Geographical Science Research
Vol.3 No.04(2014), Article
ID:14334,8
pages
DOI:10.12677/GSER.2014.34007
Study on the Ecological Habits of Main Recent Ostracodes in Fuxian Lake
College of Tourism and Geography Science, Yunnan Normal University, Kunming
Email: 564378309@qq.com
Copyright © 2014 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received: Oct. 15th, 2014; revised: Nov. 1st, 2014; accepted: Nov. 7th, 2014
ABSTRACT
The paper describes the species and ecological characteristics of the Modern Ostracodes in the Lake Fuxian, through the field acquisition of the lake surface sediment and related environment. It mainly describes the Physocupria kraelini, Heterocypris incongruens, Candonocypris noveazelandiae, Parachinocythere reticulate gen.et sp.nov to further enrich the Modern Ostracodes ecosystem in Yunnan area and play a foreshadowing role to improve the research level of the Yunnan area of earth science work.
Keywords:Ostracods, Ecological Habits, Fuxian Lake

抚仙湖主要现代介形虫生态习性研究
刘姗姗,谢曼平
云南师范大学旅游与地理科学学院,昆明
Email: 564378309@qq.com
收稿日期:2014年10月15日;修回日期:2014年11月1日;录用日期:2014年11月7日

摘 要
本文主要通过野外采集的抚仙湖表层沉积物及相关湖泊环境信息,对该湖泊中现代介形虫种类及其生态特征进行了研究。主要描述了抚仙湖中克氏瘤丽星介Physocupria kraelini、非调和异星介Heterocypris incongruens、泽氏玻星介Candonocypris novaezelandiae、鹤庆盆地新属种Parachinocythere reticulata gen. et sp. nov.进一步充实云南地区现代介形虫的生态资料,为今后将介形虫这一替代指标用于古环境古气候的恢复工作中,提高云南地区地球科学的研究水平等工作起铺垫作用。
关键词
现代介形虫,生态习性,抚仙湖

1. 引言
湖泊沉积中的微体生物对环境变化具有非常敏感的响应因此是重建环境演变和生态特征的主要生物指标,其中的介形类作为一类最重要的生物组合得到了广泛的研究[1] 。介形虫是一种水生双壳微体甲壳类动物,壳为碳酸盐。介壳作为湖泊沉积中广泛存在并且保存较好的微体化石,可以提供重要的环境变化信息,越来越多地受到环境变化研究者们的重视[2] 。介形虫分布的时间范围覆盖寒武纪到现在。在整个地质时期,介形虫从绝对的海洋环境逐步侵入到大部分水体环境中,并覆盖了很广的盐度和温度范围,包括暂时性的湖泊和池塘。有些种局限在特殊的生境内,因而可以作为指示古环境的有效指标[3] 。在区域详细的生态与湖泊学调查基础上,获得各种介形虫对各环境参数(温度、盐度、pH值、水深等)的最佳适应范围和生存忍耐极限,通过数理统计的方法,建立定量的种属组合与环境参数的转换函数,可以定量地重建古环境参数[4] 。几十年来,我国在中、新生代陆相介形虫化石研究领域已取得一系列成果。对比之下,我国对现生陆相介形虫的研究比较落后。通过对现生介形虫生态分布与环境关系的了解,可以更准确地解释化石介形虫的环境意义[5] 。
对亚洲湖泊的现代介形类的生态习性进行研究,可以丰富和加强古北区的第四纪生态环境研究手段。到目前为止,国内对现代介形进行研究的工作进展非常缓慢,主要集中在中国东南部地区,比如太湖、厦门等地。而对于相对偏远的西南地区,鲜少涉及。上个世纪八十年代,杨留法和黄保仁对抚仙湖25个表层沉积物中的介形进行了鉴定分类,共发现4科,10属,15种[6] 。其中14个都是新种,并且只提供了照片,没有进行进一步的介壳形态描述。这给后继研究者的借鉴意义不大。本文对抚仙湖现生介形几个种类进行了详细描述,旨在拓展抚仙湖现代介形虫的生态习性研究这一方向,为今后的工作起铺垫作用。
2. 抚仙湖环境描述
抚仙湖位于云南省玉溪市东北部,是云南三大高原湖泊之一,系断层陷落湖,经纬度范围是(24˚13'N~24˚46'N,102˚397'E~103˚00'E) [7] 。湖泊南北长31.5 km,最宽处11.5 km、最窄处3.2 km,呈狭长状葫芦型,是我国已探明的第二深水湖泊。最大水深151.5 m,平均水深87 m,容水量185 × 108 m3,占云南省九大高原湖泊蓄水量的66.6%,湖面面积212 km2。东、南、西三面山岭迫近湖岸,仅北部地势平坦开阔,是一个半封闭的外流湖泊[8] 。抚仙湖属于亚热带季风气候,降水受西南和东南季风影响[9] 。湖泊积水主要靠降雨补给,降水主要集中在6~8月,一年最高水位多出现在9~12月,最低水位出现在5月或者6月[10] 。在西南面有1条宽5.6 m、长2.2 km的隔河与星云湖相连,接纳星云湖来水,东岸的海口河是其唯一的出口,与南盘江上游相连[11] ,目前由于抚仙湖水位下降,基本已经成为一个内陆封闭湖泊(见图1)。
近年来随着工农业的发展及人口的增加,星云湖水体严重污染,水质下降为IV类水体,湖泊富营养
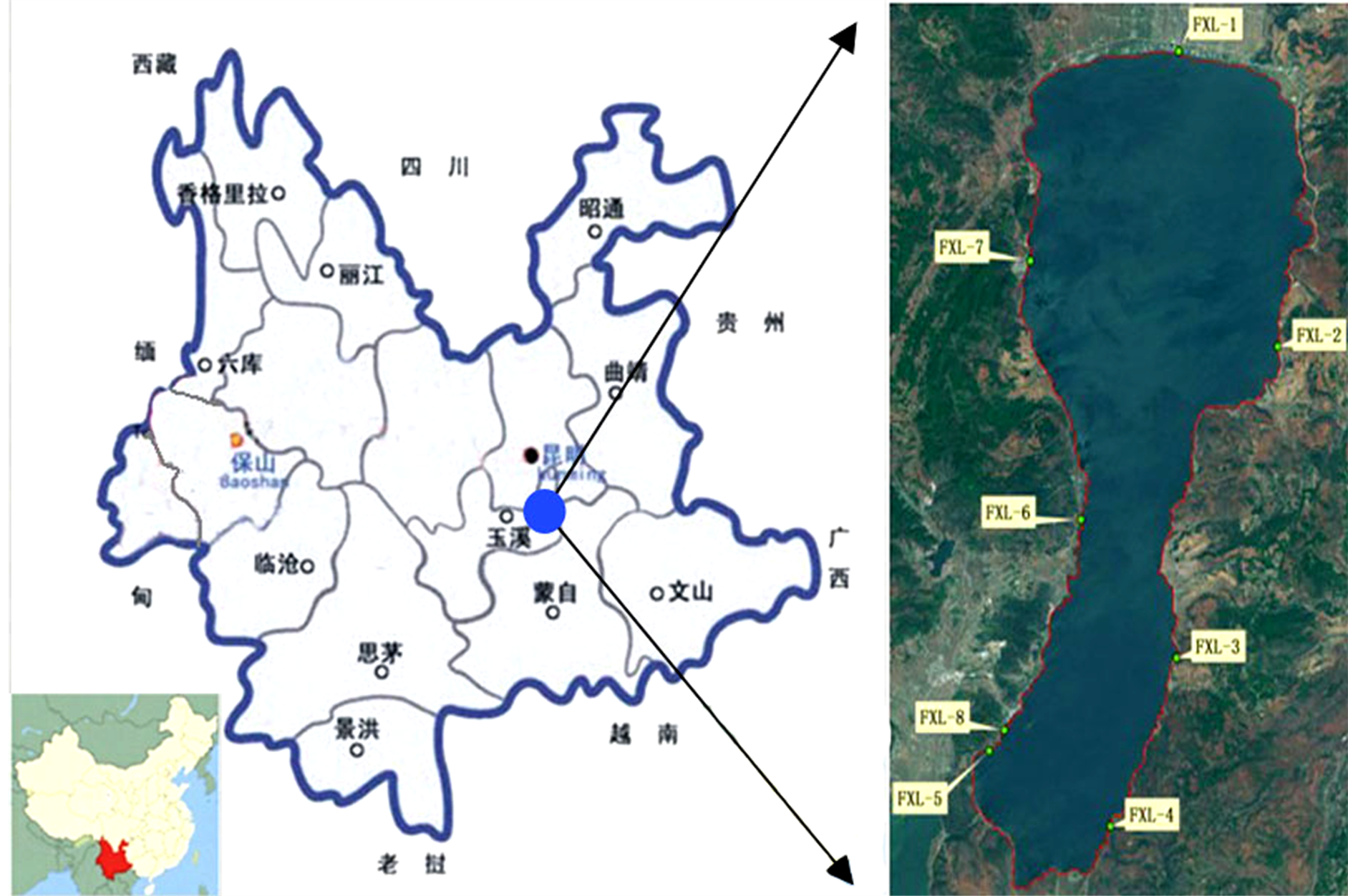
Figure 1. The position of Fuxianhu Lake and the Modern Ostracode sample sites of Fuxian Lake
图1. 抚仙湖位置及现代介形虫采样点标记示意图
化程度日趋加剧,达到中富营养型。星云湖通过隔河补充到抚仙湖的湖水在抚仙湖南部造成明显的水质污染,严重危及旅游环境[12] 。抚仙湖目前基本处于贫营养状态,但水质呈下降趋势[13] 。
3. 采样方法和样品处理方法
3.1. 野外采样
我研究小组分别于2012年6月中旬、7月中旬、8月中旬,对抚仙湖周边浅水部分进行定期的监测及采样。获得了一些现代介形样品。并对各个采样点做了简单的描述(见表1)。
3.2. 实验室样品处理
从冰箱中取出所采集的样品,分别放入已标好序号的塑料杯中解冻,称取适当的样品重量,用一套(65 μm,125 μm,250 μm)筛子分选,然后一层一层分别用去离子水冲洗装入不同的塑料杯中,直接用吸管将见到的介形虫吸入指形管中。分别从每个样品的65 μm筛子中取样品于玻璃器皿中,在徕卡(Laika)电子显微镜下观察、鉴定。对于现代介形虫,在体视显微镜下用毛笔挑出介形虫,并用解剖针对附肢进行分离处理,依次观察介形虫附肢的形态特征并进行分类鉴定与研究,用徕卡(Laika)电子显微镜照片采集器进行图片采集,获得大量图片资料。发现所获样品中有克氏瘤丽星介Physocupria kraelini、非调和异星介Heterocypris incongruens、泽氏玻星介Candonocypris novaezelandiae、鹤庆盆地新属种Parachinocythere reticulata gen. et sp. nov. [14] 。其中鹤庆盆地新属种在滇中地区还没有发现过,在禹娜所描述滇池5种介形虫中也没有[15] [16] ,这介形虫组合与滇池的大有不同。

Table 1. Sample data of Fuxian Lake
表1. 抚仙湖样品采集数据
4. 主要现代介形虫种类及其生态特征
4.1. 克氏瘤丽星介Physocypria kraepelini
1903 Physocypria kraepelini Muller, Muller, p.165, figs. 1-7 2000 Physocypria kraepelini Muller, Meisch, p. 227, fig. 97.
描述:壳体小而扁,壳壁薄。侧视近于半圆形,后中部最高,近腹部最长;背缘中部圆形拱起;腹缘中部微内凸;前缘圆,较低;后缘高,宽圆。背视凸度小,前后两端窄圆,两侧弧形外弯,后中部最厚。左瓣稍大,腹部叠覆明显,并沿前、腹缘端超出右瓣;右瓣沿背缘稍微高出左瓣。壳面光滑;活体壳体透明,浅黄色,在前部、后部和中背部表面具棕褐色斑。右瓣沿前、后腹缘具一列小瘤(见图2)。
度量(mm):长0.52~0.60,高0.35~0.43,厚0.22~0.27。
分布:广布于欧亚大陆。本次发现此种普遍地分布在整个湖盆四周的浅水地带,分布水深通常在50米以内,尤其分布在25米以内。
4.2. 非调和异星介Heterocypris incongruens
美星介亚科Cyprinotinae Bronshtein, 1947
异星介属Heterocypris Claus, 1892
非调和异星介Heterocypris incongruens (Ramdohr, 1808) (图2)
2000 Heterocypris incongruens, Meisch, p. 346, figs. 145 (A-G),146 (A-I)。
2003 Heterocypris incongruens, Mischke et al., p. 289, pl. 2, fig. 6。
2006 Heterocypris incongruens, 张玲等,430页,图版I,图10。
2010 Heterocypris incongruens, Lietal., p. 885, pl. 1, fig. 10。
描述:前部通常是宽圆,有时陡圆,后部宽圆;在两端左瓣轻微超覆右瓣。壳体表面比较光滑,但是两瓣的前面区域都很小,其次管理微刺(在背视图看最好,见图3)。扫描电镜照片说明这些小刺是通过不对称的增厚的嘴唇状的小孔通道形成的。左瓣在腹部超覆右瓣。右瓣外部的边缘地带前部和后部都伴

Figure 2. Physocypria kraepelini
图2. Physocypria kraepelini形状图

Figure 3. Electron micrograph of Heterocypris incongruens, female
图3. Heterocypris incongruens显微镜电子图,雌性
随有一排瘤。这些瘤在数量上和外型上多变,可能是尖锐的也可能是宽圆的,有时较弱或不明显。(Heterocypris incongruens软体见图4)。
大小:雌性的大小为1.2~1.9 mm (通常1.4~1.6 mm),雄性1.2~1.3 mm。
颜色:黄色至红褐色,有时为青黄色或者棕色。
分布:喜欢浅季节性的水池和小的永久性的水体,在师大图书馆南面人工湖中也有发现,数量多。偏爱基质是粘土和缺乏大型水生植物的环境。此物种在富营养化程度高、低氧的池塘中大量繁殖(如云南滇池、),例如那些被水鸟产生的有机物质高度污染的水体。在小河和江的安静区域、稻田、有时在泉眼和地下水域也有发现。
4.3. 泽氏玻星介Candonocypris novaezelandiae
爬星介亚科Herpetocypridinae Kaufmann, 1900
玻星介属Candonocy pris s¥ars, 1896
泽氏玻星介Candonocypris novaezelandiae (Baird, 1843) (图3)
1975 Candonocypris assimilis, Okubo, 1975, p. 157, fig. 1, pl. 1, figs. A) N。
1981 Candonocypris novaezelandiae, De Dekker, p. 53, figs. 6, 7。
1994 Candonocypris novaezelandiae, Eagar, p. 79, fig. 7。
度量(mm):长1.63~1.77,高0.74~0.81。
描述:壳体呈绿色。右壳前、后两端比左壳长,尤其在前端最为显著;左壳在腹部微叠覆于右壳。侧视长卵形,中部偏后最高;背缘中部弧形隆起;前端宽圆,后端窄圆;腹缘中部明显内弯。背视壳体狭长,最宽处位于中后部;两端皆尖,前端更甚。壳面光滑。前部内板宽,约为壳长的1/5;后部内板较窄,宽约为前部的1/2。前、后庭发育;结合带窄;放射毛细管短、直,数目中等。左壳织边弱,沿外缘延伸。右壳内板上具独特的粗脊状织边,在前部位于内板的中部,在腹部沿外缘延伸,在后部位于内板靠内一侧(见图5)。
 (a)
(a) (b)
(b) (c)
(c) (d)
(d) (e)
(e) (f)
(f)
Figure 4. Heterocypris incongruens, software. (a) Cleaning leg; (b) First antenna; (c) Walking leg; (d) Maxillula; (e) Second antenna; (f) Valve
图4. Helerocypris incongruens,软体。(a) 清洁足;(b) 第一触角;(c) 行走足;(d)大颚;(e)第二触角;(f)壳体
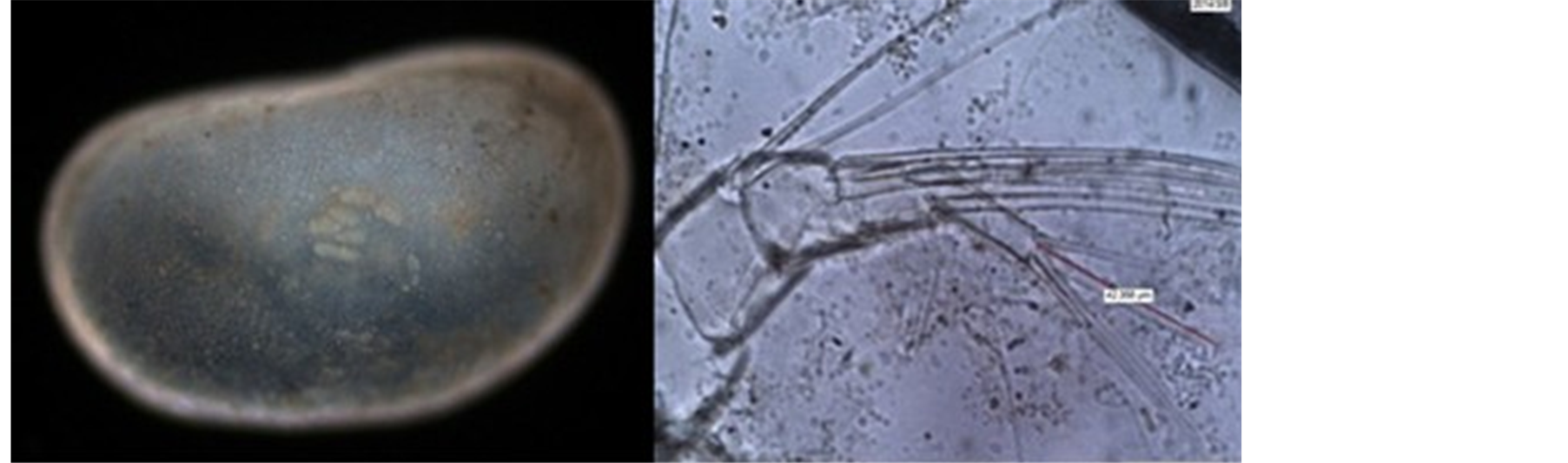
Figure 5. Electron micrograph of Candonocypris novaezelandiae and its appendages, male
图5. Candonocypris novaezelandiae显微镜电子图及其附肢图[15] ,雄性
软体描述:步足基节腹侧近末端刚毛长约与近中部刚毛相当,次末节端部具两根刚毛(该结构为玻星介属独有特征),其中一长一短。清洁足次末节腹侧具有两根刚毛。两尾叉对称;尾叉较直;尾叉附属结构与尾叉连接处具有一个三角形结构,这一特征符合爬星介亚科的主要鉴定依据。非调和异星介Helerocypris incongruens与泽氏玻星介Candonocypris novaezelandiae的尾叉细节比较(见图6)。
讨论:当前种右壳内板上的粗脊状织边十分独特,因此较容易鉴定。此次所获样品尾叉附属结构与尾叉连接处的三角形结构较De Deckker (1981)所绘图形更典型。
比较:这个物种与Fabaeformiscandona fabaeformis (Fischer, 1851)相似,大颚上都有羽状刚毛,但不同后者的是它较大的甲壳,高和长的比例基本相等,右壳比左壳长但较左壳窄。泽氏玻星介H. incongruous尾叉在连接处有明显的三角结构,而豆形豆形玻璃介F. fabaeformis没有,前者终端三个爪子较短,并有一刚毛,刚毛长度为最长终端爪子的一半左右,后者只有两个相对较长的终端爪子,刚毛很短,不易注意到(见图7)。行走足都有四节,每一节上都出现刚毛,但是刚毛的数量和长度有很大的差异。
 (a)
(a) (b)
(b)
Figure 6. Comparison of the tail fork: C. novaezelandiae have a triangular structure obviously at the junction, but H. incongruens don’t have. (a) H. incongruens, the tail fork and appendages; (b) C. novaezelandiae, the tail fork and appendages
图6. 尾叉比较:泽氏玻星介在连接处有明显的三角结构,而非调和异星介没有。(a) 非调和异星介,尾叉和附属物[15] ; (b) 泽氏玻星介,尾叉和附属物[17]
 (a)
(a) (b)
(b)
Figure 7. Comparison of the tail fork: C. novaezelandiae have a triangular structure obviously at the junction, but Fabaeformiscandona fabaeformis don’t have. (a) C. novaezelandiae, the tail fork and appendages; (b) Fabaeformiscandona fabaeformis, the tail fork and appendages
图7. 尾叉比较:泽氏玻星介在连接处有明显的三角结构,而豆形豆形玻璃介没有。(a) 泽氏玻星介,尾叉和附属物[17] ;(b) 豆形豆形玻璃介,尾叉和附属物[15]
长度:前者前三节的刚毛都各自延伸超出了下一节的长度,第三节最明显,下一节的三倍长还多,这是后者所不具有的。数量:前者在第三节末端有两根刚毛,一长一短,差别很大,后者每节末端都只有一根刚毛且短。泽氏玻星介的走足末端比豆科豆科玻璃介的曲度大(见图8)。
分布:新西兰、澳大利亚、日本、我国云南滇池、抚仙湖。
4.4. 鹤庆盆地新属种Parachinocythere reticulate gen.et sp. nov.
描述:壳型小。侧视图亚矩形或长肾形;前端圆;后端狭圆形;背缘直,稍微向后斜;腹缘在中后部向内凹。背视亚球状,两端狭,最大宽度在中间。阀门近等长。前房有两个横沟在,前面的短而浅,后一个深,达到高度的四分之三。表面覆盖网状节点,两个垂直关闭节点在后一个横沟前,挂布边缘附近一个沟微弱的节点在横沟后面。一个小三角形在左瓣的前背缘自由滑动。铰链栉齿型。闭壳肌痕是由四个延长的亚矩形排成直线组成的长卵形瘢痕模式;瘢痕近等长,从底部的第三个疤痕大一点;内收肌瘢痕下面两个近圆形的下颌瘢痕位于前腹部分。边缘区宽度适中。稀疏的唇状孔(见图9)。
备注:目前新品种的特点是它的网状节点纹饰,一个小三角形在左阀前背缘自由滑动。
分布:鹤庆盆地上,云南;中更新世至晚更新世,我国云南滇池、抚仙湖。
比较:目前的新属种与Chinocythere相类似,但是和它的甲壳,壳背后缘,和终端右侧的小三角形齿都是有所差异的。彭金兰鹤庆盆地新属种也和Neochinocythere相类似,但是和它右侧较强壮的端齿和
 (a)
(a) (b)
(b)
Figure 8. Tail fork to compare: (a) C. novaezelandiae, Walking leg; (b) Fabaeformiscandona fabaeformis (Fischer, 1851), Walking leg
图8. 尾叉比较:(a) 泽氏玻星介,走足[17] ;(b) 豆形豆形玻璃介,走足[15]

Figure 9. Electron micrograph of Parachinocythere reticulata gen.et sp. nov
图9. 鹤庆盆地新属种显微镜电子图[17]
左侧光滑的中脊也是不同的。此外,在目前的新属种里,第一个底部内收肌是次伸长状卵形和地步第三和第四内收肌的尺寸近乎是相等的。然而在湖花介科Limnocytheridae中,来自底部的第一个内收肌是小的近圆形的。
5. 结论与不足
5.1. 结论
1) 现代介形虫因为有软体特征,这是介形虫化石难以保存的部分,在显微镜下解剖并观察,鉴定起来比介形虫化石的鉴定容易得多,也更加精确;
2) 在抚仙湖发现了克氏瘤丽星介Physocupria kraelini、非调和异星介Heterocypris incongruens、泽氏玻星介Candonocypris novaezelandiae、鹤庆盆地新属种Parachinocythere reticulata gen. et sp. nov.四种介形虫,其中鹤庆盆地新属种Parachinocythere reticulata gen. et sp. nov.在滇池还未被发现过。这次在抚仙湖是滇中地区属首次报道;
3) 在抚仙湖发现的现生介形虫与前人在滇池发现的在群体组合上有很大的差别[15] ,由此说明了介形虫在不同的水体条件中具有决然不同的群体组合,为今后将介形虫这一替代指标用于古环境古气候的恢复工作中,提高云南地区地球科学的研究水平等工作起铺垫作用。
4) 与滇池现代介形比较起来,我们发现抚仙湖多了一个优势种Parachinocythere reticulata gen. et sp. nov.该种在滇池的柱状样品里有出现,但在表层沉积物及现代样品中并未见到。说明此种介形不适宜生活在湖水污染严重的水体中。
5.2. 不足
其中鹤庆盆地新属种Parachinocythere reticulata gen. et sp. nov.由于只发现壳体,所以没有得到详细的软体资料。
致 谢
致谢:谢曼平–国家基金11(01300206089918036)、云南省高端人才引进项目(2010CI111)、湖泊沉积与环境变化云南省创新团队(2010CI)资助研究成果。作者对所有在此次论文完成过程给予帮助和建议的所有老师和同学谨致谢忱!
基金项目
谢曼平–国家基金11(01300206089918036)。
参考文献 (References)
- [1] 谢曼平, 朱立平, 彭萍, 等 (2008) 8.4ka以来纳木错湖芯介形类组合的环境变化意义. 地理学报, 9, 931-944.
- [2] 李军, 余俊清 (2001) 湖相介形类壳体地球化学在环境变化研究中的应用与进展. 湖泊科学, 4, 367-374.
- [3] Lowe, J.J. and Walker, M.J.C. (1984) Reconstructing quaternary environments. 229-232.
- [4] 张家武, 何晶, 陈硕, 李双 (2009) 第四纪湖相介形类壳体化石在古环境中的应用, 种属组合研究进展与问题. 地球科学进展, 11, 1229-1233.
- [5] 禹娜, 赵泉鸿, E.L. Schornikov, 等 (2005) 太湖现生介形虫. 微体古生物学报, 2, 143-151.
- [6] 杨留法, 黄宝仁 (1983) 云南抚仙湖表层沉积物中的介形类及其分布规律的初步研究. 科学通报, 617-621.
- [7] 刘阳, 吴钢, 高正文 (2008) 云南省抚仙湖和杞麓湖流域土地利用变化对水质的影响. 生态学杂志, 3, 447-453.
- [8] 张洁, 张志 (2008) 基于MODIS数据的云南抚仙湖星云湖水质污染遥感调查方法研究. 水文地质工程地质, 5, 92-96.
- [9] 陆鸿宾 (1981) 抚仙湖的气候特征. 海洋湖沼学通报, 4, 1-2.
- [10] 胡元林, 赵光洲 (2006) 抚仙湖保护与湖区可持续发展. 经济问题探索, 9, 20-25.
- [11] 王建云, 普发贵 (2003) 抚仙湖垂向水质状况及特征研究. 玉溪师范学院学报, 19, 53-58.
- [12] 侯长定, 吴献花 (2002) 抚仙湖一星云湖出流改道工程环境影响分析. 云南地理环境研究, 2, 80-88.
- [13] 张蕊, 苏婧, 霍守亮, 陈艳卿, 纪丹凤, 王月, 吕宁磬 (2012) 抚仙湖营养状态评价及营养物水质标准制定. 环境工程技术学报, 3, 218-222.
- [14] Meisch, C. (2000) Freshwater ostracoda of western and central Europe. Spektrum, 1-522.
- [15] 禹娜, 赵泉鸿, 成鑫荣 (2010) 云南滇池淡水介形类新记录. 微体古生物学报, 4, 344-350.
- [16] 禹娜, 陈立侨, 赵泉鸿 (2007) 太湖介形虫分布与水环境因子间关系的典范对应分析. 微体古生物学报, 1, 53-60.
- [17] 彭金兰 (2002) 云南鹤庆晚第四纪介形类生态特征初探. 微体古生物学报, 2, 144-156.