系统(4.3)正平衡点的存在性。令
可以算得,

 等价于
等价于
把 代入系统(4.3)可得,
代入系统(4.3)可得,
由(H5)有
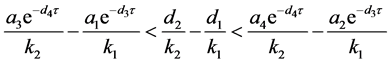
又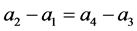 ,因此
,因此

所以(H5)等价于
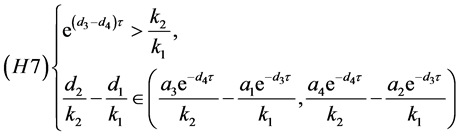
同理,(H6)等价于
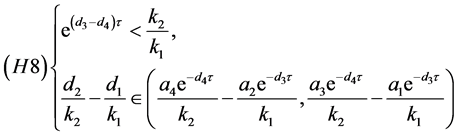
定理4.4:若(H7)和(H8)之一成立,则系统(4.3)存在唯一的正平衡点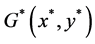 。
。
下面讨论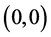 ,
,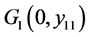 和
和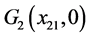 的全局稳定性。
的全局稳定性。
引理4.1 [13] :给定方程

假设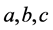 和
和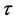 均为大于0的常数,则有
均为大于0的常数,则有
(i) 如果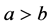 ,则
,则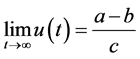 ;
;
(ii) 如果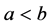 ,则
,则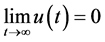 。
。
定理4.5:(i) 假设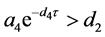 。若
。若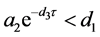 ,则
,则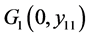 全局渐近稳定;
全局渐近稳定;
(ii) 假设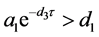 。若
。若 ,则
,则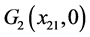 全局渐近稳定;
全局渐近稳定;
(iii) 若 和
和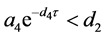 ,则
,则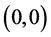 全局渐近稳定。
全局渐近稳定。
证明:令

当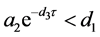 时,由引理(4.1)知当
时,由引理(4.1)知当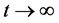 时,
时,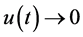 。由于
。由于
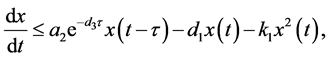
故当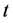 足够大时有
足够大时有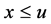 。即当
。即当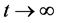 时,
时,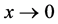 。根据渐近自治系统理论,有
。根据渐近自治系统理论,有
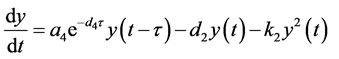
当 时,由引理(4.1)知当
时,由引理(4.1)知当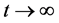 时,
时,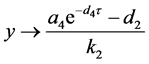 。因此,当
。因此,当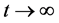 ,有
,有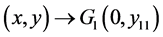 。
。
(ii)和(iii)同样用引理(4.1)可以证得,证毕。
定理4.5的(i)表明,当HaDNV-1的危害使得感染棉铃虫和正常棉铃虫的幼虫死亡率满足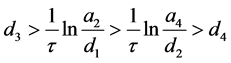 时,整个棉铃虫种群携带HaDNV-1的个体比例会下降,并使得感染棉铃虫最终会灭亡。
时,整个棉铃虫种群携带HaDNV-1的个体比例会下降,并使得感染棉铃虫最终会灭亡。
5. 总结与讨论
本文主要研究病毒HaDNV-1在棉铃虫种群中传播的情形。在第一节中根据感染棉铃虫和正常棉铃虫的生长发育,繁殖特性和垂直传播的特殊机制,建立起棉铃虫种群竞争模型。第二节主要讨论了棉铃虫种群竞争模型平衡点的存在性和局部稳定性,得到感染棉铃虫在竞争中占优势的结论,与现实检测结果相符合。第三节根据现实检测到棉铃虫感染HaDNV-1的比例逐年下降,考虑是由于HaDNV-1在传播了若干代后对棉铃虫产生一定危害,分别建立了危害造成感染棉铃虫成虫和幼虫的死亡率增加的模型,并讨论了使得感染棉铃虫比例下降甚至最终灭绝的成虫死亡率及幼虫死亡率范围。但在考虑危害造成棉铃虫幼虫死亡率增加的系统(4.3)中,暂时没有得出正平衡点稳定的条件。此外,由于感染HaDNV-1的棉铃虫幼虫和蛹发育的速度比正常棉铃虫快,即从幼虫成长到成虫的时间比正常棉铃虫短,因此可以在系统(4.3)考虑两个时滞,分别表示感染棉铃虫和正常棉铃虫的成长时间,再做进一步的讨论。
基金项目
本文得到国家自然科学基金(11371107)与教育部博士点基金(20124410110001)资助,在此表示感谢!
文章引用
陈淑庭,郭志明. 浓核病毒HaDNV-1在棉铃虫种群中传播的数学模型
Modeling the Transmission of Densovirus (HaDNV-1) in the Helicoverpa armigera Population[J]. 应用数学进展, 2016, 05(02): 242-254. http://dx.doi.org/10.12677/AAM.2016.52031
参考文献 (References)
- 1. 周志香, 王志伟, 刘文平, 等. 棉铃虫综合因素预报模型研究[J]. 中国农业气象, 2000, 21(4): 38-43.
- 2. 刘硕, 吴凤祥. 棉铃虫预测模型与模拟研究[D]: [硕士学位论文]. 保定: 河北农业大学, 2014.
- 3. 高增祥, 徐汝梅, 吴子江, 等. 基于生命表参数的棉铃虫种群动态研究[J]. 昆虫学报, 2005, 48(4): 568-575.
- 4. 苏战平, 张孝羲, 翟保平. 江苏棉区第5代棉铃虫种群动态的模拟及预测[J]. 昆虫学报, 2002, 45(4): 465-470.
- 5. Xu, P., Liu, Y., Graham, R.I., et al. (2014) Densovirus Is a Mutualistic Symbiont of a Global Crop Pest (Helicoverpa armigera) and Protects against a Baculovirus and Bt Biopesticide. PLoS Pathogens, 10, e1004490. http://dx.doi.org/10.1371/journal.ppat.1004490
- 6. 农业部网. 中国农科院专家发现棉铃虫互利共生新病毒[EB/OL]. http://news.xinhuanet.com/politics/2014-11/03/c_127173112.htm, 2014-11-03.
- 7. Georgievska, L., De Vries, R.S., Gao, P., et al. (2010) Transmission of Wild-Type and Recombinant HaSNPV among Larvae of Helicoverpa armigera (Lepidoptera: Noctuidae) on Cotton. Environmental Entomology, 39, 459-467. http://dx.doi.org/10.1603/EN09183
- 8. Sun, X., Chen, X., Zhang, Z., et al. (2002) Bolloworm Responses to Re-lease of Genetically Modefied Helicover armigera Nucleopolyhedroviruses in Cotton. Journal of Invertebrate Pathology, 81, 63-69. http://dx.doi.org/10.1016/S0022-2011(02)00144-1
- 9. Zhou, M., Sun, X.L., Sun, X.C., et al. (2005) Horizontal and Vertical Transmission of Wild-Type and Recombinant Helicoverpa armigera Single-Nucleocapsid Nucleopolyhe-drovirus. Journal of Invertebrate Pathology, 89, 165-175. http://dx.doi.org/10.1016/j.jip.2005.03.005
- 10. Sun, X., van der Werf, W., Bianchi, J.J.A., et al. (2006) Modeling Biological Control with Wild-Type and Genetically Modified Baculoviruses in the Helicoverpa armigera-Cotton System. Ecological Modelling, 198, 387-398. http://dx.doi.org/10.1016/j.ecolmodel.2006.05.011
- 11. Li, J. (2008) Differential Equations Models for Interacting Wild and Transgenic Mosquito Populations. Journal of Biological Dynamics, 2, 241-258. http://dx.doi.org/10.1080/17513750701779633
- 12. Smith, H. (2010) An Introduction to Delay Differential Equ-ations with Applications to the Life Sciences. Texts in Applied Mathematics, Vol. 57. Springer, New York.
- 13. Liu, S. and Chen, L. (2002) Extinction in Competition and Permanence in Competitive Stage-Structure System with Time Delay. Journal of Mathematical Analysis & Applications, 51, 1347-1361.