 Hans Journal of Surgery 外科, 2012, 1, 11-16 http://dx.doi.org/10.12677/hjs.2012.13003 Published Online October 2012 (http://www.hanspub.org/journal/hjs.html) AFP Specific Liver Tumor Vaccines Inhibit Hepatocellular Carcinoma Growth Yang Liu1, Yueru W ang2, Lisa H. Butterfield3 1Hepatobiliary Surgery Division, Tenth People’s Hospital, Tongji University, Shanghai 2Internal Medicine Division, First People’s Hospital, Shanghai Jiao Tong University, Shanghai 3Department of Surgery and Immunology, Cancer Institute of Pittsburgh University, Pittsburgh, USA. Email: yliu6633@yahoo.com.cn Received: Jul. 14th, 2012; revised: Jul. 26th, 2012; accepted: Aug. 1st, 2012 Abstract: Objective: Alpha-fetoprotein (AFP), a tumor-associated antigen for hepatocellular carcinoma (HCC), is an established biomarker for hepatocellular cancer (HCC). In this study, we detected antitumor activity of AFP specific T cells in vitro. Methods: We created a lentivirus expressing AFP (Lenti-AFP) and investigated the antitumor activity of AFP-specific CD4+ and CD8+ T cells(liver tumor vaccines), which were activated by either AFP peptide-pulsed or Len- ti-AFP-engineered DC in vitro. Results: Our research demonstrated that the AFP-specific T cells could efficiently kill HepG2 cells, by significantly increasing the level of IL-2, IFN-γ、TNF-α、perforin and granzyme B, as well as by in- hibiting the production of IL-10 (a negative regulator of T cell activation). Conclusion: AFP-specific CD4+ and CD8+ T cells (liver tumor vaccines) which were activated by either AFP peptide-pulsed or Lenti-AFP-engineered DC have ob- viously antitumor activity. This study provides new insight into the design of DC activated antigen-specific T cell-based clinical trials Keywords: Alpha-Fetoprotein (AFP); Dendritic Cell; Cytokine; AFP-Specific T Cells AFP 特异性肝癌疫苗抑制肝细胞癌生长 刘 扬1,王跃如 2,Lisa H. Butterfield3 1同济大学附属第十人民医院肝胆外科,上海 2上海交通大学附属第一人民医院内科,上海 3美国匹兹堡大学肿瘤研究所,外科及免疫学组,匹兹堡,美国 Email: yliu6633@yahoo.com.cn 收稿日期:2012 年7月14 日;修回日期:2012 年7月26 日;录用日期:2012 年8月1日 摘 要:目的:甲胎蛋白作为一种肿瘤特异性抗原,是临床上诊断肝癌的重要指标。本研究中,我们检测了 AFP 特异性 T细胞的体外抗肿瘤免疫活性。方法:我们采用经过 AFP 抗原决定簇肽刺激或慢病毒转染的树突状细胞 (dendritic cells, DC),活化AFP 特异性的 CD4+T细胞和 CD8+T细胞(肝癌疫苗),并在体外检测了这群T细胞对 人肝癌细胞HepG2 的杀伤活性。结果:本研究结果显示,AFP 特异性的 CD4+T细胞和CD8+T细胞体外能有效 杀伤 HepG2 细胞,上调培养体系中 IL-2、IFN-γ、TNF- α、穿孔素、颗粒酶的水平,对于T细胞活化的负调控因 子IL-10,具有显著的抑制作用。结论:经DC活化后的 AFP特异性 T细胞具有显著的体外抗肿瘤免疫活性, 为肝细胞癌的临床免疫治疗提供了依据。 关键词:甲胎蛋白;树突状细胞;细胞因子;AFP 特异性 T细胞 1. 引言 肝细胞癌(HCC)位居国人恶性肿瘤发病率第二 位,年死亡人数超过 30 万人,只有少数患者(<20%) 适合手术治疗,因此应用肝癌特异性抗原的肿瘤疫苗 Copyright © 2012 Hanspub 11  AFP 特异性肝癌疫苗抑制肝细胞癌生长 治疗成为当前研究的热点。甲胎蛋白(AFP)是肝细 胞 癌特异性抗原,约 60%~70%的HCC 患者血清 AFP 不同程度升高。 树突状细胞(DC)是目前公认的体内抗原递呈功 能最强的专职抗原递呈细胞,能有效的激活幼稚 T细 胞,处于启动、调控并维持免疫应答的中心环节,在 机体抗肿瘤免疫中具有十分重要的作用。临床研究显 示,实体肿瘤的 DC 浸润与预后较好有关,而 DC 受 损则免疫抑制增强且肿瘤恶化。Butterfield等[1,2]鉴定 出一系列可被人 T细胞识别的 HLA-A*0201 限制性 AFP 免疫决定簇肽和亚免疫决定簇肽,并应用于I/II 期临床实验(安全性和有效性)。本文作者[3,4]随后报道 了使用 AFP 抗原决定簇肽和亚决定簇肽片段刺激 DC,能有效诱发抗肝癌的特异性细胞免疫反应。有研 究显示,CD4+辅助性 T细胞可能通过促进CD8+T细 胞扩增并增加肿瘤杀伤性细胞因子的释放,发挥潜在 的抗肿瘤免疫作用[5]。 在前期研究工作的基础上[6],我们应用 HLA- A*0201 限制性AFP抗原决定簇肽和亚决定簇肽以及 慢病毒转染的 AFP 刺激的树突状细胞与人外周血 T 细胞共同培养,在体外人肝癌细胞株模型上验证了 AFP 抗原特异性T细胞的抗肿瘤免疫活性,为临床治 疗肝细胞癌提供了实验基础,现报道如下。 2. 材料与方法 2.1. 健康捐献者 通过标准的 HLA血清分型及基因分型方法确定 所有捐献者均为 HLA-A2 阳性(血液来自上海市中心 血站自愿献血者)。 2.2. 多肽合成 多肽由上海生工生物工程技术有限公司合成 (Shanghai, P.R.China),用氨基酸分析和质谱证实了氨 基酸的序列正确,多肽纯度 > 95%。 2.3. 肝癌细胞株 表达 HLA-A2+的HCC 细胞株(AFP阳性)购自美 国BD 公司。 2.4. 统计分析 用SPSS 11.0统计软件处理各组数据,实验结果 以mean ± SD 表示,采用配对t检验,p < 0.05 为具有 统计学意义。 2.5. AFP慢病毒载体的构建 以pWPXL-MOD2 为骨架,将目的基因构建到双 GFP 表达的慢病毒载体中,替换其中一个GFP,形 成 AFP 与GFP 共表达的慢病毒载体。使用 PCR 方法从 HepG2 细胞基因组中获取 AFP 基因片段,引入 BamH1 和Sal1酶切位点。对空载体pWPXL-MOD2 和PCR 的回收产物用 BamH1和Sa l1进行双酶切及连接反应。 采用 ProFection Mammalian Transfection System制备 慢病毒穿梭质粒及其辅助包装元件载体质粒。简言 之,慢病毒包装细胞转染用胰蛋白酶消化对数生长期 的293T 细胞,细胞密度为0.5 × 109/L 时,重新接种 于25 mL 的15 cm细胞培养皿,于 37℃,50 mL/L CO2 培养箱内培养。制备慢病毒包装系统中四种质粒 DNA 溶液(pRSv-REV 10 ug,pMD 1 g~pRRE 15 ug,pMD2G 7.5 ug,AFP 质粒20 ug),定容至 1800 ul,再加入 CaCl2 (2.5 mol/L)200 ul,混匀后加入2XHEBS 缓冲盐溶液 2000 ul,室温放置 20~30 min;当细胞密度达60%~70% 时转染。将 DNA 和磷酸钙混合液转移至含单层细胞 的培养液中,混匀,培养 12 h 后弃去含有转染混和物 的培养液,加入PBS 15 mL,轻摇后弃去,重复该步 骤3次;每瓶细胞中加入含 100 mL/L胎牛血清的细 胞培养液 15 mL,继续培养 48 h。收集转染 72 h的293T 细胞上清液;将收集的上清液于4℃,4000 g离心 10 min,10 cm/r,收集上清液;将上清液以0.45 μm滤 器过滤;于40 mL 超速离心管中,4℃,72,000 g/min 离心 120 min;500 ul PBS 重悬病毒沉淀。 2.6. DC的分离和培养 DC 的获取如文献所述,通过静脉穿刺抽取外周 血,通过 Ficoll 密度梯度离心法分离外周血单核细胞 (PBMC)。PBMCs(3~4 × 107)接种于 T-25 培养瓶中, 加入含抗生素及5%~10%人AB 血清的 RPMI1640 培 养基,于 37℃培养箱中培养 2 h。用PBS 轻轻漂洗, 去除未贴附的细胞,已贴附的细胞在含有 800 U/mL GM-CSF 和500 U/mL IL-4 的培养基中培养7 d。冲 洗 后收集未贴附及贴附较弱的细胞(DC)。形态学和基因 分型检测显示,这些细胞中一般含有 30%~50%的DC。 Copyright © 2012 Hanspub 12  AFP 特异性肝癌疫苗抑制肝细胞癌生长 2.7. 从PBMC 中分离 CD8+T,CD4+T淋巴 细胞 通过免疫磁珠法分别分离出 CD8+T细胞和 CD4+T细胞。流式细胞术分析结果显示,分离后得到 的CD8+T细胞,CD4+T细胞的纯度均大于 98%[3]。 2.8. 通过 AFP 抗原决定簇肽刺激以及慢病毒转 染DC 获得AFP 特异性的 CD8+T和 CD4+T细胞 通过 AFP抗原肽刺激DC 获得 AFP特异性的 CD8+T和CD4+T细胞的方法如前人所述。简言之, 室温下用含 10 μg/mL AFP 抗原肽的无血清 RPMI 1640 培养液刺激 HLA-A2+样本来源的 DC 120 min。 DC 经离心计数后,按1:20 的比例与自体同源CD8+T 细胞或 CD4+T细胞于24 孔板中共培养1周,RPMI 1640 培养液中含有抗生素,10% 人AB 血清及 10 ng/mL IL-7,每 3~4 d 添加IL-2 10 U/mL。培养一周后, 计数未贴附的细胞,用以抗原肽处理后的 DC 再刺激 1周。刺激两次后,收集细胞检测。在含 2%人AB 血 清的 RPMI1640 培养基中用携带人 AFP 全长 cDNA 的 重组慢病毒表达载体Lenti-AFP 转染 DC 2 h。转染后 的DC 以1 × 105 cells/mL 的密度铺板,与 CD8+T细胞 或CD4+T细胞共培养于含有 10%人AB血清及 25 ng/mL IL-7的培养液中(DC:T = 1:20),每隔 3 d补充 10 U/mL。 2.9. AFP特异性的 CD8+T和CD4+T淋巴细胞对 HepG2 肝癌细胞的体外杀伤活性 将上一步得到的 CD8+T细胞单独或连同 CD4+T 细胞,与 HepG2 细胞共同培养(CD8+T细胞:HepG2 = 1:1)于96孔板中,总体积 200 µL,并设淋巴细胞和靶 细胞对照,每实验组设3个复孔。培养 18 h 后加入 5 mg/mL MTT 10 µL,继续培养 6 h,弃上清,加DMSO 100 µL,于 570 nm 处测定吸光度值(A ),杀伤率 = [1 – (实验孔 A值–淋巴细胞对照孔 A值)/靶细胞对照孔 A值] × 100%。 2.10. 淋巴细胞增殖的测定 制备细胞悬液,加入到96 孔培养板中(2 × 105个/ 孔),每实验组设3个复孔。细胞加入 20 µL 1 µ Ci/孔 的[3H]胸腺嘧啶,于 37℃、5%CO2孵育箱中培养 6 h 后,用细胞收集仪将细胞收集至玻璃纤维膜上。加闪 烁液后,用液闪仪测定每分钟脉冲数(cpm),以刺激指 数(SI)表示增殖程度:SI = 实验孔 cpm/对照孔 cpm。 2.11. 细胞因子的检测 将上述单细胞悬液加入到96 孔培养板中(2 × 105 个/孔),每实验组设3个复孔。细胞于于37℃、5%CO2 孵育箱中培养 24 h 后,收集上清,用双抗体夹心 ELISA 法检测上清中细胞因子的水平。方法如下:1) 包被:将稀释好的一抗加入96 孔酶标板中,每孔100 µL,4℃过夜;2) 封闭:用含0.05%吐温–20 的PBS 洗3次后,加入 200 µL含10%FCS 的PBS,室温封 闭2 h;3) 加样:加入100 µL标准品或样品,4℃过 夜;4) 加二抗:每孔加入100 µL稀释后的二抗,室 温放置 1 h;5) 加酶:每孔加入 100 µL 稀释后的辣根 过氧化物酶,室温放置1 h;6) 加底物:将底物TMB 稀释为 0.1 mg/mL,加入0.1%H 2O2,每孔 100 µL;8) 显色:室温显色 30 min 后,加入 2 M H2SO4终止反应, 于450 nm处测吸光度值,校正波长为570 nm,根据 标准曲线换算出培养上清中细胞因子的含量,结果用 pg/mL表示。 3. 结果 3.1. AFP抗原肽刺激及慢病毒转染处理后的 DC 能显著促进 T细胞增殖 本实验体系中,以人HepG2肝癌细胞 cDNA为 模板,PCR 扩增出 AFP 基因片段,构建成Lenti-AFP 慢病毒载体或阴性对照病毒Lenti-LacZ(mock)。以该 慢病毒载体转染293T 细胞,包装慢病毒表达载体, 再以包装好的慢病毒体外感染 DC。首先,我们检测 了慢病毒转染率和 AFP表达情况。如图 1(A)所示, 以18SRNA 为内参,AFP过表达率为(5.31 ± 0.11)。 同时也用蛋白印记分析的方法对细胞裂解物中 AFP 的含量进行验证。结果如图1(A)和1(B)所示。 (A) AFP在慢病毒转染 DC 中的基因表达结果, 以18SRNA 为内参。(B) AFP的蛋白印记分析结果。 结果以 mean ± SD 表示,与阴性对照相比, **p < 0.01, 实验重复3次,得到类似结果。 Copyright © 2012 Hanspub 13 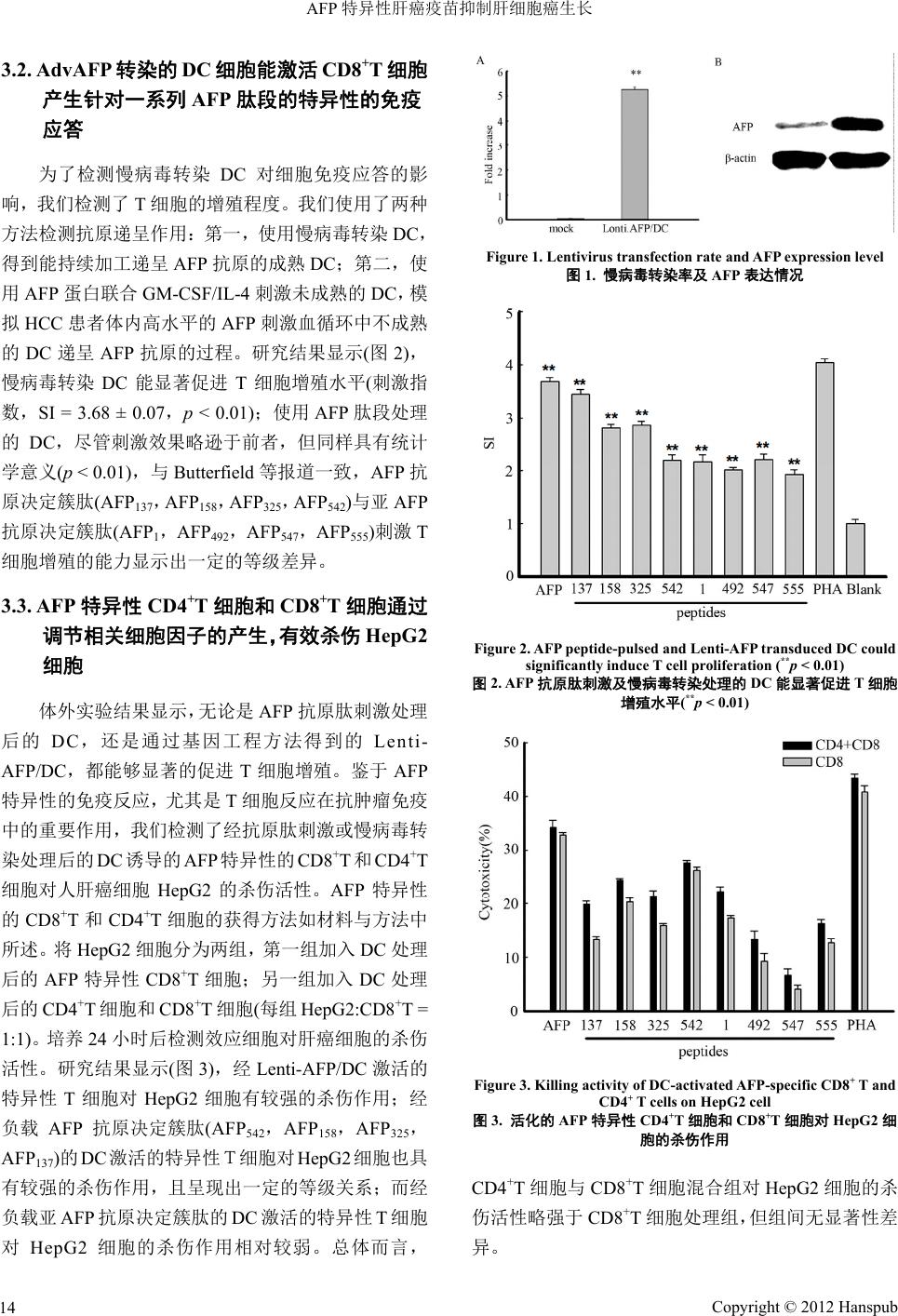 AFP 特异性肝癌疫苗抑制肝细胞癌生长 3.2. AdvAFP转染的 DC 细胞能激活 CD8+T细胞 产生针对一系列 AFP 肽段的特异性的免疫 应答 为了检测慢病毒转染 DC 对细胞免疫应答的影 响,我们检测了 T细胞的增殖程度。我们使用了两种 方法检测抗原递呈作用:第一,使用慢病毒转染 DC, 得到能持续加工递呈AFP抗原的成熟DC;第二,使 用AFP 蛋白联合GM-CSF/IL-4 刺激未成熟的 DC,模 拟HCC患者体内高水平的 AFP刺激血循环中不成熟 的DC 递呈 AFP抗原的过程。研究结果显示(图2), 慢病毒转染 DC 能显著促进 T细胞增殖水平(刺激指 数,SI = 3.68 ± 0.07,p < 0.01);使用 AFP 肽段处理 的DC,尽管刺激效果略逊于前者,但同样具有统计 学意义(p < 0.01),与 Butterfield 等报道一致,AFP 抗 原决定簇肽(AFP137,AFP158,AFP325,AFP542)与亚 AFP 抗原决定簇肽(AFP1,AFP492,AFP547,AFP555)刺激 T 细胞增殖的能力显示出一定的等级差异。 3.3. AFP特异性 CD4+T细胞和 CD8+T细胞通过 调节相关细胞因子的产生,有效杀伤 HepG2 细胞 体外实验结果显示,无论是 AFP 抗原肽刺激处理 后的 DC,还是通过基因工程方法得到的 Lenti- AFP/DC,都能够显著的促进 T细胞增殖。鉴于 AFP 特异性的免疫反应,尤其是 T细胞反应在抗肿瘤免疫 中的重要作用,我们检测了经抗原肽刺激或慢病毒转 染处理后的DC 诱导的 AFP 特异性的CD8+T和CD4+T 细胞对人肝癌细胞 HepG2的杀伤活性。AFP 特异性 的CD8+T和CD4+T细胞的获得方法如材料与方法中 所述。将HepG2 细胞分为两组,第一组加入 DC 处理 后的 AFP 特异性CD8+T细胞;另一组加入 DC 处理 后的 CD4+T细胞和CD8+T细胞(每组 HepG2:CD8+T = 1:1)。培 养24 小时后检测效应细胞对肝癌细胞的杀伤 活性。研究结果显示(图3),经 Lenti-AFP/DC 激活的 特异性 T细胞对 HepG2 细胞有较强的杀伤作用;经 负载 AFP 抗原决定簇肽(AFP542,AFP158,AFP325, AFP137)的DC激活的特异性T细胞对 HepG2 细胞也具 有较强的杀伤作用,且呈现出一定的等级关系;而经 负载亚 AFP 抗原决定簇肽的 DC 激活的特异性 T细胞 对HepG2细胞的杀伤作用相对较弱。总体而言, Figure 1. Lentivirus transfection rate and AFP expression level 图1. 慢病毒转染率及 AFP 表达情况 Figure 2. AFP peptide-pulsed and Lenti-AFP transduced DC could significantly induce T cell proliferation (**p < 0.01) 图2. AFP抗原肽刺激及慢病毒转染处理的 DC 能显著促进 T细胞 增殖水平(**p < 0.01) Figure 3. Killing activity of DC-activated AFP-specific CD8+ T and CD4+ T cells on HepG2 cell 图3. 活化的 AFP 特异性CD4+T细胞和 CD8+T细胞对 HepG2细 胞的杀伤作用 CD4+T细胞与 CD8+T细胞混合组对HepG2 细胞的杀 伤活性略强于CD8+T细胞处理组,但组间无显著性差 异。 Copyright © 2012 Hanspub 14 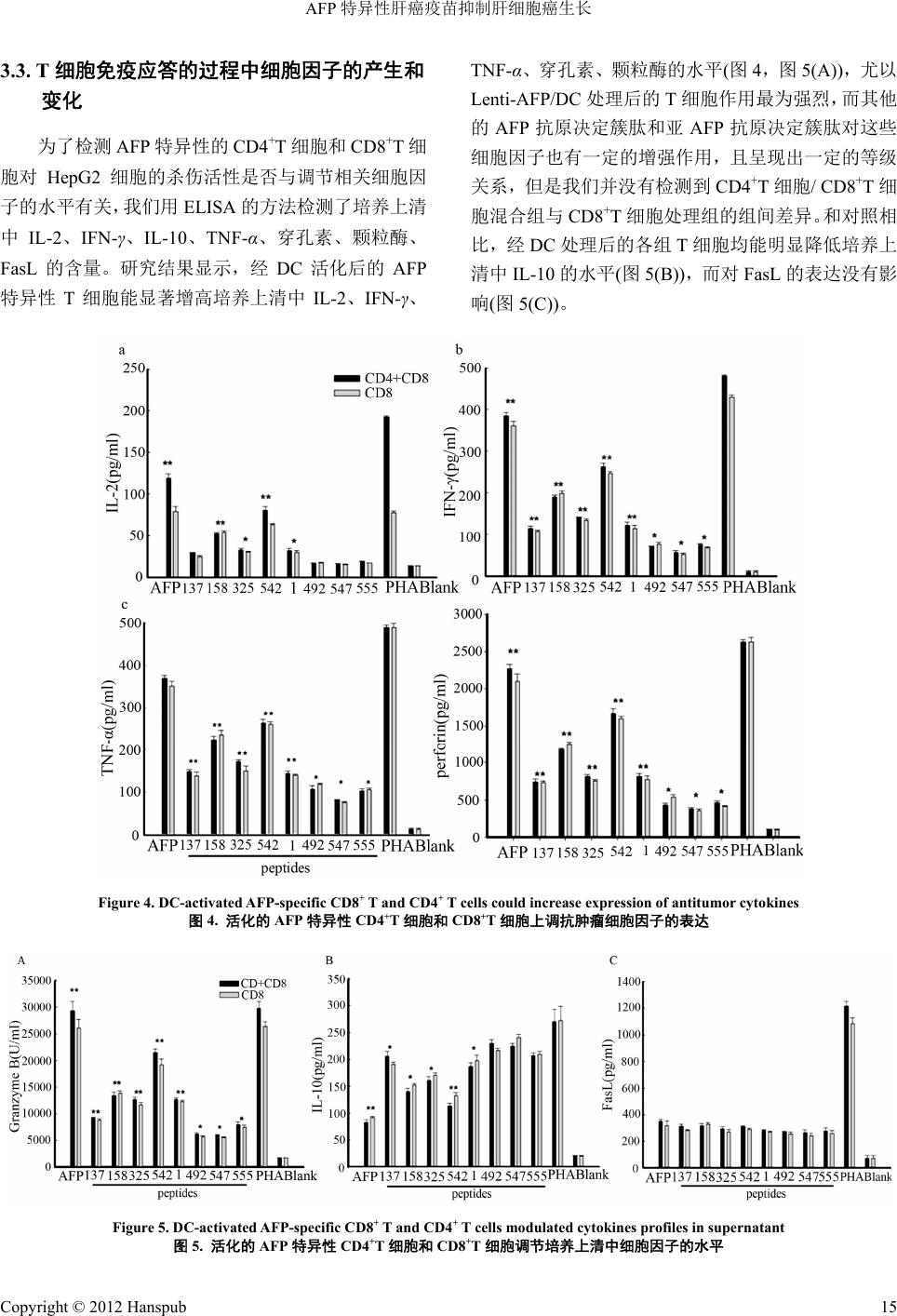 AFP 特异性肝癌疫苗抑制肝细胞癌生长 Copyright © 2012 Hanspub 15 3.3. T细胞免疫应答的过程中细胞因子的产生和 变化 TNF-α、穿孔素、颗粒酶的水平(图4,图5(A)),尤以 Lenti-AFP/DC 处理后的T细胞作用最为强烈,而其他 的AFP 抗原决定簇肽和亚 AFP 抗原决定簇肽对这些 细胞因子也有一定的增强作用,且呈现出一定的等级 关系,但是我们并没有检测到 CD4+T细胞/ CD8+T细 胞混合组与 CD8+T细胞处理组的组间差异。和对照相 比,经 DC 处理后的各组T细胞均能明显降低培养上 清中 IL-10 的水平(图5(B)),而对 FasL 的表达没有影 响(图5(C))。 为了检测AFP 特异性的 CD4 +T细胞和 CD8+T细 胞对 HepG2 细胞的杀伤活性是否与调节相关细胞因 子的水平有关,我们用 ELISA 的方法检测了培养上清 中IL-2、IFN-γ、IL-10、TNF-α、穿孔素、颗粒酶、 FasL 的含量。研究结果显示,经 DC活化后的 AFP 特异性 T细胞能显著增高培养上清中 IL-2、IFN-γ、 Figure 4. DC-activated AFP-specific CD8+ T and CD4+ T cells could increase expression of antitumor cytokines 图4. 活化的 AFP 特异性CD4+T细胞和 CD8+T细胞上调抗肿瘤细胞因子的表达 Figure 5. DC-activated AFP-specific CD8+ T and CD4+ T cells modulated cytokines profiles in supernatant 图5. 活化的 AFP 特异性CD4+T细胞和 CD8+T细胞调节培养上清中细胞因子的水平  AFP 特异性肝癌疫苗抑制肝细胞癌生长 4. 讨论 抗原递呈是T细胞活化过程中的一个重要步骤, 树突状细胞(DC)是功能强大的专职抗原递呈细胞,是 机体免疫的起始细胞[7]。近年来,发现肿瘤及其微环 境通过多种途径减少肿瘤患者体DC 的数量并可抑制 DC 的成熟,从而下调或抑制其抗原提呈及免疫起始 功能,并且可以诱导调节性DC 的产生,介导免疫耐 受,最终实现免疫逃逸。故调节DC 细胞在免疫耐受 与免疫应答之间的平衡成为肿瘤疫苗治疗的一个重 要发展方向。目前国内外研究的主要方法有在体外培 养DC 的过程中加入外源性肿瘤匀浆或纯化蛋白,或 直接将肿瘤抗原导入DC,输入体内后激活CD4+T和 CD8+T细胞,发挥抗肿瘤免疫活性。在本研究中,我 们用慢病毒载体转染AFP,以及多个 AFP 抗原决定簇 肽片段处理的树突状细胞与人外周血 T细胞共培养, 诱导 AFP特异性的 T细胞免疫应答,体外在人肝癌 细胞模型上验证了抗原特异性T细胞的抗肿瘤免疫活 性。实验结果显示,AFP 特异性的 CD4+T和CD8+T 细胞对人 AFP 阳性肝癌 HepG2株细胞有显著的杀伤 活性。 抗肿瘤免疫应答反应中涉及大量细胞因子表达 的变化。我们的体外研究结果显示,AFP 特异性的 CD4+T和CD8+T细胞能显著增加细胞培养上清液中 IL-2、IFN-γ、TNF-α、穿孔素、颗粒酶的含量,证实 其抗肿瘤免疫活性与这群细胞因子表达上调密切相 关。IL-10 是已知的抗原特异性T细胞活化的抑制剂, 在HCC患者血清中可以检测到 IL-10 含量的升高;在 DC-T细胞共培养体系中加入外源性的 IL-10 中和性 抗体可以促进Th1 细胞功能。这些现象提示,降低 IL-10 水平对于抗肿瘤免疫具有十分积极的意义。在 本研究中,我们的实验结果显示,经慢病毒转染 DC 处理,或是抗原肽刺激的 DC处理后的 AFP 特异性 CD4+T和CD8+T细胞能显著降低IL-10 的含量。 目前认为杀伤性 T细胞(CTL)诱导的细胞凋亡途 径有两条分子机制:一是FasL/Fas 通路,另一个是 Perforin/Granzyme B通路。在本研究中,我们检测了 培养上清和血清中 Perforin/Granzyme B以及 FasL的 水平,实验结果显示,AFP 特异性T细胞能显著增加 Perforin/Granzyme B 的表达水平,而对 FasL 的表达水 平没有影响。因此,我们推论 AFP 特异性 CD4+T和 CD8+T细胞对肿瘤细胞的杀伤作用与 Perforin/Gran- zyme B通路有关。在本研究中,我们使用 AFP 特异 性的 CD8+T细胞单独或与 CD4+T细胞联用,处理 HepG2细胞。实验结果显示,CD4+T细胞/CD8 +T细 胞混合处理组,或是单纯 CD8+T细胞处理组均对 HepG2 细胞有较强的杀伤活性,但混合处理组的杀伤 活性更强,且体外实验结果显示,混合组有更多的IL-2 的产生,表明CD4+辅助性 T细胞可能通过促进CD8+T 细胞扩增发挥潜在的抗肿瘤免疫作用[4]。 总言之,经 DC 活化后的AFP 特异性 CD4+T和 CD8+T细胞(肝癌疫苗)具有显著的体外抗肝细胞癌免 疫活性,有望成为 AFP 阳性肝细胞癌的免疫治疗新方 法,也为今后肝细胞癌的临床治疗提供了可靠的实验 基础。 参考文献 (References) [1] L. H. Butterfield, A. Ribas, D. M. Potter and J. S. Economou. Spontaneous and vaccine induced AFP-specific T cell phenol- types in Subjects with AFP-positive hepatocellular cancer. Can- cer Immunol. Immunother, 2007, 56: 1931-1943. [2] L. H. Butterfield, A. Ribas, V. B. Dissette, et al. A phase I/II trial testing immunization of hepatocellular carcinoma patients with dendritic cells pulsed with four AFP peptides. Clinical Cancer Research, 2006, 12: 2817-2825. [3] Y. Liu, S. Daley, V. N. Evdokimova, et al. Hierarchy of alpha fetoprotein (AFP)-specific T cell responses in subjects with AFP- positive hepatocellular cancer. Journal of Immunology, 2006, 177: 712-721. [4] Y. Liu, L. H. Butterfield, X.-H. Fu, et al. Lentivirally engineered DC activate AFP-specific CD8+/CD4+ T cells whcich inhibit he- patocellular carcinoma growth in vitro and in vivo. Interna- tion- al Journal of Oncology, 2011, 39: 245-253. [5] V. N. Evdokimova, Y. Liu, D. M. Potter and L. H. Butterfield. AFP specific CD4+ helper T cell responses in healthy donors and HCC patients. Journal of Immunotherapy, 2007, 30: 425-437. [6] 刘扬, 陆崇德, 吴孟超等. HLA-A*0201 限制性 AFP 抗原决定 簇肽特异性肝癌疫苗的制备[J]. 同济大学学报(医学版), 2009, 30: 30-33. [7] A. K. Palucka, H. Ueno, J. Fay, et al. Dendritic cells: A critical player in cancer therapy? Journal of Immunology, 2008, 31: 793-805. Copyright © 2012 Hanspub 16 |