Dynamical Systems and Control
Vol.
10
No.
01
(
2021
), Article ID:
39636
,
10
pages
10.12677/DSC.2021.101003
模拟实际光照的近日节律模型研究
袁梓馨,孙 文
长江大学信息与数学学院,湖北 荆州

收稿日期:2020年11月30日;录用日期:2020年12月23日;发布日期:2020年12月31日

摘要
为适应外界昼夜周期变化,哺乳动物根据外界环境以及生物体内各种因素变化,通过生物钟的导引来维持其正常运转。已有文献假定每天的光照是固定值,但实际中每天光照是随时间变化的。本文首先建立更加符合实际的光照模型,然后讨论在不同光照强度下腹外侧神经元振子数占总神经元振子数的比值以及神经递质耦合强度的离散程度对生物钟的导引的影响。和在周期光暗循环下相比,在模拟实际光照下的光照强度也对生物钟的导引起决定性作用,腹外侧所占神经元振子比值与神经递质耦合强度的离散程度对生物钟的导引影响也较小,背内侧与腹外侧的分裂现象更少,生物钟导引到外界周期的能力更强。
关键词
近日节律,视交叉上核(SCN),耦合强度的离散
The Circadian Model with the Simulated Actual Illumination
Zixin Yuan, Wen Sun
School of Information and Mathematics, Yangtze University, Jingzhou Hubei

Received: Nov. 30th, 2020; accepted: Dec. 23rd, 2020; published: Dec. 31st, 2020

ABSTRACT
To adapt to the circadian changes of the outside world, mammals regulate the biological clocks to maintain their normal operation according to the changes in the outside environment and various internal biological factors. In the existing literature, the illumination of each day is assumed to be fixed, but it changes over time. This paper firstly establishes a more realistic illumination model, then discusses the influence of the dispersion degree of neurotransmitter coupling strength and the ratio of the number of ventrolateral neurons to the total number of neurons under different light intensity on the entrainment of the SCN. The experimental results show that the illumination intensity plays a decisive role in the entrainment of the SCN, while the ratio of neurons in the ventrolateral region and the dispersion degree of neurotransmitter coupling intensity have little influence on the entrainment of the SCN. Compared with the periodic dark cycle, the illumination intensity also plays a decisive role in the entrainment, and the ratio of the neurons in the ventrolateral area and the dispersion degree of the neurotransmitter coupling intensity also have little influence on it. However, under the simulating actual illumination, there are fewer splits and a stronger ability of the entrainment to the external cycles.
Keywords:Circadian Rhythm, Suprachiasmatic Nucleus (SCN), Dispersion of Coupling Strength

Copyright © 2021 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
由于地球的昼夜交替,各类生物体逐渐建立了相应的生理机能 [1]。这种在外界自然昼夜循环下,生物体为了保持与外界光暗循环相同周期而建立的生物节律,称为近日节律(一般在22小时到28小时之间)。近日节律的研究具有非常重要的生物学意义,在多个领域都有应用。这是复杂系统理论在生物体中的一个具体体现 [2] [3] [4] [5]。哺乳动物的近日节律受到体内位于下丘脑视交叉上核(SCN)的内源性时钟控制 [6] [7]。
SCN由大量神经元组成,分为左右两个核,在结构和功能上可划分为腹外侧(VL)和背内侧(DM) [8]。腹外侧能够直接接受来自视网膜的光信号,背内侧接受来自腹外侧的信号 [9]。这些神经元在没有周期光暗循环情况下能够周期地运动,称为自振荡 [10]。自振荡是SCN能够输出自振荡周期信号保持近日节律的根本原因 [11]。其核心机制是基因转录翻译组成的负反馈环 [12] [13]。SCN的神经元之间存在着耦合,不同的神经元之间自振荡周期不同,实验发现神经元的自振荡周期服从正太分布,在22小时到28小时之间 [14] [15]。神经元之间通过各种神经递质进行耦合,同步后能使整个SCN神经网络达到统一的周期 [16] [17]。若神经元受到影响,SCN未达到统一周期,腹外侧与背内侧周期不同步,称腹外侧与背内侧分裂。
SCN能够根据外界环境以及体内各种因素的变化控制生物的近日节律,维持生物体的正常运转 [18]。SCN与外界光暗循环周期保持同步的作用方式,称为导引 [19]。SCN能够导引的外界光暗循环周期范围称为导引范围。实验 [20] 发现光照强度能够增加SCN对外界光暗循环的导引能力。
由于光照和神经递质对SCN保持近日节律周期有重要影响 [21] [22],Gonze [14] 等人综合考虑光照和神经递质耦合,模拟单个神经元振子的自振荡,建立了描述近日节律的Goodwin模型。通过研究发现SCN的自振荡周期与耦合强度的大小成正比。顾长贵 [23] 在Goodwin模型的基础上,建立周期光暗循环下的近日节律模型,考虑耦合强度(服从正态分)的离散程度以及腹外侧神经元振子数占SCN总神经元振子数的比值对SCN的导引范围的影响,讨论了整个SCN网络的集体行为。发现在周期26小时的外界周期光暗循环下,耦合强度的离散能够显著增加SCN的导引能力,但在外界周期为22小时下,SCN的导引能力几乎不增加。
在顾长贵 [23] 的模型中,光照波形为方波。即在周期光暗循环中,假定有光照时其强度取恒定值,无光照时光照强度为0。但是实际生活中,一天中的光照非恒定值,而是随时间变化的。同时考虑到地理位置和季节的不同,实际光照强度会不同,耦合强度的离散程度以及腹外侧神经元振子数占SCN总神经元振子数的比值对SCN导引都有不同影响,因此有必要对在模拟实际光照条件下SCN的近日节律进行研究。
2. 模型描述
周期光暗循环下的近日节律模型 [21] 如下:
(1)
(2)
(3)
(4)
其中 、 、 、 和 分别是第i个神经元振子的mRNA、蛋白、抑制子、神经递质和标度因子,N为总振子数, 是光照项,n是Hill系数。时钟基因mRNA 产生蛋白 ,然后激活转录抑制子 ,组成单个神经元振子的负反馈环。神经递质 由mRNA 感生,不同神经元振子通过神经递质 的平均场F进行耦合。 (满足正态分布)表示每个神经元振子对平均场F的耦合强度,其平均值 ,标准偏差 代表离散程度。当 时,耦合全同。按照文献 [23] 使标度因子 满足均值为1,标准偏差为0.05的正态分布,可以使每个神经元振子的自振荡周期固定在24小时,能够更好地讨论耦合强度的离散程度 [24] [25] 在模拟实际光照下对SCN导引范围的影响。
由于SCN的背内侧和腹外侧对光照信号的感应程度不同,实验 [26] 发现在同一核内,腹外侧能耦合背内侧,但背内侧对腹外侧的影响非常小。用 代表腹外侧神经元振子数占总神经元振子数的比例,定义标号为1到pN (取整数)的神经元振子属于腹外侧,其余神经元振子属于背内侧。其中腹外侧神经递质平均场 [23] 为
(5)
背内侧神经递质平均场 [23] 为
(6)
因此神经递质的平均场
(7)
相同步序参量可以描述腹外侧和背内侧之间的周期同步程度,其定义如下 [27]:
(8)
代表对时间求平均,其中 和 分别代表从平均场 和 的时间序列中用希尔伯特转换提取出来的相位信息。 时代表腹外侧和背内侧的相位完全不一致, 时代表腹外侧和背内侧的相位完全一致。
周期光暗循环下的近日节律模型中光照波形是方波形式,光照项为 [23]
(9)
考虑到实际光照强度在一天内随时间的变化情况,将光照项改为模拟实际光照,即
(10)
其中t为外界时间, 为外界光暗循环的周期, 为光照强度( ), 代表外界时间t对外界光暗循环的周期 取模,振子总数N = 100,其他模型参数同文献 [25]:
采用四阶Runge-Kutta算法进行计算,步长0.1 h,舍弃前2000步暂态,保留最后的4000步,计算腹外侧和背内侧的平均场时间序列的周期。
3. 结果分析
3.1. 腹外侧振子数占总振子数的比值p对SCN周期的影响
为方便起见,假设耦合强度全同。首先讨论腹外侧振子数占总振子数的比值p对腹外侧和背内侧导引到外界周期的影响。在模拟实际光照下,SCN周期变化情况如图1所示。图1(a)中, ,腹外侧和背内侧周期与外界周期同步程度很高,腹外侧与背内侧周期峰值接近;图1(b)中, ,腹外侧与背内侧的周期都与外界光暗循环的周期一致,腹外侧与背内侧周期峰值相同,且相位同步。因此在相同的光照强度下,增加腹外侧振子数可以增加SCN对外界光暗循环的导引能力,能够帮助SCN与外界光暗循环周期保持一致。在其他条件相同时,在周期光暗循环下的腹外侧振子数占总振子数的比值p对腹外侧和背内侧导引到外界周期的情况如图2所示。从图1(a)与图2(a)可以看出,当腹外侧振子数占总振子数的比值p较小时,模拟实际光照下的腹外侧和背内侧的周期更为同步;从图1(b)与图2(b)可以看出,当腹外侧振子数占总振子数的比值p较大时,模拟实际光照下的腹外侧与背内侧与外界周期保持一致的程度更高。这说明模拟实际光照能够增加SCN对外界的导引能力。因此,在实际光照下腹外侧神经元振子数目对SCN的导引有重要影响。
为了更清楚地了解模拟实际光照下,不同的 值对SCN与外界周期关系的影响,用p-Kf平面内的色相图来描述平均场的周期变化。图3显示在模拟实际光照下,耦合强度全同时,腹外侧和背内侧的周期在p-Kf平面内的变化。图3(a)、图3(b)表示在外界周期22小时时,腹外侧(背内侧)与外界环境达到统一周期(深蓝色)。图3(c)、图3(d)表示在外界周期26小时时,腹外侧(背内侧)与外界环境达到周期一致(红棕色)。在外界周期为22小时和26小时下,在光照强度大于一个阈值后,腹外侧周期与外界周期能够保持同步,不受p值的变化影响;但背内侧若要与外界周期保持一致则同时受Kf和p值的影响;在相同的光照下,背内侧与外界周期保持周期同步所需的p值更大。因此,在模拟实际光照下,外界周期为26小时比周期为22小时的腹外侧与背内侧更易发生分裂。
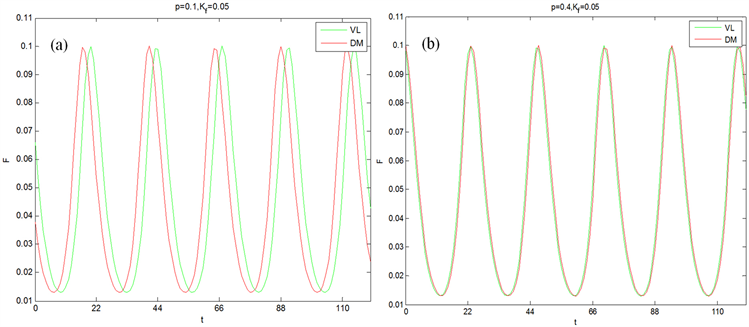
Figure 1. The time series of the mean field of the ventrolateral part (VL) and dorsomedial part (DVLM) under the simulated actual illumination with a period of 22 hours
图1. 在周期为22小时的模拟实际光照条件下,腹外侧(VL)和背内侧(DM)的平均场的时间序列图
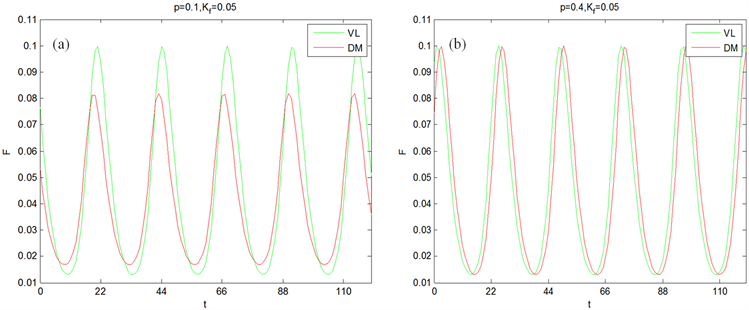
Figure 2. The time series of mean field of the ventrolateral part (VL) and dorsomedial part (DM) under the periodic light-dark cycle with a period of 22 hours [23]
图2. 在周期为22小时的周期光暗循环条件下,腹外侧(VL)和背内侧(DM)的平均场的时间序列图 [23]
在周期光暗循环下,腹外侧和背内侧的周期在p-Kf平面内的变化如图4所示。与图4相比,在模拟实际光照下,代表腹外侧(背内侧)与外界周期一致的深蓝色(红棕色)区域面积均有增加,这表明模拟实际光照能帮助SCN导引到外界周期;在模拟实际光照下比在周期光暗循环下,光照强度的阈值和p值更小,这说明在模拟实际光照下腹外侧能够更快与外界周期达到同步。
3.2. 耦合强度的离散对SCN周期的影响
考虑耦合强度的离散程度对SCN周期的影响,用色相图描述在模拟实际光照下SCN的周期变化情况。图5显示在周期为22小时的模拟际光照下,耦合强度离散时的腹外侧和背内侧周期在p-Kf平面内的变化。在周期为22小时的周期光暗循环下,耦合强度离散时的SCN的周期在p-Kf平面内的变化如图6所示。

Figure 3. The ventrolateral part (VL) and dorsomedial part (DM) periodic hue diagrams in the p-Kf plane under the simulated actual illumination
图3. 模拟实际光照下,腹外侧(VL)和背内侧(DM)周期在p-Kf平面内的色相图
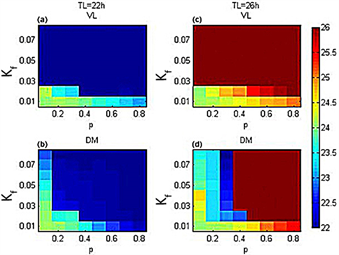
Figure 4. Periodic hue diagrams of ventrolateral part (VL) and dorsomedial part (DM) in the p-Kf plane under the periodic light-dark cycle [23]
图4. 周期光暗循环下,腹外侧(VL)和背内侧(DM)周期在p-Kf平面内的色相图 [23]
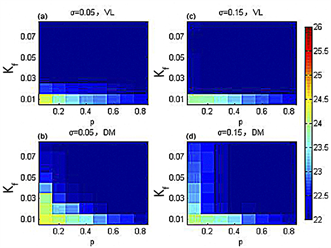
Figure 5. Under the simulated actual illumination with a period of 26 hours, the periodic hue diagrams of the ventrolateral part (VL) and dorsomedial part (DM) in the p-Kf plane
图5. 周期22小时模拟实际光照下,腹外侧(VL)和背内侧(DM)周期在p-Kf平面内的色相图
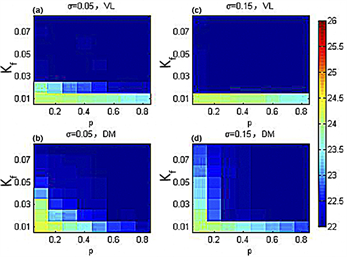
Figure 6. Under the periodic light-dark cycle with a period of 22 hours, the ventrolateral part (VL) and dorsomedial part (DM) periodic hue diagram in the p-Kf plane [23]
图6. 周期22小时周期光暗循环下,腹外侧(VL)和背内侧(DM)周期在p-Kf平面内的色相图 [23]
图5(a)、图5(b)表示在标准偏差 时,腹外侧(背内侧)与外界周期达到周期一致(深蓝色)。图5(c)~(d)表明在标准偏差 时,腹外侧(背内侧)与外界周期达到统一周期(深蓝色)。可以看出,在外界周期为22小时的模拟实际光照下,随着耦合强度离散程度的增加,代表腹外侧(背内侧)与外界周期达到同步的深蓝色区域面积显著增加,说明耦合强度离散程度的增加能帮助SCN更快地导引到外界周期。与图6相比,在外界周期为22小时的模拟实际光照下,代表腹外侧(背内侧)与外界周期达到一致的深蓝色区域面积均略有增加,说明模拟实际光照能够增加SCN导引到外界周期的能力。
在周期为26小时的模拟实际光照下,图7显示在耦合强度离散时的SCN周期在p-Kf平面内的变化。图7(a)、图7(b)表示在标准偏差 时,腹外侧(背内侧)与外界周期达到同步(红棕色)。图7(c)、图7(d)表明在标准偏差 时,腹外侧(背内侧)与外界周期达到统一(红棕色)。可以看出,在外界周期为26小时的模拟实际光照下,随着耦合强度离散程度的增加,代表腹外侧与外界周期一致的红棕色区域面积无明显变化,代表背内侧与外界周期同步的红棕色区域面积显著增加。这说明在周期为26小时的模拟实际光照下,增加耦合强度的离散能够显著增加SCN导引到外界周期的能力。
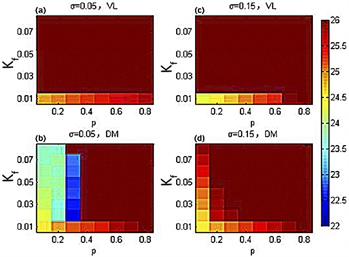
Figure 7. Under the simulated actual illumination with a period of 26 hours, the ventrolateral part (VL) and dorsomedial part (DM) periodic hue diagrams in the p-Kf plane
图7. 在周期为26小时的模拟实际光照下,腹外侧(VL)和背内侧(DM)的周期在p-Kf平面内的色相图
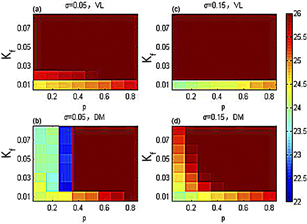
Figure 8. Underthe periodic light-dark cycle with a period of 26 hours, the periodic hue diagrams of the ventrolateral part (VL) and dorsomedial part (DM) in the p-Kf plane [23]
图8. 周期为26小时的周期光暗循环下,腹外侧(VL)和背内侧(DM)的周期在p-Kf平面内的色相图 [23]
在周期为26小时的模拟实际光照下,图8显示在耦合强度离散时的SCN周期在p-Kf平面内的变化。与图8对比,在模拟实际光照下,代表腹外侧(背内侧)与外界周期一致的红棕色区域面积略增加,说明模拟实际光照能增加SCN导引到外界周期的能力。
3.3. 相同步序参量
相同步序参量能够衡量网络的同步程度。我们进一步在p-Kf平面内讨论在实际光照下耦合强度的离散程度对相同步序参量的影响。
图9显示在模拟实际光照下相同步序参量在p-Kf平面内的变化。可以看出,在外界周期为22小时下,耦合强度的离散程度的增大使序参量 值略有减小,说明耦合强度离散程度的增大能增加腹外侧与背内侧的分裂。在外界周期为26小时的模拟实际光照下,耦合强度的离散程度的增大使序参量R值显著增加,即SCN与外界环境保持周期同步的能力增强。
在周期光暗循环下,相同步序参量在p-Kf平面内的变化如图10所示,腹外侧与背内侧在22小时的周期光暗循环下更易分裂。可以看出,在模拟实际光照下,序参量R值在外界周期为22小时下整体略微增大,腹外侧与背内侧的分裂减少;在外界周期为26小时下,序参量R值整体显著增大,SCN与外界环境更易保持周期同步。即在模拟实际光照的近日节律模型中相同步程度更高,这也说明我们改进的近日节律模型精度更高。
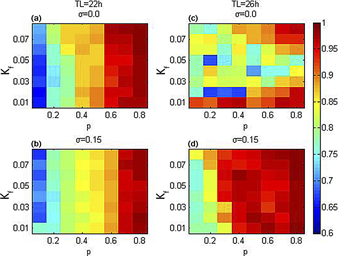
Figure 9. The variation diagram of phase synchronization order parameters in the p-Kf plane under the simulated actual illumination
图9. 模拟实际光照下,相同步序参量在p-Kf平面内的变化图
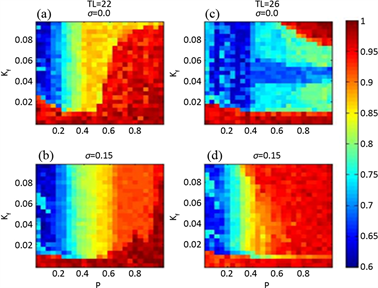
Figure 10. The variation diagram of phase synchronization order parameters in the p-Kf plane under the periodic light-dark cycle [23]
图10. 周期光暗循环下,相同步序参量在p-Kf平面内的变化图 [23]
4. 结论
本文首先提出更加符合实际的近日节律模型,然后讨论在不同光照强度下腹外侧神经元振子数占总神经元振子数的比值以及神经递质耦合强度的离散程度对生物钟的导引的影响。数值图像结果表明本文构建的近日节律模型比已有模型精度更高。说的更具体一些,和在周期光暗循环下一样,在模拟实际光照下的光照强度同样对SCN的导引起决定性作用,腹外侧所占神经元振子比值与神经递质耦合强度的离散程度对SCN的导引影响也较小;但在模拟实际光照下的背内侧与腹外侧的分裂现象更少,SCN导引到外界周期的能力更强。
文章引用
袁梓馨,孙 文. 模拟实际光照的近日节律模型研究
The Circadian Model with the Simulated Actual Illumination[J]. 动力系统与控制, 2021, 10(01): 24-33. https://doi.org/10.12677/DSC.2021.101003
参考文献
- 1. Gonze, D., Bernard, S., Waltermann, C., Achim, K. and Hanspeter, H. (2005) Spontaneous Synchronization of Coupled Circadian Oscillators. Biophysical Journal, 89, 120-129.
https://doi.org/10.1529/biophysj.104.058388 - 2. Welsh, D.K., Takahashi, J.S. and Kay, S.A. (2010) Suprachiasmatic Nucleus: Cell Autonomy and Network Properties. Annual Review of Physiology, 72, 551-577.
https://doi.org/10.1146/annurev-physiol-021909-135919 - 3. Vasalou, C., Herzog, E.D. and Henson, M.A. (2009) Small-World Network Models of Intercellular Coupling Predict Enhanced Synchronization in the Suprachiasmatic Nucleus. Journal of Biological Rhythms, 24, 243-254.
https://doi.org/10.1177/0748730409333220 - 4. 孙文, 陈忠. 基于一个控制器的非线性耦合复杂网络牵制控制同步研究(英文) [J]. 长江大学学报(自然科学版)理工卷, 2010, 7(3): 9-13.
- 5. Scheer, F.A.J.L., Wright, K.P., Kronauer, R.E. and Czeisler, C.A. (2007) Plasticity of the Intrinsic Period of the Human Circadian Timing System. PLoS ONE, 2, e721.
https://doi.org/10.1371/journal.pone.0000721 - 6. Bernard, S., Gonze, D., Cajavec, B., Herzel, H. and Kramer, A. (2007) Synchronization Induced Rhythmicity of Circadian Oscillators in the Suprachiasmatic Nucleus. PLoS Computational Biology, 3, 667-676.
https://doi.org/10.1371/journal.pcbi.0030068 - 7. Smith, M.R., Burgess, H.J., Fogg, L.F., Eastman, C.I. and Shin, Y. (2009) Racial Differences in the Human Endogenous Circadian Period. PLoS ONE, 4, e6014.
https://doi.org/10.1371/journal.pone.0006014 - 8. 孙芳玲, 郭德玉, 王文. 光照对大脑功能的影响研究进展[J]. 照明工程学报, 2017, 28(6): 16-19.
- 9. 邢陈, 宋伦. 昼夜节律产生和维持的调控系统[J]. 军事医学, 2017, 41(8): 698-702.
- 10. 顾长贵. 不同光照条件下的哺乳动物近日节律模型[D]: [博士学位论文]. 上海: 华东师范大学, 2011.
- 11. Ute, A., Granada, A.E., Westermark, P.O., Markus, H., Achim, K. and Hanspeter, H. (2014) Coupling Governs Entrainment Range of Circadian Clocks. Molecular Systems Biology, 6, 438.
https://doi.org/10.1038/msb.2010.92 - 12. Locke, J.C., Westermark, P.O., Kramer, A. and Herzel, H. (2008) Global Parameter Search Reveals Design Principles of the Mammalian Circadian Clock. BMC Systems Biology, 2, 22.
https://doi.org/10.1186/1752-0509-2-22 - 13. Noguchi, T., Watanabe, K., Ogura, A. and Yamaoka, S. (2004) The Clock in the Dorsal Suprachiasmatic Nucleus Runs Faster than That in the Ventral. The European Journal of Neuroscience, 20, 3199-3202.
https://doi.org/10.1111/j.1460-9568.2004.03784.x - 14. 秦宁, 闵清, 胡文祥. 生物钟的调控[J]. 交叉科学快报, 2018, 2(4): 93-102.
- 15. 贾兴, 孙海义. 复杂网络同步控制方法研究综述[J]. 动力系统与控制, 2018, 7(4): 318-327.
- 16. 郭向东, 陈芳跃. 特征多项式理论在布尔控制网络中的应用[J]. 动力系统与控制, 2018, 7(3): 214-225.
- 17. 李嘉敏, 宾红华, 黄振坤. 具有不匹配参数的脉冲离散网络准同步[J]. 动力系统与控制, 2017, 6(4): 158-163.
- 18. 谭亚华, 谭建国. 混合时滞随机Hopfield神经网络的均方渐近稳定性[J]. 动力系统与控制, 2019, 8(4): 263-270.
- 19. 秦曦明, 郭金虎. 哺乳动物生物钟同步化的研究进展[J]. 科学通报, 2017, 62(25): 2849-2856.
- 20. 肖义军, 钟磊发. 哺乳动物昼夜节律的调控及其分子机制[J]. 生物学通报, 2018, 53(5): 2-4.
- 21. Welsh, D.K. (2009) Limits of Single-Cell Autonomy in the Suprachiasmatic Nucleus. Sleep & Biological Rhythms, 7, 252-259.
https://doi.org/10.1111/j.1479-8425.2009.00425.x - 22. Baba, K., Ribelayga, C.P., Iuvone, P.M. and Gianluca, T. (2018) The Retinal Circadian Clock and Photoreceptor Viability. Advances in Experimental Medicine and Biology, 1074, 345-350.
https://doi.org/10.1007/978-3-319-75402-4_42 - 23. Gu, C., Wang, J. and Liu, Z. (2009) Free-Running Period of Neurons in the Suprachiasmatic Nucleus: Its Dependence on the Distribution of Neuronal Coupling Strengths. Physical Review E, 80, Article ID: 030904.
https://doi.org/10.1103/PhysRevE.80.030904 - 24. Reppert, S.M. and Weaver, D.R. (2001) Molecular Analysis of Mammalian Circadian Rhythms. Annual Review of Physiology, 63, 647.
https://doi.org/10.1146/annurev.physiol.63.1.647 - 25. Huang, R.C. (2018) The Discoveries of Molecular Mechanisms for the Circadian Rhythm: The 2017 Nobel Prize in Physiology or Medicine. Biomedical Journal, 41, 5-8.
https://doi.org/10.1016/j.bj.2018.02.003 - 26. Liang, X., Tang, M., Dhamala, M. and Liu, Z. (2009) Phase Synchronization of Inhibitory Bursting Neurons Induced by Distributed Time Delays in Chemical Coupling. Physical Review E, 80, Article ID: 066202.
https://doi.org/10.1103/PhysRevE.80.066202 - 27. 岳敏, 杨禹, 郭改丽, 秦曦明. 哺乳动物生物钟的遗传和表观遗传研究进展[J]. 遗传, 2017, 39(12): 1122-1137.