Botanical Research
Vol.07 No.03(2018), Article ID:25011,7
pages
10.12677/BR.2018.73037
Construction and Transformation of Recombinant Plasmid of Gene SSCD1 Promoter and GUS Gene in Arabidopsis thaliana
Peilin Liu1, Tiantian Zhi1, Chunmei Ren1,2*
1College of Bioscience and Biotechnology, Hunan Agricultural University, Changsha Hunan
2Crop Gene Engineering Key Laboratory of Hunan Province, Changsha Hunan
Received: Apr. 22nd, 2018; accepted: May 16th, 2018; published: May 23rd, 2018

ABSTRACT
The tyrosine degradation pathway is an important metabolic pathway in animals, but in plants, less is known about this pathway. Our previous study found that: in Arabidopsis thaliana, the SSCD1 gene encodes the final enzyme of the tyrosine degradation pathway, fumarate lace to acetate hydrolase (FAH), and mutation of this gene results in cell death in Arabidopsis under short day. To investigate the tissue expression characteristics of the SSCD1 gene, a fusion expression vector of Arabidopsis thaliana SSCD1 gene promoter and GUS gene was constructed and transformed into Arabidopsis thaliana, and the transgenic plant was obtained and identified by GUS histochemical staining. The GUS histochemical staining showed that the GUS gene driven by the SSCD1 gene promoter was strongly expressed in the hypocotyls, but weakly in the cotyledons and roots. However, the expression was almost absent in the true leaves. The results of this study indicate that the SSCD1 gene plays a more important role in hypocotyls, which is very important for deeply exploring tyrosine degradation pathway in plants.
Keywords:Arabidopsis, SSCD1 Mutant, Tyrosine Degradation Pathway, GUS
拟南芥SSCD1基因启动子与GUS重组载体的构建与转化
刘佩琳1,支添添1,任春梅1,2*
1湖南农业大学生物科学技术学院,湖南 长沙
2作物基因工程湖南省重点实验室,湖南 长沙

收稿日期:2018年4月22日;录用日期:2018年5月16日;发布日期:2018年5月23日

摘 要
酪氨酸降解途径是动物中一条重要的代谢途径,而在植物中对该途径了解较少。我们前期研究发现:在模式植物拟南芥中,SSCD1基因编码酪氨酸降解途径最后一个酶——延胡索酰乙酰乙酸酶(fumaryl acetoacetate hydrolase),该基因突变后导致拟南芥在短日照下出现细胞死亡。为了探讨SSCD1基因的组织表达特征,本研究构建了拟南芥SSCD1基因启动子与GUS的融合表达载体并转入拟南芥,得到其转基因植株,经GUS组织化学染色,发现SSCD1基因启动子驱动的GUS基因在下胚轴中表达较强,子叶和根中表达较弱,而在真叶中几乎没有表达。该研究结果表明SSCD1基因在下胚轴中发挥着更重要的作用,对深入探讨植物中酪氨酸降解途径具有重要的作用。
关键词 :拟南芥,SSCD1突变体,酪氨酸降解途径,GUS

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
酪氨酸降解途径在动物体内发挥着重要作用,在人类和动物中若被阻断将会造成很严重的损伤,甚至死亡 [1] 。延胡索酰乙酰乙酸酶(FAH)参与酪氨酸降解的最终途径,其主要作用是将延胡索酰乙酰乙酸水解成乙酰乙酸和延胡索酸进入三羧酸循环彻底分解 [2] 。在动物中,FAH合成中断会导致动物患上酪氨酸血症并引起死亡 [3] 。而在植物体内,编码FAH酶的SSCD1基因突变会导致拟南芥在短日照条件下发生细胞死亡,而长日照下表型正常 [4] ,因此利用基因工程技术来研究SSCD1基因的表达模式有助于了解酪氨酸降解途径在植物体内的作用机制。
启动子处于基因转录起始位点的上游,其作为重要顺式元件参与了一系列基因的表达和调控,并能被RNA聚合酶识别与结合 [5] 。将启动子与GUS基因融合,通过转基因实验验证基因的表达活性和组织特异性表达是研究基因表达模式的一种常用方法 [6] [7] 。
本试验以拟南芥为研究材料,通过构建SSCD1基因启动子以及GUS的融合表达载体,随后将其导入拟南芥植株内,通过抗性筛选得到稳定可遗传的转基因植株,进一步采用GUS组织化学染色,检测SSCD1基因的启动子是否成功转入和正常激活GUS基因。本实验的完成为后续该基因的研究提供一定借鉴与帮助。
2. 试验材料与方法
2.1. 试验材料
2.1.1. 植物材料及培养条件
拟南芥(Arabidopsis thaliana)野生型Col-0由湖南农业大学作物基因工程湖南省重点实验室植物信号传导课题组提供。
使用消毒液(20% bleach + 0.1% Triton100)将拟南芥种子浸泡10 min,用无菌水于超净工作台上清洗3~4次,适当密度接种在MS固体培养基上。4℃黑暗春化3 d后,转入人工气候箱生长,培养温度为(22 ± 2)℃,光周期为长日照(16 h光照/8 h黑暗) [8] 。
2.1.2. 表达载体
本试验采用的载体为pCAMBIA1301,该载体的启动子为35S启动子,标记基因为GUS 基因,抗性基因为卡那霉素抗性基因,植物筛选标记基因为潮霉素抗性基因。
2.1.3. 菌株
本试验所用到大肠杆菌E. coli DH5α和根癌农杆菌GV3101均由湖南农业大学作物基因工程湖南省重点实验室植物信号传导课题组提供。
2.1.4. 试剂
DNA提取试剂盒购自北京天根生化科技有限公司,PCR所用的反应体系以及卡那霉素和氨苄青霉素均购自于北京TaKaRa公司,质粒提取试剂盒和切胶回收试剂盒购自于AXYGEN公司,酶切连接用到的试剂以及PstI酶和NcoI酶均购自北京宝生物工程有限公司,其他试剂购自于国药集团化学试剂有限公司。
2.1.5. 培养基
LB培养基配方为胰蛋白胨10 g/L,酵母粉5 g/L,NaCl 10 g/L,琼脂粉15 g/L (固体培养基需要加),调pH = 7.0;YEB培养基配方为胰蛋白胨10 g/L,酵母粉10 g/L,NaCl 5 g/L,琼脂粉15 g/L (固体培养基需要加),调pH = 7.0。
2.2. 试验方法
2.2.1. 拟南芥总DNA的提取
根据DNA提取试剂盒中的说明书进行提取。
2.2.2. PCR引物的设计及SSCD1基因启动子的扩增
通过TAIR网站获取拟南芥SSCD1的基因启动子序列,经软件plantcare分析后选取ATG前2000 bp序列(启动子区域),利用Primer Premier 5软件设计其特异性引物SSCD1-F/SSCD1-R,在5’端分别加上PstI和NcoI酶切位点,引物序列为:
SSCD1-F,5’-AACTGCAGCTCTCCAACGACAACT-3’(下划线处为酶切位点PstI)
SSCD1-R,5’- CATGCCATGGTGACGATACAGATAC-3’(下划线处为酶切位点NcoI)
使用高保真酶对试剂盒提取的拟南芥野生型Col-0基因组DNA进行PCR扩增。退火温度根据引物合成时的预测温度生成(退火温度为60℃)。将扩增得到的PCR产物经琼脂糖(1.0%)凝胶电泳检测,然后回收目的条带并纯化。
2.2.3. SSCD1启动子基因与GUS融合载体重组
将pCAMBIA1301质粒载体(参照质粒提取试剂盒说明书获得载体)和扩增得到的SSCD1启动子片段用PstI和NcoI限制性内切酶在37℃下双酶切过夜,得到的线性载体与启动子片段分别回收后在T4连接酶作用下连接至预期位点,载体构建如图1。体系为10 × T4 Buffer 2 μL,线性化载体6 μL,插入片段扩增产物1 μL,T4 ligase 0.5 μL,ddH2O 10.5 μL。将配置好的体系16℃过夜。从−80℃冰箱里取出大肠杆菌感受态细胞,置于冰上融化,加入20 μL连接产物混匀,在冰上放置30 min;42℃热击90 s,然后迅速转至冰浴,放置1~2 min,加入700 μL LB液体培养基,37℃摇床下振荡培养45 min,6000 g室温下离

Figure 1. Construction of vector pSSCD1::pC1301
图1. pSSCD1::pC1301载体构建
心2 min,去部分上清(剩100 μL),然后混匀浓缩菌液,取50 μL菌液均匀涂布于含50 mg/L Kan的LB固体培养基上,于37℃恒温培养箱倒置,培养过夜 [9] [10] 。挑取阳性克隆并进行测序,将测序后验证正确的重组质粒通过电击转化至根癌农杆菌GV3101中 [11] [12] ,选取阳性克隆进行菌落PCR鉴定。
2.2.4. 重组质粒转化农杆菌与筛选
挑选单菌落溶于10 µL无菌水中,取1 µL进行菌落PCR,检测阳性克隆后,接种于含Kan (50 mg/L)、GEN (50 mg/L)、Rif (25 mg/L)的YEB液体培养基,黑暗条件下,28℃培养至菌液OD600值为0.5~0.6时取出,6000 g离心2 min后弃上清,加入适量悬浮液(5%蔗糖 + 1/2 MS + 0.02% Silwet-L77 + 0.01%6-BA),控制悬浮液中农杆菌浓度(OD600)于0.6~1.0,浸染拟南芥幼嫩花蕾(花序浸染法) [13] 。收取T0代转基因种子,春化后,铺种于添加25 mg/L的Hyg抗性的MS培养基上,筛选出阳性转基因植株。
2.2.5. GUS组织化学检测
将转基因抗性植株浸到GUS染液中,置于37℃保温箱中避光过夜。用50%,70%,100%的浓度梯度乙醇依次漂洗样品,每次浸泡5 min,加入100%乙醇浸泡直至完全脱色,最后在体视显微镜下拍照记录。GUS染色液配制:将X-gluc溶于磷酸钠缓冲液中(含100 mmol/L pH 7.0的磷酸钠缓冲液,10 mmol/L EDTA,5 mmol/L铁氰化钾,5 mmol/L亚铁氰化钾),终浓度为1 mmol/L。
3. 结果与分析
3.1. SSCD1基因启动子的克隆
将扩增产物用琼脂糖凝胶电泳检测,电泳结果显示扩增产物符合预期大小(PCR产物预期大小为2000 bp),且没有非特异性扩增条带(图2),克隆所得目的片段可用于下一步实验。
3.2. pSSCD1::GUS重组载体的构建
对目的片段和pCAMBIA1301载体双酶切,回收的目的片段和目的载体,并进行连接转化。随机挑选阳性单菌落进行菌落PCR检测(图3),挑选阳性单菌落进行扩大培养,提取质粒进行双酶切检测。结果发现(图4)选取与预期结果相符的目的片段(泳道1、2、3、5)进行测序,经比对分析发现目的基因与GUS之间的结合位点与预期一致,表明pSSCD1::GUS融合表达载体成功构建。(4号泳道由于菌污染导致出现三条带,所以舍弃。)
3.3. pSSCD1::GUS重组载体的遗传转化
将pSSCD1::GUS重组表达载体转入根癌农杆菌GV3101中,并进行PCR检测,结果如图5所示,在转入质粒的农杆菌样本中成功检测到大小为2000 bp左右与目的片段大小相似的条带,证明重组质粒成功转入到农杆菌中。挑选转入成功的农杆菌进行扩大培养,通过浸花法转入拟南芥Col-0野生型植株中,进行融合基因的稳定表达实验。潮霉素对融合表达载体拟南芥幼苗生长无抑制作用,将浸染的当代
 注:M为Marker (2 K),Line 1-4为SSCD1基因启动子片段
注:M为Marker (2 K),Line 1-4为SSCD1基因启动子片段
Figure 2. PCR result of SSCD1 promoter
图2. SSCD1基因启动子PCR扩增结果
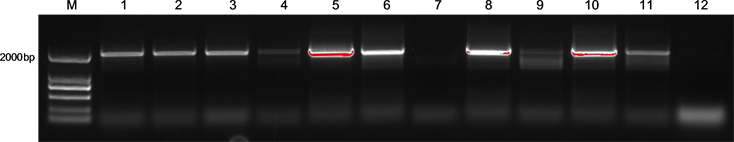 注:M为Marker (2 K),Line 1-12为菌落PCR
注:M为Marker (2 K),Line 1-12为菌落PCR
Figure 3. PCR result of recombinant colonies
图3. 菌落PCR检测
 M为Marker (10 K),Line 1-5为重组载体双酶切片段
M为Marker (10 K),Line 1-5为重组载体双酶切片段
Figure 4. SSCD1::GUS recombinant vector PstI and NcoI double digestion
图4. SSCD1::GUS重组载体PstI与NcoI双酶切
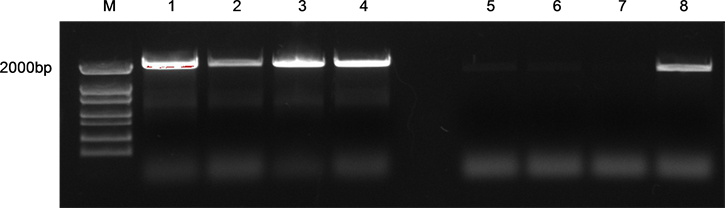 注:M为Marker (2K),Line 1-8为农杆菌阳性克隆
注:M为Marker (2K),Line 1-8为农杆菌阳性克隆
Figure 5. PCR of Agrobacterium colony
图5. 农杆菌菌落PCR
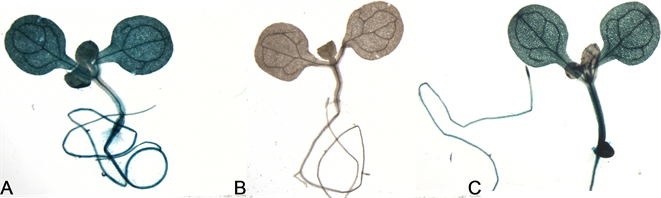 注:A.阳性对照植株;B.阴性对照植株;C.转基因植株
注:A.阳性对照植株;B.阴性对照植株;C.转基因植株
Figure 6. Stain identification of GUS
图6. GUS染色检测结果
种子铺种在含有潮霉素(25 mg/L)的MS培养基上低温春化,在长日照下培养14 d发现具有融合表达载体的幼苗能正常生长即获得转基因植物,转化失败的拟南芥幼苗无法正常生长;而未进行浸染的拟南芥Col-0幼苗在潮霉素培养基上同样无法正常生长。这说明能正常生长的幼苗基因组中成功转入了融合表达载体。
3.4. GUS报告基因定性检测
将转基因植株种子铺种在含潮霉素的MS培养基上,长日照(16 h光照/8 h黑暗)条件下生长7 d。挑选抗性幼苗进行GUS组织化学染色,利用体式显微镜观察染色结果。结果表明:转入含35 S启动子pCAMBIA1301质粒的阳性植株(图6A)中GUS基因在各个部位都高度表达,而未在阴性对照植株中表达(图6B),说明GUS染色方法可行。在同样方法染色的转入SSCD1基因启动子与GUS融合表达载体的转基因植株(图6C)中,GUS基因在下胚轴中表达较强,子叶和根中有表达,但表达比阳性对照弱,而在真叶中几乎没有表达。
4. 结论与讨论
随着基因工程技术飞速发展,基因克隆技术在遗传育种和形状改良等方面体现了十分重要的价值 [14] ,而启动子则是基因表达载体构建中必不可少的原件 [15] 。基因表达的特异性主要取决于该基因的特异性启动子,而启动子本身不具有基因特异性,可启动下游任何基因表达,因此启动子表达的部位可以推测该基因的表达部位。目前转基因工程的主要任务是寻找阻止基因沉默的有效途径和发展可诱导的启动子 [16] 。
本研究成功构建了拟南芥SSCD1基因启动子与GUS的表达载体,通过凝胶电泳图像与测序结果,证明该重组载体即为目的载体。将该重组载体顺利转入拟南芥中并对转化后代用潮霉素进行筛选,获得具有抗性的植株,进一步通过GUS染色法检测,结果表明,SSCD1基因启动子驱动的GUS基因在下胚轴中表达较强,子叶和根中表达较弱,而在真叶中几乎没有表达。该研究结果表明SSCD1基因在拟南芥的不同组织部位有着其特异性的表达,该基因可能在下胚轴中发挥着更重要的作用,但其特异性表达和在下胚轴中的功能有待进一步研究。
文章引用
刘佩琳,支添添,任春梅. 拟南芥SSCD1基因启动子与GUS重组载体的构建与转化
Construction and Transformation of Recombinant Plasmid of Gene SSCD1 Promoter and GUS Gene in Arabidopsis thaliana[J]. 植物学研究, 2018, 07(03): 287-293. https://doi.org/10.12677/BR.2018.73037
参考文献
- 1. Colditz, P.B., Yu, J.S. and Billson, F.A. (1984) Tyrosinaemia II. The Medical Journal of Australia, 141, 244-245.
- 2. Dixon, D.P. and Edwards, R. (2006) Enzymes of Tyrosine Catabolism in Arabidopsis thaliana. Plant Science: An International Journal of Experimental Plant Biology, 171, 360-366.
https://doi.org/10.1016/j.plantsci.2006.04.008 - 3. Lindblad, B., Lindstedt, S. and Steen, G. (1977) On the Enzymic Defects in Hereditary Tyrosinemia. Proceedings of the National Academy of Sciences of the United States of America, 74, 4641-4645.
https://doi.org/10.1073/pnas.74.10.4641 - 4. Han, C., Ren, C., Zhi, T., Zhou, Z., Liu, Y., Chen, F., Peng, W. and Xie, D. (2013) Disruption of Fumarylacetoacetate Hydrolase Causes Spontaneous Cell Death under Short-Day Condition in Arabidopsis. Plant Physiology, 162, 1956-1964.
https://doi.org/10.1104/pp.113.216804 - 5. 陈柳. 农杆菌介导的高赖氨酸基因在油菜中的转化[D]: [硕士学位论文]. 扬州: 扬州大学, 2008.
- 6. 陈庆, 叶放, 宁永强, 等. 拟南芥AtGLR1.3和AtGLR3.3启动子的GUS基因融合表达[J]. 宁波大学学报(理工版), 2010, 23(3): 1-5.
- 7. 王文芳, 刘爱玲, 李琦, 等. 水稻CesA4基因启动子与GUS基因融合表达[J]. 现代生物医学进展, 2009, 9(8): 1459-1461.
- 8. 袁利兵, 张琨, 彭志红, 等. 拟南芥psc1突变体降低蔗糖诱导的花青素积累[J]. 湖南农业大学学报(自科版), 2011, 37(2): 139-142.
- 9. Froger, A. and Hall, J.E. (2007) Transformation of Plasmid DNA into E. coli Using the Heat Shock Method. Journal of Visualized Experiments Jove, 6, 253.
https://doi.org/10.3791/253 - 10. 李春苑. 拟南芥抗寒基因CBF1的克隆、表达载体的构建及鸡冠花离体再生体系的建立[D]: [硕士学位论文]. 兰州: 兰州大学, 2009.
- 11. 邹智, 吴刚, 卢长明. 影响农杆菌转化效率的植物因子H2A1基因的克隆、序列分析及其植物表达载体的构建[J]. 生物技术通报, 2007(6): 83-87.
- 12. 马艺沔. 利用转基因拟南芥研究质外体钙调素在植物生长发育中的作用[D]: [硕士学位论文]. 石家庄: 河北师范大学, 2003.
- 13. Chua, N.H., Niu, Q.W., Henriques, R., et al. (2006) Agrobacterium-Mediated Transformation of Arabidopsis thaliana Using the Floral Dip Method. Nature Protocols, 1, 641.
https://doi.org/10.1038/nprot.2006.97 - 14. 陈常庆. 基因工程技术的现状与发展[J]. 生命科学, 1992(2): 4-7.
- 15. 王莹, 韩烈保, 曾会明. 高等植物启动子克隆方法的研究进展[J]. 生物技术通报, 2007(3): 97-100.
- 16. 贾建航, 王斌. 植物抗病基因克隆研究进展[J]. 中国生物工程杂志, 2000, 20(1): 21-26.