设为首页
加入收藏
期刊导航
网站地图
首页
期刊
数学与物理
地球与环境
信息通讯
经济与管理
生命科学
工程技术
医药卫生
人文社科
化学与材料
会议
合作
新闻
我们
招聘
千人智库
我要投搞
办刊
期刊菜单
●领域
●编委
●投稿须知
●最新文章
●检索
●投稿
文章导航
●Abstract
●Full-Text PDF
●Full-Text HTML
●Full-Text ePUB
●Linked References
●How to Cite this Article
Hans Journal of Agricultural Sciences
农业科学
, 2012, 2, 49-57
http://dx.doi.org/10.12677/hjas.2012.23009
Published Online September 2012 (http://www.hanspub.org/journal/hjas.html)
Pre-Study on the Molecular Mechanism of
Self-Compatibility in Longtanzhenzhuli
Yanping Ye
1*
, Shaoxi Huang
2
, Liping Meng
1
1
College of Agriculture, Guangxi University, Nanning
2
College of Agricultural Sciences in Guangxi, Nanning
Email:
*
yeyanping0705@tom.com
Received: May 25
th
, 2012; revised: Jul. 18
th
, 2012; accepted: Aug. 6
th
, 2012
Abstract:
Longtanzhenzhuli which is a new variety of plum with high quality were used as the experimental
materials in this research. A specific primer was designed based on the conserved sequences of plum (
Prunus
salicina
Lindl.)
S-Rn ase
(
S
) genes, two specific fragments were isolated from Longtanzhenzhuli genome DNA
with tese
specific primers. The sequencing results showed that these fragmen
ts were confirmed as the plum
S
genes. One was first cloned from
Prunus salicina
and named
PsS-RNase
-28, with length of 727 bp,
containing 367 bp intron and 360 bp Exton. Comparing with
Prunus spinosa
S-RNase S
1 gene (EF636467.1),
it showed high identity (98%), more than 79% with other plums
S-RNase
genes. The other fragment with 718
bp in length, containing 343 bp intron and 375 bp Exton, showed high identity (99%) with
Prunus salicina
PsS-RNase-h
gene (AB084148.1), and more than 85% with other plums
S-RNase
genes. And sequences
follow-up analysis of these fragments would be benefited to study on the mechanism of self-compatibility of
Longtanzhenzhuli.
Keywords:
Longtanzhenzhuli; Self-Incompatibility; Self-Compatibility;
S-Rnase Genes
优良品种龙滩珍珠李自花结实分子机理初探
叶燕萍
1*
,黄绍西
2
,孟丽萍
1
1
广西大学农学院,南宁
2
广西农业科学院园艺研究所,南宁
Email:
*
yeyanping0705@tom.com
收稿日期:
2012
年
5
月
25
日;修回日期:
2012
年
7
月
18
日;录用日期:
2012
年
8
月
6
日
摘
要:
本研究主要以广西本地生长的优质的具自花结实李品种龙滩珍珠李为材料,根据李属雌蕊
S-RNase
基因的序列信息设计了一对特异性保守引物,对龙滩珍珠李基因组
DNA
进行
PCR
扩增,并
对扩增产物进行克隆、测序以及序列分析,成功克隆出两个
S-RNase
基因片段,其中一个为在中国李
中首次发现并命名为
PsS-RNase-
28
,大小为
727 bp
,其中包含有
367 bp
的内含子,仅有
360 bp
的外
显子编码序列,与黑刺李
(
Prunus spinosa
)
S-RNase S1
基因
( EF636467.1)
同源率达到
98%
,与其它李
S-
RNase
基因的同源率都达到
79%
以上。另一个扩增片段大小为
718 bp
,包含有
343 bp
的内含子,其中
仅有
375 bp
的外显子编码序列,与中国李
(
Prunus salicina
)
PsS-RNase
-
h
基因
(AB084148.1)
相似性达到
99%
,与其它李
S-RNase
基因的相似性都达到
85%
以上。这两个基因片段的获得及其后续分析有利于
进一步探讨龙滩珍珠李自花结实的分子机理。
关键词:
龙滩珍珠李;自交不亲和;自花结实;
S-RNase
(
S
)
基因
*
通讯作者。
Copyright © 2012 Hanspub
49
优良品种龙滩珍珠李自花结实分子机理初探
Copyright © 2012 Hanspub
50
1.
引言
蔷薇科果树如苹果、梨、杏、甜樱桃、李、扁桃、
梅等树种的大部分品种存在典型的配子体型自交不
亲和性。因此,生产上必须配置授粉树、虫媒授粉或
人工授粉以保证主栽品种获得较高的产量,这增加了
生产成本和难度,还浪费有限的土地资源。因而弄清
自交不亲和性的分子机理,鉴定果树品种的
S
基因型,
寻找或创造果树自交亲和性材料,以达到简化栽培方
式、降低果园生产成本、获得更高的经济效益的目的。
也正因为如此,对于果树自交不亲和的克服及其分子
机制的研究,成为当今研究的热点。陈迪新等
[1]
在不
同花龄,对“丰水”和“菊水”
2
个梨品种进行自花
授粉,发现蕾期授粉和延迟授粉都能显著提高它们自
花授粉的坐果率。李晓等
[2,3]
研究表明,甜樱桃自交不
亲和程度与其雌蕊的发育状态密切相关,蕾期授粉和
延迟授粉都能够在一定程度上克服甜樱桃的自交不
亲和性,但蕾期授粉的作用更加明显。蕾期或延迟授
粉克服自交不亲和的原因在于
S-RNase
在开花之前才
大量形成,开花后
S
-
RNase
的含量下降,花蕾中未成
熟的雌蕊不存在
S
-
RNase
或其含量极少,不具备开花
当天花朵的那种有效抑制花粉管伸长的能力。另外,
在各种自然或人为诱导条件下,自交不亲和性植物也
会因
S
基因发生突变丧失花粉或者花柱特异性识别,
进而表现自交亲和性。吴华清等
[4]
研究表明,“奥嗄二
十世纪”基因组中存在
S-RNase-
4
基因,但不能遗传
给后代。
Tao
等
[5]
发现果梅品种“
Benissashi
”和
“
Kensaki
”也表现自交亲和性,
Ushijima
等
[6]
进一步
研究发现这
2
个品种都含有发生突变了的
SFB
基因
(
SFBf
)
,该基因的编码区插入了约
6.8 kbp
的核苷酸序
列,导致
SFBf
的转录提前终止,从而使得携带该基
因的花粉不能被
Sf
-
RNase
识别和抑制,最后表现自交
亲和性,属于花粉突变体。
Vilanova
等
[7]
研究认为杏
品种
Canino
自交亲和的原因是该品种的花粉
S
基因
SFBc
发生了突变,即编码区插入了
358 bp
的核苷酸
残基。吴华清等
[8]
以“黄花梨”及其芽变品种“大果
黄花”为材料,研究表明,黄花和大果黄花是基于
S-RNase
基因的
S
基因型,发现两者均含有
S-RNase-
1
和
S-RNase-
2
基因;而且两品种的这
1
对雌蕊
S-RNase
基因均特异性地在花柱中表达,表达量没有明显差
异,表明“大果黄花”和“黄花”的雌蕊
S-RNase
基
因并无差异。由此他们推断:大果黄花的自交亲和性
突变,是由于花粉自交不亲和性功能丧失,从而表现
出自花授粉能够结实。配子体型自交不亲和性的二倍
体物种多倍体化会导致自交亲和,如人工诱导染色体
数目变异可以克服配子体型自交不亲和性。
Entani
等
[9]
将自交不亲和的矮牵牛
(
Petunia hyrbida
)PB
切段浸
泡在
0.02%
的秋水仙素,过夜,然后生根,获得了自
交亲和的四倍体
PF
。转基因方法创制自交亲和性品种
具有可行性,通过转入额外的
S
基因培育果树的自交
亲和品种,也可通过基因沉默的方式调节
S-RNase
的
表达量而产生自交亲和现象。
Broothaerts
等
[10]
对苹果
品种“早捷”
(Elstar)
进行转基因操作,转入了
35S
启
动子基因片段使花柱中的
S-RNase
基因发生基因沉
默,花柱中没有
S
-
RNase
的产生,从而获得自交亲和
性植株。
近年来,随着分子生物学技术的发展及各方面研
究的深入,自交不亲和性的分子机理研究取得了许多
突破性进展,极大地丰富了果树配子体型自交不亲和
性机理的学术理论
[4-10]
。目前国际上研究较多的和已
鉴定出
S
基因型的果树树种有苹果、甜樱桃、杏和梨
4
种,对于李的自交不亲和性和
S
基因型的研究已有
一些进展,黄绍西
(2009)
采用李属雌蕊
S
-
RNase
基因
特异性保守引物进行
PCR
和
RT-PCR
,并对扩增产物
进行克隆、测序和序列比对,在中国的
10
个传统李
品种中获得了
8
个符合雌蕊
S
特异性识别决定因子必
要条件的新基因,同时鉴定了中国李
25
个品种的
S
-
RNase
s
基因型。在中国的传统李品种中鉴定出了更
丰富的
S
-
RNase
基因多态性。并且认为中国李
S
-
RNase
-
h
在所研究样本中表现最高的基因频率,预
示着该基因可能与某些优良农艺性状有连锁关系。龙
滩珍珠李是广西科研单位与当地农民合作
10
年选育
出来的优质李新品种,树势生长旺,可自花结实,不
需要授粉树,适应性强,没有病虫害,产量高,对土
壤条件要求不高,在较为贫瘠的地段生长结果良好,
抗寒性也较好,是中国李中少有的能自花结实品质优
良的李品种。而对于像广西龙滩珍珠李能够自花结实
品质优良这样特殊的李品种
S
基因型的研究未见报
道。研究和鉴定龙滩珍珠李的
S
基因型,研究其如何
克服自交不亲和现象,从分子生物学的角度分析其自
交亲和的机理。龙滩珍珠李
S
基因型的鉴定,对于李
优良品种龙滩珍珠李自花结实分子机理初探
品种的遗传改良和栽培利用具有一定的指导意义,也
为丰富
S
基因的遗传信息,深入开展自交不亲和遗传
机制提供实验支持。
2.
材料与方法
2.1.
龙滩珍珠李
S
基因的克隆和
S
基因型的鉴定
2.1.1.
龙滩珍珠李总
DNA
的提取
本实验采用较适合于李叶片基因组
DNA
提取的
改良
CTAB
法,具体步骤如下:
1)
取约
0.3~0.5 g
新鲜叶片,剪成
5 mm
2
左右大
小放入研钵中,再加入少量液氮迅速研磨。加入
3
或
4
次液氮反复研磨,直至磨碎成细粉末状为止。
2)
将样品粉末装入
1.5 ml
离心管中,加
800
μ
l
CTAB
缓冲液
(
含
2% PVP)
,
24
μ
l
β
-
巯基乙醇。
3)
混匀后置
65
℃水浴中
30 min
,每隔
10 min
摇
匀一次。
4)
水浴后将样品在
12,000 rpm
离心
5 min
。
5)
吸取上清,加等体积的
CI(
氯仿
:
异戊醇
=
24:1)
,混匀后于摇床上摇匀
15 min
,
10,000 rpm
离心
10 min
。
6)
吸取上清,加等体积的
PCI(
苯酚
:
氯仿
:
异戊醇
= 25:4:1)
,上下颠倒混匀,
12,000 rpm
离心
10 min
。
7)
吸取上清后再次加等体积的
CI
抽提,
12,000
rpm
离心
10 min
。
8)
吸取上清加
2
倍体积无水乙醇,
1/10
体积的
NaAc
,沉淀
DNA
。
9) –20
℃静置
30 min
。
12,000 rpm
离心
15
min
,
用
70%
乙醇漂洗沉淀
2~3
次,室温下风干至无乙醇味
时
(30 min
左右
)
加
500 ml
无菌水溶解。
10)
加
10 mg/ml
的
Rnase
A(
天根公司
)1
μ
l
,
37
℃
水浴
1 h
。
11)
等体积的
CI
抽提,吸取上清加入
1/10
体积
的
NaAc
和
2
倍体积无水乙醇沉淀
DNA
,于
–20
℃静
置
30 min
。
12) 12,000 rpm
离心
15 min
,沉淀用
70%
乙醇洗
2~3
次,室温干燥,溶解于适量无菌水中
(20~50
μ
l)
。
13)
取
2
μ
l
样品在
0.8%
的琼脂糖凝胶上电泳,电
压
100 V
,
30 min
后结束电泳,将凝胶转移到溴化乙
锭溶液中染色
5 min
,紫外检测并拍照,观察主带是
否清晰,有无降解,是否含有
RNA
。
14)
取
2
μ
l DNA
溶液加纯水稀释至
400
μ
l
,紫外
分光光度计测定,
波长
200~300 nm
。根据
OD
260
/OD
280
估计其纯度,根据以下公式计算
DNA
的浓度。并用
无菌水调节
DNA
浓度为
50~100 ng/
μ
l
。
DNA
的质量浓度
=
50 × OD
260
×
稀释倍数
(
单
位:
mg/ ml )
。
2.1.2.
引物设计
蔷薇科李属
S-RNase
具有以下结构特点
(
图
1)
,与
其他蔷薇科
S-RNase
一样,具有
5
个保守区
(C1-C5)
和一个高变区
RHV(
或
HV)
,第一个内含子位于信号
肽和
C1
区之间,第二个内含子位于高变区,而高变
区
(RHV)
则位于
C2
和
C3
之间。
根据蔷薇科李属
S-RNase
结构特点,分别在
C2
上游区设计引物
Pru-C2(TGGCCAAGTAATTATTCAAACCC)
,在
C5
下游区设计引物
Pru-C5(CAAAATACCACTTCATGTAACAAC)
,用这
对引物对龙滩珍珠李基因组
DNA
进行
PCR
扩增。
2.1.3.
S
-
等位基因
PCR
扩增反应体系和反应条件
用引物
Pru-C2
与
Pru-C5
对龙滩珍珠李的基因组
DNA
进行扩增,其反应体系如下:
Template DNA 1.0
μ
l
10 × PCR Buffer 2.5
μ
l
MgCl
2
(25 mmol/L) 2.5
μ
l
dNTP Mixture(2.5 mmol/L) 2.5
μ
l
Primer 1: Pru-C2 (2.5 mmol/L) 0.625
μ
l
Primer 2: Pru-C5 (2.5 mmol/L) 0.625
μ
l
Taq DNA polymerase (5 U/
μ
l) 0.2
μ
l
ddH
2
O 15
μ
l
Total volume 25
μ
l
反应程序如下:
Step 1
94
℃
3 min
Step 2
94
℃
30 s
Step 3
56
℃
45 s 35 cycles
Step 4
72
℃
1 min 30 s
Step 5
72
℃
10 min
Step 6
4
℃
END
Cop
yright © 2012 Hanspub
51
优良品种龙滩珍珠李自花结实分子机理初探
Figure 1. The structure of
Prunus S-RNase
图
1.
李属
S-RNase
结构示意图
2.1.4.
凝胶上回收特异性
DNA
目的片段
对
PCR
产物进行琼脂糖凝胶电泳检测,获得
1
条特异性条带。采用
UNIQ-10
柱离心式
DNA
胶回收
试剂盒
(
上海生工
)
来回收此条带。操作步骤如下:
1)
用琼脂糖凝胶电泳将目的
DNA
片段与其他
DNA
尽可能分开,然后用干净的手术刀割下所要的
DNA
琼脂块,放入
1.5 ml
离心管中。
2)
按每
100 mg
琼脂糖凝胶加入
400
μ
l Binding
Buffer
,置于
50
℃
~60
℃水浴中
10 min
,使胶彻底融
化。加热融胶时,每
2 min
混匀一次。
3)
将
UNIQ-5
柱放入
2 ml
收集管中,将融化的
胶溶液转移到
UNIQ-5
柱中,室温放置
2 min
后,
8000
rpm
室温离心
1 min
。
4)
取下
UNIQ-5
柱,倒掉收集管中的废液,将
UNIQ-5
柱放入原收集管中,放入
450
μ
l Wash
Solution
,
8000 rpm
室温离心
1 min
。
5)
重复步骤
4
。
6)
去下
UNIQ-5
柱,倒掉收集管中的废液,将
UNIQ-5
柱放入原收集管中,让离心管盖敞开着,
12,000 rpm
室温下离心
1 min
。
7)
将
UN
IQ-5
柱放入一根新的
1.5 ml
离心管中,
柱膜中央加入
30
μ
l Elution Buffer
或水
(pH 7.0)
,室温
或
37
℃放置
2 min
,提高
DNA
的洗脱效率。
8) 12,000 rpm
高速离心
1 min
,离心管中的收集
液即含有回收的
DNA
片段,可立即使用或保存于
–20
℃备用。
2.1.5.
目的
DNA
片段与
T
载体的连接
采用
PUCm-T
载体
(
上海生工
)
,连接体系如下:
回收的
DNA
1
μ
l
PUCm-T Vector (50 ng/
μ
l) 1
μ
l
10 × Buffer 1
μ
l
T4 DNA ligase 1
μ
l
ddH
2
O 6
μ
l
Total volume 10
μ
l
混匀后,
20
℃
连接过夜。
2.1.6.
大肠杆菌感受态细胞的转化及阳性克隆的筛选
参照《分子克隆实验指南》
(
萨姆布鲁克等,
1996)
并加以改进。
1)
在
1.5 ml
离心管中将连接产物加入到
200
μ
l
感受态细胞
(
新鲜制备或
–70
℃保存均可
)
,轻轻摇匀。
2)
将离心管在冰上放置
20 min
后,放在
42
℃水
浴中热激
90 s
,立即把离心管放置在冰上冷却
2 min
。
3)
加入
800
μ
l LB
液体培养基
(
不含
AMP)
,混匀。
37
℃,
200 rpm
培养
45~60 min
。
4)
离心,去上清,将剩余的
150 ml
菌液重悬后
均匀涂布于
LB
平板上
(
用蓝白斑筛选法,平板表面涂
有
40
μ
l X-Gal
,
4
μ
l IPTG)
,室温放置
2 h
左右,
37
℃倒置培养
16 h
以上,直至出现单菌落。
X-Gal
储液
(20 ng/ml)
的配制为
0.02 g X-Gal
溶于
1 ml
的二甲基甲酰胺,外包铝或黑纸,以防光破坏,
储于
–20
℃;
IPTG
储液
(200 ng/ml)
配制为
1 ml
蒸馏水
中溶解
200 mg IPTG
后,用
0.22
μ
m
滤膜过滤除菌,
装于
eppendorf
管中。
2.1.7.
重组质粒的
PCR
鉴定
1)
配制
200 ml LB
液体培养基,
121
℃高压灭菌
20 min
后,冷却至
55
℃左右时加入
200
μ
l 100
μ
g/ml
的氨苄青霉素,混匀后分装于已灭菌的
eppendorf
管
中。
2)
待蓝白斑显色后挑取平板上的白色单菌落,放
入已冷却的
LB
液体培养基中,
37
℃,
200 rpm
过夜
(16
h
以上
)
震荡培养,至饱和状态
(OD
600
= 4
左右
)
。
3)
取
5
μ
l PCR
产物用相同引物和反应条件进行
再次
PCR
鉴定,反应体系如下:
菌液
1.0
μ
l
10 × PCR Buffer 2.5
μ
l
MgCl
2
(25 mmol/L) 2.5
μ
l
dNTP (2.5 mmol/L) 2.5
μ
l
Primer 1 (2.5 mmol/L) 0.625
μ
l
Primer 2 (2.5 mmol/L) 0.625
μ
l
Taq DNA polymerase (5 U/
μ
l) 0.2
μ
l
ddH
2
O 15
μ
l
Total volume 25
μ
l
Cop
yright © 2012 Hanspub
52
优良品种龙滩珍珠李自花结实分子机理初探
4)
用
1.5%
的琼脂糖电泳检测是否获得目的条
带,将获得目的条带的菌液直接由上海生工测序。
2.1.8.
特异片段序列分析
序列分析采用
DNAMAN
软件与
http://www.bio-soft.net/sms/index.html
在线分析软件,
然后在
NCBI
的
Ge
nBank
、
DDBJ
数据库与
http://www.ebi.ac.uk/Tools/clustalw
在线分析软件,进行
核
苷酸序列比对。用
DNAMAN
和
Clustalx 1.83
、
Clustalw
软件进行
S-RNase
序列的同源分析比对。
3.
结果与分析
3.1.
龙滩珍珠李
S
基因的克隆
3.1.1
龙滩珍珠李总
DNA
的提取
提取的龙滩珍珠李基因组
DNAOD
260
/OD
230
比值
大于
2.0
,
OD
260
/OD
280
比值约为
1.8
,质量好,产量高,
在
0.8%
琼脂糖凝胶电泳检测中,主带清晰,无降解现
象
(
图
2)
,完全可以满足后续
PCR
扩增要求。
3.1.2.
S
-
等位基因
PCR
扩增
本实验分别对
Taq
酶、模板
DNA
、
Mg
2+
、
dNTP
和引物的使用量进行调整,以使
PCR
反应体系达到最
优化,效果最佳。最终确定的最适合用量分别为:
Ta q
酶
1 U
、模板
DNA 1
μ
l
、
Mg
2+
、
2.5
μ
l
、
dNTP 2.5
μ
l
、
Primer 1 0.625
μ
l
、
Primer 2 0.625
μ
l
,共
25
μ
l
的反应
体系。
经过反复试验对
PCR
反应程序进行调整,特别是
对不同退火温度和时间以及循环数进行调整,以使
PCR
扩增效果最好。最终确定的最佳反应程序是:
94
℃预变性
3 min
,
94
℃变性
30 s
,
56
℃退火
45 s
,
72
℃延伸
1 min 30 s
,
35
个循环,
72
℃后延伸
10 min
。
建立了最佳
PCR
反应体系和反应程序,进行
PCR
扩增,结果如图
3
所示,得到
1
条清晰的特异扩增
Figure 2. Genome DNA of Longtanzhenzhuli by the CTA B method
1 and 2, PCR products; Lane M, -1500 marker
Figure 3. Allele –s
e
genes from
图
3.
龙
条带,
片段大小在
700 bp~00 bp
之间,接近
800 bp
。
3.1.3.
凝胶上回收特异性
DNA
片段
-10
柱式
DNA
凝胶
3.1.4.
重组质粒的
PCR
鉴定
,在位置
700 bp~800 bp
对应
1
3.2.
龙滩珍珠李
S
基因型的鉴定
龙滩珍珠李
S
-
RNase
基因的序列分析
数据库以及生
物软
接位点一样
(Finc
DL
pecific PCR amplification of
S-RNas
LongtanzhenzhuliLane
滩珍珠李
S
基因的
PCR
扩增
8
本实验使用上海生工生产的
UNIQ
回收试剂盒,对目的片段进行回收纯化,如图
4
所示,对电泳分离出的条带进行切割回收纯化,结果
条带较清晰,可以进行下一步实验。
如图
5
电泳检测结果显示
的出现了更为清晰的条带,说明目的片段已被转
入大肠杆菌中。将出现条带的菌液送至上海生工进行
测序,测序结果为
727 bp
和
718 bp
,
分别用
S-RNase
、
S-RN
ase
2
来表示。
1)
通过
NCBI
的
GenBank
、
DDBJ
件分析显示,珍珠
S-RNase-
1
片段在中国李中首次
发现并命名为
PsS-RNase-
28
,其中包含有
367 bp
的内
含子
(
见图
6)
,
仅有
360 bp
的外显子编码序列,编码
119
个氨基酸,通过
GeneBank
数据库的
BLAST
同源性分
析结果表明
(
表
1)
,与黑刺李
(
Prunus spinosa
)
S-RNase
S1
基因
(EF636467.1)
相似性达到
98%
,与其它李
S-
RNase
基因的相似性都达到
79%
以上。
与大多数植物在内含子
/
外显子剪
ham
,
1994
;
Beuppu
等,
2002)
,中国李
S-RNase
基因内含子的剪切位点也具有
GT/AG
保守序列,此
外,第一个剪切位点的左侧有两个相对保守的碱基
TG
,第二个剪切位点的右侧第一个碱基通常为
T
,附
近
4
个碱基富含
C
。
图
2.
龙滩珍珠李基因组
DNA
Cop
yright © 2012 Hanspub
53
优良品种龙滩珍珠李自花结实分子机理初探
Copyright © 2012 Hanspub
54
根据图
7
,对中国李的
S-RNase-
28
和黑刺李
S-RN
ase-
1
的
DNA
序列进行比较,结果表明,前者的内含子省略
了
117 bp
,在编码区两者只有
4
个碱基的差异。
在氨基酸序列上
(
图
8)
,中国李的
S-RNase-
28
和
黑刺李的
S-RNase-
1
两者没有区别。氨基酸序列相同,
其功能是否相同还需要进行进一步研究。
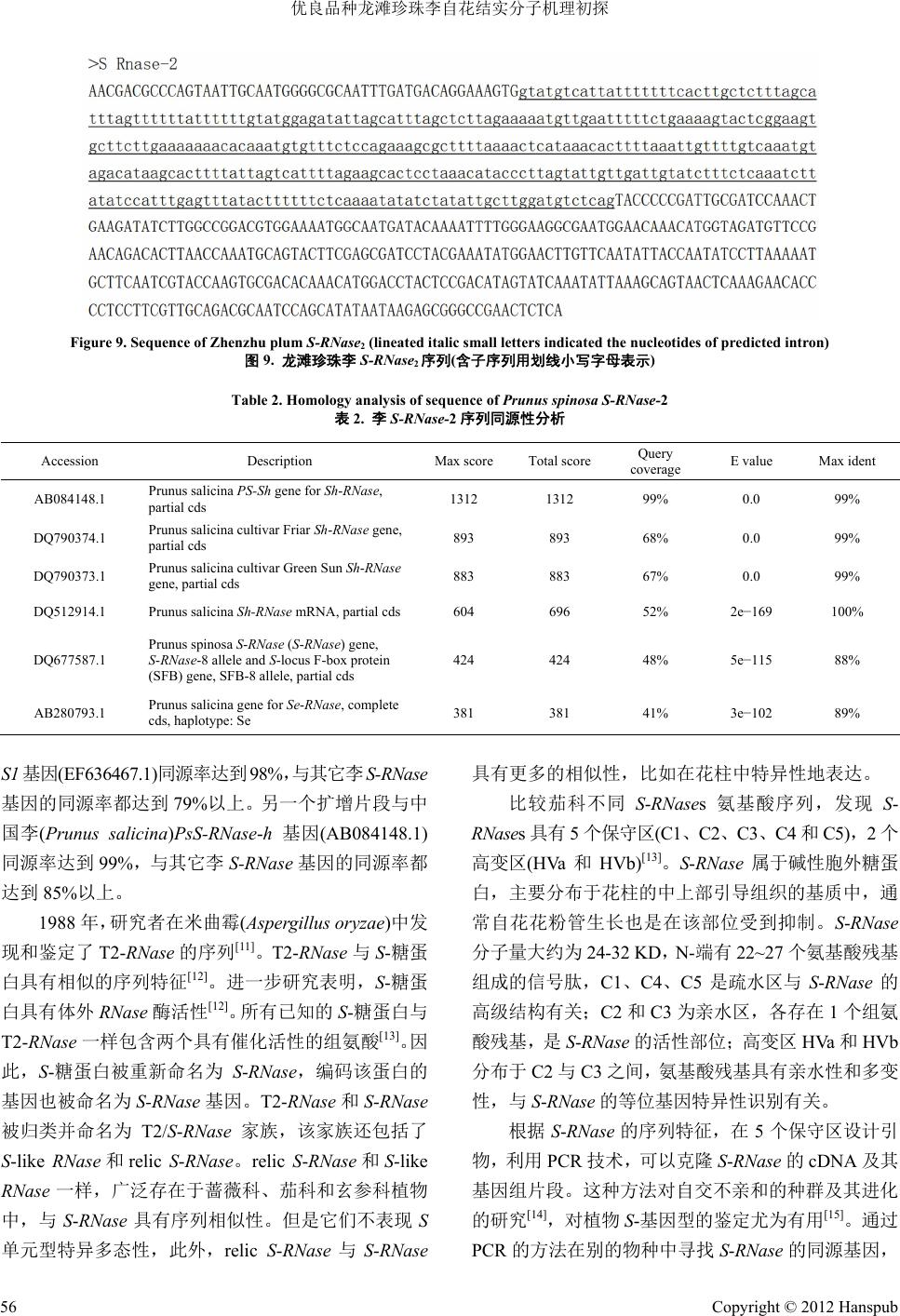
2)
龙滩珍珠李
S-RNase-
2
片段包含有
343 bp
的内含
子
(
见图
9)
,其中仅有
375 bp
的外显子编码序列,编码
124
个氨基酸,通过
GeneBank
数据库的
BLAST
同源性
分析结果表明
(
表
2)
,与中国李
(
Prunus salicina
)
PsS-
RNase-h
基因
(AB084148.1)
同源率达到
99%
,与其它李
S-RNase
基因的同源率都达到
85%
以上。
Figure 4. Electrophoresis detection of
PCR product
M
:
DL-1500 marker
图
4.
回收
PCR
产物的电泳检测
4
讨论与结论
Figure 5. The PCR detection of plamids M: DL marker
sequence oZhenzhu plum
S-RNase
1
(
PsS-RNase-
28)
Accession Description Max scoreTotal scoreQuery coverage E value Max ident
-1500
本研究采用
PCR
、克隆和测序的方法,在广西龙
滩珍珠李基因组
DNA
中获得了
2
个基因片段,通过
NCBI
进行序列比对和同源性序列搜索,分析了这
2
个基因片段,其中一个为在中国李中首次发现并命名
为
PsS-RNase-
28
,与黑刺李
(
Prunus spinosa
)
S-RNase
图
5.
转化菌落的
PCR
鉴定
Table 1.
Homology a nalysis off
表
1.
龙滩珍珠李
S-RNase
1
(
PsS-RNase-
28)
序列同源性分析
EF636467.1 Prunus spinosa
S
-locus
S
-
R
rtial cds
Nase
S1 gene, pa
846 1285 99% 0.0 98%
DQ677587.1
llele and
344 344 42% 4e
−
91 86%
Nase
-9 allele and
EU833958.1
allele
318 318 41% 2e
−
83 85%
DQ677586.1
se
-7-1 allele and
318 318 41% 2e
−
83 85%
DQ646488.1
and
311 311 40% 4e
−
81 85%
plete cds
se
-3-1 allele and
l cds
2 allele and
S-locus F-box protein (SFB) gene, SFB-3-2 allele, partial cds
Prunus spinosa
S
-
RNa s e
(
S
-
RNase
) gene,
S
-
RNase
-8 a
S-locus F-box protein (SFB) gene, SFB-8 allele, partial cds
Prunus salicina gene for
Sm
-
RNase
, partial cds AB093135.1
335 335 43% 2e
−
88 85%
AB093134.1 Prunus salicina gene for
Sl
-
RNas
e
, partial cds
324 324 42% 5e
−
85 85%
DQ677588.1
Prunus spinosa
S
-
RNase
(
S
-
RNase
) gene,
S-R
S-locus F-box protein (SFB) gene, SFB-9 allele, partial cds
P
runus spinosa
S
7-2
RNase
(
S
-
RNase
) gene,
S
-
RNase
-
S
7-2
322 322 40% 2e
−
84 86%
and SFB (SFB) gene, SFB7-2 allele, partial cds
P
runus spinosa
S
-
RNase
(
S
-
RNase
) gene,
S
-
RNa
S-locus F-box protein (SFB) gene, SFB-7-1 allele, partial cds
P
runus salicina haplotype Sb
S
-locus-F-box (SFB) gene
S
-
RNase
(
S
) gene, Sb allele, partial cds
Prunus salicina gene for
Sb
-
RNase
, com
AB252413.1
311 311 40% 4e
−
81 85%
DQ512912.1 Prunus salicina
S
7-
RNase
mRNA, partial cds
311 311 37% 4e
−
81 87%
DQ677584.1
Prunus spinosa S-
RNase
(
S
-
RNase
) gene,
S
-
RNa
S-locus F-box protein (SFB) gene, SFB-3-1 allele, partial cds
Prunus salicina cultivar wickson
Sk
-
RNase
gene, partial cds
309 309 62% 1e
−
80 79%
EU113311.1
305 305 42% 2e
−
79 84%
AB093133.1 Prunus salicina gene for
Sk
-
RNase
, partial cds
305 305 42% 2e
−
79 84%
AB084143.1 Prunus salicina
PS
-Sb
gene for
Sb
-
RNase
, partia
305 305 40% 2e
−
79 85%
DQ677585.1
P
runus spinosa
S
-
RNas e
(
S
-
RNase
) gene,
S
-
RNase
-3-
303 303 62% 7e
−
79 79%
优良品种龙滩珍珠李自花结实分子机理初探
Figure 6. Sequence of Zhenzhu plum
S-RNase
1
(
PsS-RNa se-
28)
(lineated italic small letters indicated the nucleotides of predicted intron)
图
6.
龙滩珍珠李
S-RNase
1
(
PsS-RNase-
28)
序列
(
内含子序列用划线小写字母表示
)
Figure 7. Aligment between Zhenzhu plum
S-RNase
1
(
PsS-RNase-
28) and
Prunus spinosa
S-RNase-
1
图
7.
龙滩珍珠李
S-RNase
1
(
PsS-RNase-
28)
和黑刺李
S-RNase-
1
的
DNA
序列比较
Figure 8. Aligment of the am
ino acid between Zhenzhu plum
S-RNase
1
(
PsS-RNase-
28) and
Prunus spinosa
S-RNase-
1
图
8.
龙滩珍珠李
S-RNase
1
(
PsS-RNase-
28)
和黑刺李
S-RNase-
1
的氨基酸序列比较
Cop
yright © 2012 Hanspub
55
优良品种龙滩珍珠李自花结实分子机理初探
Copyright © 2012 Hanspub
56
Figure 9. Sequence of Zhenzhu plum
S-RNase
2
(lineated italic small letters indicated the nucleotides of predicted intron)
图
9.
龙滩珍珠李
S-RNase
序列
(
含子序列用划线
小写字母表示
)
T
表
2.
李
S-RNase
序列同源性分析
Accession Description ore
Query
coverage
E value Max ident
2
able 2. Homology analysis of sequence of
Prunus spinosa S-RNase-
2
-
2
Max score Total sc
A
Prunus salicina
P
Sh
-
RNase
,
partial cds
B084148.1
S-Sh
gene for
1312 1312 99% 0.0 99%
DQ790374.1
Prunus salicina cultivar Friar
Sh
-
RNase
gene
partial cds
,
l cds
2e
AB280793.1
381 381 41% 3e
−
102 89%
893 893 68% 0.0 99%
DQ790373.1
Prunus salicina cultivar Green Sun
Sh
-
RNa s e
gene, partia
883 883 67% 0.0 99%
DQ512914.1 Prunus salicina
Sh-RNase
mRNA, partial cds
604 696 52%
−
169 100%
DQ677587.1
Prunus spinosa
S
-
RNa s e
(
S
-
RNase
) gene,
S
-
RNase
-8 allele and
S
-locus F-box protein
(SFB) gene, SFB-8 allele, partial cds
Prunus salicina gene for
Se
-
RNase
, complete
cds, haplotype: Se
424 424 48% 5e
−
115 88%
S1
基因
(EF636467.1)
同源率达到
98%
,
与其它李
S-RNase
者在米曲霉
(
Aspergillus oryzae
)
中发
现和
单元型特
异多态性,此外,
relic
S
-
RNase
与
S
-
RNase
具有更多的相似性,比如在花柱中特异性地表达。
白,
进化
的研
PCR
的方法在
别的物种中寻找
S
-
RNase
的同源基因,
基因的同
源率都达到
79%
以上。另一个扩增片段与中
国李
(
Prunus salicina
)
PsS-RNase
-
h
基因
(AB084148.1 )
同源率达到
99%
,与其它李
S-RNase
基因的同源率都
达到
85%
以上。
1988
年,研究
鉴定了
T2-
RNase
的序列
[11]
。
T2 -
RNase
与
S
-
糖蛋
白具有相似的序列特征
[12]
。进一步研究表明,
S
-
糖蛋
白具有体外
RNase
酶活性
[12]
。所有已知的
S
-
糖蛋白与
T2-
RNase
一样包含两个具有催化活性的组氨酸
[13]
。因
此,
S
-
糖蛋白被重新命名为
S
-
RNase
,编码该蛋白的
基因也被命名为
S-RNase
基因 。
T2-
RNase
和
S
-
RNase
被归类并命名为
T2/
S
-
RNase
家族,该家族还包括了
S
-like
RNase
和
relic
S
-
RNase
。
relic
S
-
RNase
和
S
-like
RNase
一样,广泛存在于蔷薇科、茄科和玄参科植物
中,与
S
-
RNase
具有序列相似性。但是它们不表现
S
比较茄科不同
S
-
RNase
s
氨基酸序列,发现
S
-
RNase
s
具有
5
个保守区
(C1
、
C2
、
C3
、
C4
和
C5)
,
2
个
高变区
(HVa
和
HVb)
[13]
。
S
-
RNase
属于碱性胞外糖蛋
主要分布于花柱的中上部引导组织的基质中,通
常自花花粉管生长也是在该部位受到抑制。
S
-
RNase
分子量大约为
24-32 KD
,
N-
端有
22~27
个氨基酸残基
组成的信号肽,
C1
、
C4
、
C5
是疏水区与
S
-
RNase
的
高级结构有关;
C2
和
C3
为亲水区,各存在
1
个组氨
酸残基,是
S
-
RNase
的活性部位;高变区
HVa
和
HVb
分布于
C2
与
C3
之间,氨基酸残基具有亲水性和多变
性,与
S
-
RNase
的等位基因特异性识别有关。
根据
S
-
RNase
的序列特征,在
5
个保守区设计引
物,利用
PCR
技术,可以克隆
S
-
RNase
的
cDNA
及其
基因组片段。这种方法对自交不亲和的种群及其
究
[14]
,对植物
S
-
基因型的鉴定尤为有用
[15]
。通过
优良品种龙滩珍珠李自花结实分子机理初探
可以初步认为该物种也具有
S
-
RNase
介导的配子体型
自交不亲和性现象。比如,通过这种方法其他学者在
金鱼草植物
(
Antirrhinum hispanicum
)
克隆了
S
-
RNase
同
源基因的
cDNA
,并发现这个物种也表现配子体自交
不亲和性。
S-RNase
的系统发育分析提示不同科植物
间的配子体自交不亲和起源于共同的祖先。
虽然中国李
S-RNase-
28
和黑刺李
S-RNase-
1
的氨
基酸序列相似性高达
100%
,但前者的内含子省略了
117 bp
,在编码区两者也有
4
个碱基的差异。张树军
等
[6]
对中国李
S-RNase-
26
与杏
S-RNase-
2
进行了序列
比对
的同源性,这说明
S-RN
智年博士。
参考
长
特性
[J].
南京农业大学学报
, 2004, 27(3): 34-37.
[2]
李晓
,
吴华清
,
吴俊
,
陶书田
,
张绍铃
.
不同发育阶段甜樱桃
长的差异
[J].
果树学报
, 2008,
apricot
4,
: 955-957.
87: 732-735.
: 304-311.
分析
,结果在编码区仅有
2
个碱基差异,而在内
含子区域,无论长度还是碱基组成,二者差异都比较
大,两者推导的氨基酸序列相似性达到
99%
,具有相
同的高变区
(RHV)
。在甜樱桃中也报道了相似的结果。
Zisovich
等
[16]
在西洋梨
(
Pyrus communis
)
上更进一步
的证明了具有相同高变区
(RHV)
的
S
-
RNase-i
和
S
-
RNase-n
基因具有不同的等位基因特异性识别。因
此,即使是氨基酸序列相似性高达
100%
,其功能是
否一样还需要进一步的研究。
龙滩珍珠李具有自花结实而且品质优良,本研究
成功克隆得到的两个基因片段中,一个为曾被报道过
的中国李
S-RNas e-h
,另一个新基因与曾被报道过的
李属其它
S-RNases
基因有高度
ases
氨基酸序列中从
C2
到
C5
的区域是正常的,
导致其自花结实的原因很可能在于
C1
之前或
C5
之后
的区域出现了不同于其他李品种的变化,产生了变
异,具体原因有待于进一步研究。
6.
致谢
感谢广西农科院作物遗传改良生物技术重点实
验室及魏源文博士、黄诚梅博士、邓
文献
(References)
[1]
陈迪新
,
张绍铃
,
陶书田
.
沙梨花粉原位萌发与花粉管生
花授粉后花粉萌发和花粉管生
25(4): 501-505.
[3]
李晓芳
,
李茂福
,
韩振海
,
许雪峰
,
李天忠
. “
鸭梨
”
芽变
“
闫庄
梨
”
自交亲和性分子机制初步研究
[J].
园艺学报
, 2007, 35(1):
8. 13-1
[4]
吴华清
,
齐永杰
,
张绍铃
. “
奥嗄二十世纪
”
梨自交亲和性分子
机制及其遗传特性研究
[J].
园艺学报
, 2008, 35(8): 1009-1116.
吴华清
衡伟
,
李晓
[5]
, ,
黄绍西
,
张绍铃
. “
大果黄花
”
梨自交亲和
性变异机制研究
[J].
南京农业大学学报
, 2007, 30(2): 29-33.
[6]
R. Tao, T. Habu, A. Namba, H. Yamane, F. Fuyuhiro, K.
Iwamoto and A. Sugiura. Inheritance of
S
f
-
RNase
in Japanese
apricot (
Prunus mume
) and its relation to self-compatibility.
Theoretical and Applied Ge
netics, 2002, 105: 222-228.
[7]
K. Ushijima, H. Yamane, A. Watari, E. Kakehi, K. Ikeda, N. R.
Hauck, A. F. Iezzoni and R. Ta
o. The S haplotype-specific F-box
protein gene, SFB, is defective in self-compatible haplotypes of
Prunus avium
and
P. mume
. Plant Journal, 2004, 39: 573-586.
[8]
S. Vilanova, M. L. Badenes, L. Burgos, J. Martinez-Calvo, G.
Llacer, and C. Romero. Self-compatibility of two
selections is associated with
two pollen-part mutations of
different nature. Plant Physiology, 2006, 142: 629-641.
[9]
W. Broothaerts, J. Keumans and I. V. Nerum. Self-fertile apple
resulting from
S-RNase
gene silencing. Plant Cell Reports, 200
22: 497-501.
[10]
黄绍西
.
中国李
(
Prunus salicina
)
自交不亲和性与中国樱桃
(
P.
pseudocerasus
)
自交亲和性机制的初步研究
[D].
南京
:
南京
农业大学
, 2009.
[11]
Y. Kawata, et al. Amino-acid sequence of ribonucease T2 from
Aspergillus oryzae
. European Journal of Biochem, 1988, 103:
408-418.
[12]
B. A. Mcclure, V. Haring, P. R. Ebert, et al. Style self-income-
patibility gene products of nicotiana alata are ribonucleases.
Nature, 1989, 342
[13]
T. R. Ioerger, A. G. Clark, T.-H. Kao. Polymorphism at the
self-incompatibility locus in
Solanaceae predates
speciation.
Proc Natl Acad Sci USA, 1990,
[14]
A. D. Richman, W. Broothaerts, J. R. Kohn. Self-incompatibility
Rnase
from the three plant families: Homology or convergence.
Am J Bot, 1997, 84: 912-917.
[15]
R. Tao, H. Yamane, H. Sassa, et al. Identification of stylar
RNase
associated with gametophytic self-incompatibility in almond
(
Pyrus dulcis
). Plant Cell Physiology, 1997, 38
[16]
张树军
,
黄绍西
,
张绍铃等
.
携李等
15
个李品种
S
基因型鉴
定及其多态性分析
[J].
果树学报
, 2008, 25(3): 338-342.
Cop
yright © 2012 Hanspub
57