Bioprocess
Vol.
08
No.
04
(
2018
), Article ID:
27433
,
8
pages
10.12677/BP.2018.84008
A Hypothesis of Chemical Evolution of the Origin of Life
Hua Zhu
Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla Yunnan
Received: Oct. 5th, 2018; accepted: Oct. 29th, 2018; published: Nov. 6th, 2018

ABSTRACT
The microspheres constituted by proteinoids synthesized from Fox’s simulation experiments. They had peptide bond structure and weak catalysis, as well as proliferated themselves. Such microspheres were believed the models for primitive life. Due to lack of metabolism and self-reproduction, the microspheres could not meet requirements of life. Thus, how microspheres could evolve into primitive life remained unsolved mysteries. The microspheres were supposed a dissipative structure and the processes of absorption and hydrolysis could be balanced to maintain their stability by consuming proteinoid. Proteinoid molecules differed in their life span s, which were mainly determined by their multi-space structures. Consequently, molecule selection and retention could occur spontaneously in microspheres and lead to a more organized and stabilized structure of the whole microsphere with time through dissipative process. More complex chain network of chemical reactions could happen in microspheres because the proteinoid with complex, ordered multi-space structure and relatively high catalytic activity would retain. In such microspheres, nucleotides could produce and further aggregate into RNA. The synthesis of real proteins could take place with RNA as the template catalyzed by proteinoids or RNA inside microspheres. When template-based protein molecules replaced the proteinoid inside the microspheres, a protein-based self-catalyzed network of chemical reactions could take place. It is plausible if Fox’s proteinoids microspheres are to dawn on a dissipative structure, then molecule selection could occur spontaneously by “dissipative” proteinoids, and the microspheres would acquire catalytic activity due to preserving the proteinoid with a large molecular weight and relatively complex and ordered multi-space structure, and relatively high catalytic activity. Thus the microspheres would spontaneously go to self-organizing, and evolve into primitive life.
Keywords:Proteinoids, Microspheres, Dissipative Structure, Molecule Selection, Self Organization, Chemical Evolution, Origin of Life
生命起源的化学演化假设
朱 华
中国科学院西双版纳热带植物园,云南 勐腊
收稿日期:2018年10月15日;录用日期:2018年10月29日;发布日期:2018年11月6日

摘 要
在佛克斯的模拟实验上,氨基酸能发生聚合,生成一种类似蛋白质的高聚合物,这类高聚合物在水溶液中能够自动聚集成小球状的多分子体系,称之为微球体。微球体具有界膜和一定的内部结构,并能通过分裂的方式增殖,曾被认作是原始生命的雏形。模拟实验形成的类蛋白分子具有一定的多维空间结构和微弱的催化作用,但因不是模板合成,不能复制,其构成的微球体缺乏生命的基本特征—新陈代谢和自我繁殖,此模拟实验碰到了死结。生命的基本特征是新陈代谢和自我繁殖,实现这一过程是核酸与蛋白质共同作用的复杂化学反应,此复杂化学反应如何在原始地球环境出现仍是未解之谜。模拟实验中的微球体具有一个特征,即能不断地从溶液中吸收进类蛋白分子,弥补其内的类蛋白分子的水解,只要溶液中有足够的类蛋白分子的供应,微球体就能长期保存。微球体的这一特征符合耗散结构,在对类蛋白分子的“耗散”中,自发发生了由类蛋白分子自身多维空间结构的复杂有序程度相对差异所引起的留存性选择(分子选择),即分子量较大、具有相对复杂的多维空间结构的类蛋白分子较稳定及寿命较长,在自然水解中更易被保留,微球体也会因构成它的类蛋白分子相对复杂和有序,使其有序化、组织化和稳定化,会在一定程度上发生自组织。无独有偶,大分子的催化功能由其多维空间结构的状态决定,微球体通过“耗散”作用和分子选择,保留下来的正是这样的类蛋白分子。微球体可能因组成分子有相对较高的催化活性,使得在原始地球的热泉或海洋里难于自然发生的很多复杂的化学反应能在其内发生。在某些微球体内可能发生了与有机物分解相偶联的核苷酸生成的能量连锁反应,核苷酸可能进一步聚合生成核糖核酸和氨基酰–核糖核酸,并最终发生了以核糖核酸为模板,以氨基酰–核糖核酸为氨基供体,类蛋白质催化(或核糖核酸参与催化)的真正蛋白质合成,新陈代谢的基本内容形成,这样的微球体发生质变,原始生命也就诞生。微球体具有耗散结构特征,在对溶液中类蛋白分子的“耗散”过程中,发生了由类蛋白分子自身多维空间结构的复杂有序程度相对差异所引起的留存性选择,将使其有序化并发生自组织,这是微球体向原始生命演化的自发过程,并最终将导致原始生命的产生。
关键词 :类蛋白质,微球体,耗散结构,分子选择,自组织,化学演化,生命起源

Copyright © 2018 by author and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
上世纪50年代,美国学者米勒模拟原始地球条件,成功地合成了氨基酸 [1] 。氨基酸在原始地球条件下能够形成,或者陨石带来,已成为不争的事实。继米勒之后,美国学者佛克斯模拟原始地球条件,加热各种氨基酸的混合物时,氨基酸发生了聚合,生成一种类似蛋白质的高聚合物,分子量可达8000~20,000。他称之为类蛋白质,并推测这种氨基酸的热聚合反应可以在原始地球上的一些“热地区”发生。他把类蛋白质溶于水时,它们会自动聚集成一种小球状的多分子体系,他称之为微球体。微球体具有双层的界膜和一定的内部结构,可以通过出芽和分裂的方式增殖,并有某些微弱的催化作用。福克斯认为,这种微球体就是地球上生命起源的开端 [2] [3] [4] 。
米勒和福克斯等人模拟实验的成功,使生命起源之谜得以初步解开,模拟实验合成的类蛋白质,虽有肽键结构及微弱的催化作用,但不能形成蛋白质分子那样高度有序的、精密的多维空间结构。生命的特征是新陈代谢和自我繁殖。新陈代谢必须是建立在一套极复杂精密的化学反应网络基础上,它要求化学反应的高度精密和高速度进行,这是类蛋白分子所不能胜任的。同样,生物的自我繁殖是由核酸和蛋白质共同完成的。在模拟实验中核酸的合成比较困难,到目前也只能合成一些分子量不大的寡聚核苷酸。这样的话,在原始地球的“热泉”或海洋中,由类蛋白质构成的缺乏催化活性的微球体如何才能进化成原始生命并完成自我繁殖,核酸与蛋白质的联系怎样建立,仍是未解之谜。
目前生命起源仍是处于假设阶段,为科学家进一步研究探索提供思路。笔者对佛克斯模拟实验产生的类蛋白微球体怎样向原始生命进化,最终演化成原始生命,做出如下的假设。
2. 类蛋白微球体向原始生命演化的假设
佛克斯的模拟实验发现,类蛋白微球体在适当条件下,能不断地从周围溶液中吸收进新的类蛋白分子,弥补体系内不可避免的类蛋白分子的自然水解,通过消耗溶液中的类蛋白分子而生长和维持结构的稳定。只要溶液中有足够的类蛋白分子的供应,微球体结构就能长期保存 [3] 。佛克斯的模拟产生的类蛋白微球体是最有可能的原始生命形成的雏形。
普里戈金提出了耗散结构理论 [5] 。耗散结构理论认为,远离平衡态的开放系统通过不断地与外界交换物质和能量,在外界条件变化达到一定阈值时,可以通过内部的作用产生自组织现象,使系统从原来的无序状态自发地转变为时空上和功能上的宏观有序状态,形成新的、稳定的有序结构。这种非平衡态下的新的有序结构就是耗散结构(dissipative structure)。也就是说,这种远离平衡态形成的新的稳定的宏观有序结构,需要不断与外界交换物质或能量才能维持。熵值是一种在封闭的热力体系中不能做功的一定数量的热能的计量单位。热力学第二定律认为,在孤立的热力学系统中,系统的熵值总是不断增加,导致孤立的热力学系统无序度增高,熵越大,系统就会从有序进到无序的稳定状态。生命体则相反,是从无序进到有序的远平衡态的开放系统。生命体必须靠新陈代谢来维持:从环境中不断地吸取负熵,补偿熵值不可避免的增加。
回到类蛋白微球体,它们能不断地从周围溶液中吸收进新的类蛋白分子,弥补体系内不可避免的类蛋白分子的自然水解,通过消耗溶液中的类蛋白分子而生长和维持结构的稳定。这个特征正是耗散结构特征,因此,类蛋白微球体自身就是一种耗散结构。微球体并非处于平衡态,它们能不断地从周围溶液中吸收类蛋白分子,就是吃进“序”或负熵,而微球体内类蛋白分子的自然水解就是失“序”或熵增加,并能因这个过程的恒常进行,吸收与水解过程达到均衡而维持微球体结构的稳定,并且逐渐地,微球体结构还能从无序到有序,向着复杂化与组织化方向进化。
在原始地球上,类蛋白物质形成的途径和方式很多,数量也很大。并不是所有类蛋白分子的寿命都一样。类蛋白分子的寿命在很大程度上由它的多维空间结构的状态所决定。一般来说,大分子的多维空间结构越复杂精密有序,它就能形成越强的自我保护,不易被水解,分子的寿命也越长。原始地球上非模板合成的类蛋白分子,尽管多维空间结构的复杂有序程度一般较差,毕竟会有一些分子在一级结构上有相对较长的正常α-螺旋,从而在多维空间结构上就相对复杂有序,这样的分子不易水解,较稳定,寿命长。结果,在原始海洋中自发地进行着这样一种选择,那就是由类蛋白分子自身多维空间结构的复杂有序程度相对差异所引起的留存性选择。分子量较大的、具有相对较复杂有序的多维空间结构的类蛋白分子,因不易被水解而寿命相对较长,会在原始海洋中被保存积累,相反的则被水解掉,这样,类蛋白分子本身在溶液中会不断进化,亦即分子自身的留存性选择。最有意义的是在类蛋白微球体内自发进行的这种分子选择。在微球体对类蛋白分子的耗散过程中,当它吸收进分子量较大、多维空间结构复杂有序的类蛋白分子时,因它们较稳定,寿命较长,会在微球体内被保存和积累;反之则被分解消耗。通过这个耗散过程,简单的、不稳定的分子被耗散了,较复杂的、稳定的分子被保存了,使得组成微球体的类蛋白分子变得越来越复杂,越来越有序,整个微球体的结构也会相应变得越来越有序、组织化和稳定化。
大分子的催化功能与其多维空间结构的状态有关,一定催化功能的获得完全取决于一定的、高度有序的多维空间结构的获得。蛋白质分子有酶活性无非就是它有折叠的、高度精密有序的多维空间结构,在它的某些部位与底物之间形成某种契合或有种种效应。研究也证明,当RNA分子折叠形成某种多维空间结构时,它就有催化活性 [6] 。类蛋白分子也一样,分子量较大、形成了较为复杂有序的多维空间结构分子,就具有相对较高的催化活性。微球体在分子筛选中保留下来的正是这样的分子。因此,在微球体在自发地变得越来越复杂,结构越来越有序的演化过程中,它会因组成分子获得相对较高的催化活性而获得催化活性。也就是说,微球体作为一种耗散结构,在对类蛋白等物质的耗散过程中,通过自发的分子选择,实现了自组织(self organization)。自组织理论是20世纪60年代末期提出并发展起来的一种系统理论,是指一个系统在内在机制的驱动下,自发地从简单向复杂方向发展,不断地提高自身的复杂度和精细度的过程,也就是指一个系统通过与外界交换物质、能量和信息,不断地降低自身的熵含量,提高其有序度的过程。
类蛋白微球体在最初形成时是简单和缺乏催化活性的,由于其自身的耗散结构特征,通过对类蛋白分子的耗散,它们会变得越来越复杂,越来越有序,越来越具催化活性,越来越远离平衡态。类蛋白微球体的耗散结构特征,类蛋白分子的多维空间结构的状态决定其寿命的长短,以及决定其催化活性,并在微球体内自发发生了分子选择,实现了自组织,这几点的相容和一致性使得微球体向生命体的演化成为必然过程。笔者暂称自然演化形成的具有耗散结构特征,具备自组织和其构成分子具有一定催化活性的非平衡态的类蛋白微球体为“活化微球体”。在活化微球体内,化学反应的速度提高了,化学反应的方向性和有序性的程度提高了,甚至还会发生更为复杂的化学反应连锁网络。这样就创造另一个特殊微环境,使得在原始地球的热泉或海洋里难于自然发生的很多复杂的化学反应能在这样的微球体内就可能发生。
因无穷多的这样的活化微球体在原始地球上的可能存在,它们各自的组成、结构、组织化和活化的程度并不完全相同,在其内发生的化学反应的形式、内容也各有不同,从而创造了无穷多种微环境,使得各种可能的化学反应都可能在活化微球体内发生。在某些活化微球体内,发生了与有机物分解相偶联的核苷酸生成的能量连锁反应,核苷酸可能进一步聚合生成RNA,短的RNA分子能与氨基酸结合生成氨基酰-RNA,可能在一些活化微球体内最终发生了以RNA为模板,以氨基酰-RNA为氨基供体,类蛋白质催化(或许RNA也参与了催化)的真正蛋白质合成。核酸与蛋白质的联系也在此时建立,故核酸与蛋白质的联系在原始生命诞生以前就已开始。模板合成的蛋白质具有规整的肽键,正常的α-螺旋二级结构,能折叠形成高度有序的多维空间结构,因而能获得高效催化性和专一性。新合成的蛋白质不断加入到原来的化学反应网络中去,不断补充和丰富微球体内的各种化学反应,逐步替换类蛋白分子。当某些活化微球体内模板合成的蛋白质分子逐步替换了催化它们自身合成的类蛋白分子时,高效的、蛋白质合成的自催化反应网络便发生,新陈代谢的基本内容形成,活化微球体发生飞跃的质变,原始生命也就诞生(图1)。
3. 讨论
Matveev [7] 提出“生物相”中会出现特殊的物理、化学条件,有利于生物前的演化过程,生命应是起源于“生物相”。所谓“生物相”是由蛋白质与其化合的水形成,佛克斯的类蛋白微球体就是这样一种“生物相”。生物前演化的重要过程可能会在微球体内发生并在一定程度上与外部(包括海洋在内)环境隔离,微球体具有界膜与环境介质隔开,符合这个特征。研究发现钾离子介质中的肽合成速度比同等浓度钠离子介质快3到10倍 [8] ,因此,生命应起源于含钾量较高的环境,亦即有人提出的所谓钾池。Ishima et al. [9] 研究了类蛋白微球粒和介质之间的离子分配,提出微球体内的K+浓度比介质高出千倍,微球体具备向生命演化的化学条件。Matveev [7] 提出生命起源中的关键是要出现一个不平衡物理过程,即非平衡态。微球体能不断地从周围溶液中吸收进新的类蛋白分子,弥补体系内不可避免的类蛋白分子的自然水解,通过消耗溶液中的类蛋白分子而生长和维持结构的稳定。微球体具有的这种耗散结构特征使得它向生命的演化成为可能。微球体能因这个过程的恒常进行,吸收与水解过程达到均衡而维持微球体结构的稳定,并通过自发发生的由类蛋白分子自身多维空间结构的复杂有序程度相对差异所引起的留存性选择(分子选择),实现自组织。
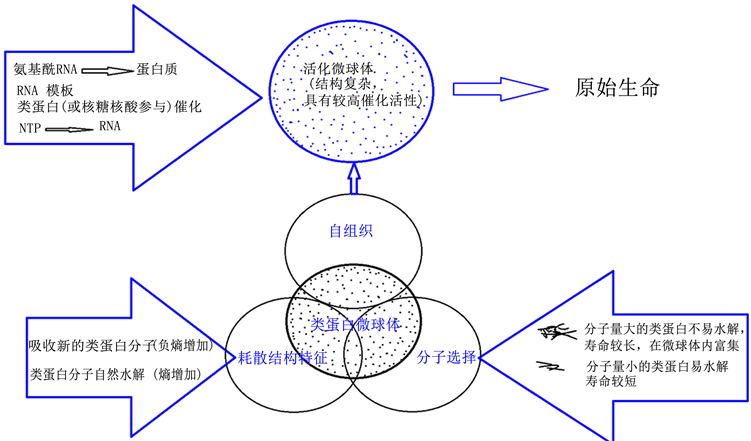
Figure 1. Diagram of proteinoid microspheres evolved into primitive life
图1. 类蛋白微球体向原始生命的演化示意图
笔者关于生命起源假设的最初思想于1986年提出 [10] ,后来于1991年发表在中国版《科学》杂志上 [11] 。该文在一些考虑和假设上类似后来多个学者发表的模拟实验和学说,例如,Huber和Wächtershäuser在模拟实验上建议了通过激活氨基酸可能生成肽 [12] 。Huber et al.进一步提出了一个可能的原始肽循环 [13] 。在笔者的假设上,由于耗散结构作用和自发的分子选择,在微球体内更为复杂的化学反应网,包括肽的生成是可能的。在生命起源上,一直有两种不同的观点,即先有自我复制或遗传体系 [14] [15] ,还是先有新城代谢 [16] 。Orgel提出了生命起源必须是等待一个比RNA简单的遗传聚合物,及一个有效的、潜在的、生命起源前的组分单体的合成途径出现。他建议了相对单纯但复杂的有机分子通过自组织,自催化循环或许会大量出现而且可用,这在原则上能解释了组分单体的起源 [14] 。在笔者提出的假设上,在一些活化微球体内可能产生了核苷酸,核苷酸可能进一步聚合形成RNA,因为在活化微球体内,化学反应的速度提高了,还会发生更为复杂的化学反应连锁网络,这与Orgel的建议是相符的。
RNA分子兼有催化和信息储存双重角色,这为反对遗传优先提供了支持 [6] 。Shapiro提出,现存的所谓复制者能作为模板复制它们自己,但最初它们从何而来仍未知道,它们一定是由无组织的混合物自发产生,这样的话,对先有新陈代谢理论应给予更多关注 [17] 。Anet也陈述了生命起源的化学反应或者是自发的,或者是被无机分子催化,或者被寡肽或类蛋白催化,这些类蛋白又可能是随机形成或相互催化形成 [16] 。De Duve阐明原始新陈代谢一定是依赖于一个适当的复杂化学过程,并建议ATP及其他NTPs在参与形成首个RNA分子之前可能作为RNA合成的先驱 [18] 。De Duve也陈述了原始新陈代谢的另一个属性就是它一定已经依赖了一套稳固的化学反应,这套化学反应能保持足够时间以致酶催化的新陈代谢出现,这个时间可能持续了几千年。在笔者的假设上,活化微球体创造了一个特殊微环境,其内许多复杂的化学反应能够发生,在某些活化微球体内,可能发生了以RNA为模板,由类蛋白质催化,或RNA共同参与催化的真正蛋白质合成。笔者的假设与Orgel、Shapiro和Anet等人的观点是相容的。佛克斯模拟实验形成的类蛋白微球体因具有耗散结构特征,使它们能保持其结构的稳定性,在其特殊微环境里,许多复杂的化学反应能够发生,在足够长的时间内,是可能发生了以RNA为模板,由类蛋白质催化,或RNA共同参与催化的真正蛋白质合成。
Carter Jr. & Wolfenden研究发现三维蛋白质结构所采用的遗传编码在不同的阶段都发生了进化,最初的进化是基于氨基酸的大小,后来是基于与水溶液中的球状折叠蛋白的兼容性,并提出通过利用tRNA的氨基接受臂来进行遗传编码,以此来确保最早的肽是具有交替的大小侧链的β结构,这种结构能够与RNA相互作用,并在此基础上编码具有更强催化活性的球状构造的蛋白质 [19] 。Wolfenden等人提出了新建议,更可能的一种情形是并非RNA催化了肽键的形成,而是肽键催化了RNA的形成,认为生命产生之前,就已经存在氨基酸和核苷酸的相互作用,后来导致蛋白质和RNA的共同产生 [20] 。这些最近的发现在一定程度上也是与我提出的假设是一致的。
Baum & Vetsigian阐述了生命起源问题的核心是解释有能力进行遗传和开放进化的化学反应系统的出现,一旦这样的系统出现了,适应性进化就可以取代。他们建议进行生命起源以前的选择实验是研究生命起源的首选,并希望来自多个学科背景的科学家将设计具体的实验以研究生命样(life-like)的化学系统,揭示自我繁殖和适应进化的证据 [21] 。我的假设也是与他们的想法相符的。
4. 结论
佛克斯通过模拟实验形成了类蛋白微球体,类蛋白质虽有肽键结构及微弱的催化作用,但这类微球体如何能进化成原始生命并完成自我繁殖,核酸与蛋白质的联系怎样建立,仍是生命起源的未解之谜。笔者提出的假设认为由微球体进化成原始生命,经过几个过程。佛克斯的模拟实验生成的类蛋白微球体能不断地从周围溶液中吸收进新的类蛋白分子,弥补体系内不可避免的类蛋白分子的自然水解,通过消耗溶液中的类蛋白分子而生长和维持结构的稳定。类蛋白微球体的这个特征正是耗散结构的特征。通过耗散过程及由类蛋白分子自身多维空间结构的复杂有序程度相对差异所引起的留存性选择(分子选择),微球体会发生自组织。因留存了分子量较大、多维空间结构复杂有序并具有一定催化功能的类蛋白分子,微球体也会逐渐获得相对较高的催化活性,使得较为复杂的化学反应网络在微球体内的发生成为可能。发生了自组织的微球体提供了一个特殊微环境,使得在原始海洋或热泉里难于自然发生的很多复杂的化学反应,如核酸的合成等,能在微球体内发生。在含有了相当量的核酸分子的微球体内,进一步发生了以核酸为模板,由类蛋白质催化或核糖核酸参与催化的真正蛋白质合成,核酸与蛋白质的联系也在此时建立。当模板合成的蛋白质逐步替换了类蛋白分子,并最终发生了由真正蛋白质催化的,以核酸为模板的蛋白质合成的自催化系统时,微球体就发生飞跃的质变,新陈代谢的化学反应网络的基本内容形成,转变成原始生命。笔者提出的这个类蛋白微球体向原始生命演化的假设,与当前涉及生命起源研究的发现基本是一致和相符的。
致谢
中国科学院西双版纳热带植物园胡彦如研究员、张帆、林华博士、刘长宁研究员提供了宝贵意见,谨此致谢!
基金项目
国家自然科学基金资助项目(41471051, 31170195, 41071040)。
文章引用
朱 华. 生命起源的化学演化假设
A Hypothesis of Chemical Evolution of the Origin of Life[J]. 生物过程, 2018, 08(04): 61-68. https://doi.org/10.12677/BP.2018.84008
参考文献
- 1. Miller, S.L. (1953) Production of Amino Acids under Possible Primitive Earth Conditions. Science, 117, 528-529. https://doi.org/10.1126/science.117.3046.528
- 2. Fox, S.W. (1965) The Origins of Prebiological Systems and of Their Mole-cular Matrices. Academic Press, New York.
- 3. Fox, S.W., Jungck, J.R. and Nakashima, T. (1974) From Proteinoid Microsphere to Contemporary Cell: Formation of Internucleotide and Peptide Bonds by Proteinoid Particles. Origins of Life, 5, 227-237. https://doi.org/10.1007/BF00927027
- 4. Nakashima, T. and Fox, S.W. (1981) Formation of Peptides from Amino Acids by Single or Multiple Additions of ATP to Suspensions of Nucleoproteinoid Microparticles. Biosystems, 14, 151-161. https://doi.org/10.1016/0303-2647(81)90064-2
- 5. Prigogine, I. and Nicolis, G. (1977) Self-Organization in Non-Equilibrium Systems. Wiley, New York.
- 6. Gilbert, W. (1986) Origin of Life: The RNA World. Nature, 319, 618. https://doi.org/10.1038/319618a0
- 7. Matveev, V.V. (2017) Comparison of Fundamental Physical Properties of the Model Cells (Protocells) and the Living Cells Reveals the Need in Protophysiology. International Journal of Astrobiology, 16, 97-104. https://doi.org/10.1017/S1473550415000476
- 8. Dubina, M.V., Vyazmin, S.Y., Boitsov, V.M., Nikolaev, E.N., Popov, I.A., Kononikhin, A.S., Eliseev, I.E. and Natochin, Y.V. (2013) Potassium Ions Are More Effective than Sodium Ions in Salt Induced Peptide Formation. Origins of Life and Evolution of Biospheres, 43, 109-117. https://doi.org/10.1007/s11084-013-9326-5
- 9. Ishima, Y., Przybylski, A.T. and Fox, S.W. (1981) Electrical Membrane Phe-nomena in Spherules from Proteinoid and Lecithin. BioSystems, 13, 243-251. https://doi.org/10.1016/0303-2647(81)90004-6
- 10. 朱华. 地球上生命的起源[M]//地质系统管理研究——交差科学与地学辩证法专辑. 成都地质学院自然辩证法研究会《地质系统管理研究》编辑部, 1986: 37-41.
- 11. 朱华. 生命起源的一种假设[J]. 科学(季刊), 1991, 43(1): 11, 52.
- 12. Huber, C. and Wächtershäuser, G. (1998) Peptides by Activation of Amino Acids with CO on (Ni,Fe) S Surfaces: Implications for the Origin of Life. Science, 281, 670-672. https://doi.org/10.1126/science.281.5377.670
- 13. Huber, C., Eisenreich, W., Hecht, S., et al. (2003) A Possible Primordial Peptide Cycle. Science, 301, 938-940. https://doi.org/10.1126/science.1086501
- 14. Orgel, L.E. (2000) Self-Organizing Biochemic Alcycles. PNAS, 97, 12507. https://doi.org/10.1073/pnas.220406697
- 15. Orgel, L.E. (2004) Prebiotic Chemistry and the Origin of the RNA World. Critical Reviews in Biochemistry and Molecular Biology, 39, 99-123. https://doi.org/10.1080/10409230490460765
- 16. Anet, F.A. (2004) The Place of Metabolism in the Origin of Life. Current Opinion in Chemical Biology, 8, 654-659.
- 17. Shapiro, R. (2000) A Replicator Was Not Involved in the Origin of Life. IUBMB Life, 49, 173-176. https://doi.org/10.1080/713803621
- 18. De Duve, C. (2003) A Research Proposal on the Origin of Life. Origins of Life and Evolution of Biospheres, 33, 559-574. https://doi.org/10.1023/A:1025760311436
- 19. Carter Jr., C.W. and Wolfenden, R. (2015) tRNA Acceptor Stem and Anticodon Bases Form Independent Codes Related to Protein Folding. Proceedings of the National Academy of Sciences of the United States of America, 112, 7489-7494. https://doi.org/10.1073/pnas.1507569112
- 20. Wolfenden, R., Lewis Jr., C.A., Yuan, Y. and Carter Jr., C.W. (2015) Temperature Dependence of Amino Acid Hydrophobicities. Proceedings of the National Academy of Sciences of the United States of America, 112, 7484-7488. https://doi.org/10.1073/pnas.1507565112
- 21. Baum, D. and Vetsigian, K. (2017) An Experimental Framework for Generating Evolvabel Chemical Systems in the Laboratory. Origins of Life and Evolution of Biospheres, 47, 481-497. https://doi.org/10.1007/s11084-016-9526-x