Botanical Research
Vol.
08
No.
02
(
2019
), Article ID:
28767
,
11
pages
10.12677/BR.2019.82015
Studies on Four Salvia Species Based on Chromosome and Pollen Morphological Comparison
Shi Feng
Shanghai Chenshan Plant Science Research Center, Chinese Academy of Sciences/Shanghai Chenshan Botanical Garden, Shanghai

Received: Jan. 12th, 2019; accepted: Jan. 25th, 2019; published: Feb. 1st, 2019

ABSTRACT
This study on four Salvia species was based on morphological comparison (S. daiguii, S. cavaleriei var. simplicifolia, S. chinensis). The first part of results showed that the chromosome number of four taxa was 16, and the basic chromosome number was x = 8. However, there were clear distinctions among the shape, size between the chromosomes of four population of Salvia. The haploid length of S. chinensis was 17.373 μm, the chromosome length of S. chinensis was ranged from 1.394 μm to 2.500 μm; the haploid length of S. cavaleriei var. simplicifolia 1 was 11.104 μm, the chromosome length of S. cavaleriei var. simplicifolia 1 was ranged from 1.092 μm to 1.723 μm; the haploid length of S. cavaleriei var. simplicifolia 2 was 14.714 μm, the chromosome length of S. cavaleriei var. simplicifolia 2 was ranged from 1.553 μm to 2.471 μm; the haploid length of S. daiguii was 17.629 μm, the chromosome length of S. daiguii was ranged from 1.514 μm to 3.028 um. The second part of results indicated that there were clear distinctions among the shape, size and exine sculpture between four populations of Salvia. The minimum of P/E was 1.118 (S. daiguii, S0418) and the maximum of P/E was 1.393 (S. cavaleriei var. simplicifolia 2, S0708). The most narrow germinal furrow was found in S. cavaleriei var. simplicifolia 2 (S0708), and the most widest germinal furrow was found in S. daiguii (S0418). There was significant difference between the protuberance of the germinal furrow of four taxa: ditch-shaped (S. daiguii, S0418), punctiform (S. cavaleriei var. simplicifolia 1, S0751), punctiform (S. chinensis, S0171), smooth (S. cavaleriei var. simplicifolia 2, S0708). This study distinguished their species taxonomy, and also provided scientific basis for the development of medical plant resources.
Keywords:Salvia, Chromosome Number, Chromosome Karyotype, Pollen Morphology, Morphological Comparison
4种鼠尾草属植物的染色体和花粉形态比较研究
冯时
中国科学院上海辰山植物科学研究中心/上海辰山植物园,上海
收稿日期:2019年1月12日;录用日期:2019年1月25日;发布日期:2019年2月1日

摘 要
本研究通过染色体和花粉的形态学特征分析,对外部形态极其相似的4个居群的药用鼠尾草属植物(华鼠尾草、张家界鼠尾草和2个血盆草居群)做了微观形态的对比分析。研究发现,4种鼠尾草的染色体数目均为2n = 2x = 16,染色体基数均为x = 8,但4个居群材料间的染色体长度和形态有明显的差异。结果显示:华鼠尾草(S0171)的染色体组总长为17.373 μm,染色体绝对长度变异范围是1.394~2.500 μm;血盆草1 (S0751)的染色体组总长为11.104 μm,染色体绝对长度变异范围是1.092~1.723 μm;血盆草2 (S0708)的染色体组总长为14.714 μm,染色体绝对长度变异范围是1.553~2.471 μm;张家界鼠尾草(S0418)的染色体组总长为17.629 μm,染色体绝对长度变异范围是1.514~3.028 μm。4个鼠尾草属植物居群的花粉大小差异不大,但形状和表面纹饰差异显著,四个不同居群的花粉长宽比最小的为1.118 (张家界鼠尾草,S0418),最大的为1.393 (血盆草2,S0708);萌发沟宽度最小的为0.48 μm (血盆草2,S0708),最大的为4.2 μm (张家界鼠尾草,S0418);每一居群沟底突起情况均不同:沟壑状突起(张家界鼠尾草,S0418),点状突起(血盆草1,S0751),点状突起(华鼠尾草,S0171),无突起(血盆草2,S0708)。本研究同时探讨了鼠尾草属植物花粉的多样性。为鼠尾草属植物的形态学研究积累了一定资料,同时可作为上述几种鼠尾草属植物物种间鉴定与系统分类的较可靠依据。
关键词 :鼠尾草,染色体数目,核型分析,花粉形态,形态学比较

Copyright © 2019 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
鼠尾草属(Salvia)是唇形科(Lamiaceae)最大的一个属,有近1000种 [1] ,分布世界各地。中国原产鼠尾草82种,以西南横断山区物种多样性最为丰富 [2] 。鼠尾草属具有较高的利用价值,在医药、食品、化妆品、景观园艺等领域有着广泛地应用。欧美国家对鼠尾草的资源利用较早较充分,原产自欧洲的鼠尾草(S. officinalis, sage, garden sage)是几种草药、食品、化妆品的常用成分或添加剂,用于治疗咽喉炎,也是厨房中常备的调味品,被列入美国食品药品管理局(FDA)和欧洲药典(6th ed.)的安全药物名录。在园艺领域,鼠尾草的观赏价值也获得广泛利用和认可,原产美洲的一串红(S. splendens)很早就被引入我国,在大型活动的主题花境和家庭园艺中常常能看到其身影。
现代植物化学和药理学研究表明,鼠尾草属植物主要含有两大类活性成分,水溶性的酚酸类和脂溶性的二萜酮类。以丹参(S. miltiorrhiza)为例,主要具有抗氧化作用,对心脑血管动脉硬化、心肌梗塞、降低心肌耗氧量、抗凝血及血栓形成以及内脏微循环等具有治疗和改善作用。在丹参之外,我国各地还普遍利用鼠尾草属的多种植物作为丹参替代品或治疗其他疾病。如甘西鼠尾草(S. przewalskii)、南丹参(S. bowleyana)和云南鼠尾草(S. yunnanensis)常作为西北、华南和西南地区的丹参替代品;而荔枝草(S. plebeia)在抗菌消炎、华鼠尾草(S. chinensis)在消肿散结、三叶鼠尾草(S. trijuga)在活血调经、红根草(S. prionitis)在抗肿瘤方面的作用则比较突出 [3] [4] 。
鼠尾草属植物的一般形态学特征是:草本或半灌木或灌木。叶为单叶或羽状复叶。轮伞花序2至多花,组成顶生或腋生的假总状花序。花萼筒形或钟形,二唇形。花冠筒内藏或外伸,平伸或向上弯或腹部增大。能育雄蕊2,生于冠筒喉部的前方,花丝短,水平生出或竖立,药隔延长,线形,横架于花丝顶端,以关节相联结,成丁字形。药隔上臂顶端着生椭圆形或线形有花粉的药室,下臂或粗或细,顶端着生有花粉或无花粉的药室或无药室,二下臂分离或联合。退化雄蕊2,生于冠筒喉部的后边,呈棍棒状或小点,或不存在。子房4全裂,小坚果卵状三形或长圆状三棱形,无毛,光滑。该属植物以异花授粉为主,遗传变异较大,亲缘关系复杂,属内的分类系统和资源现状较为混乱,对鼠尾草属植物种间亲缘关系的研究也鲜见报道。
本研究的4种材料为2个血盆草(Salvia cavaleriei var. simplicifolia)居群、华鼠尾草(S. chinensis)以及张家界鼠尾草(S. daiguii),其中血盆草和华鼠尾草是地方习用的药用植物,血盆草两个居群的分布生境和形态学差异较明显,因此本研究为进一步厘清这几种药用鼠尾草的系统关系以及其药用价值奠定基础。
2. 染色体和花粉形态在分类中的意义
根据传统手段进行分类有时候会将形态相近的同属植物认定为同一个种。当传统分类学无法准确地对宏观形态相近物种进行区分时,我们可以从植物细胞的微观结构来一探究竟。染色体数目、核型分析和花粉形态观察是常见的植物微观分类手段。
2.1. 光镜下染色体的比较
由于任何一种生物的染色体核型(chromosome karyotype)是相当固定的,它的染色体数目(chromosome number)、染色体基数(chromosome basic number)、染色体倍性(chromosome ploidy)、染色体组总长度(haploid chromosome length, HCL)、染色体长度的变异范围、染色体绝对长度(chromosome Length, CL)、染色体相对长度(relative length %, RL%)、最长染色体与最短染色体的比值(L/S)、着丝点位置、最长臂与最短臂的比值(臂比)变异范围、不对称性、核型类型等,在种内具有非常稳定的模式(即核型) [5] - [11] 。因此,本研究选取4 种鼠尾草属植物进行根尖染色体压片,观察其染色体数目和形态。可为鼠尾草属植物的分类和药用资源的应用提供参考。
根据国内外多项研究成果证实,自然状态下,鼠尾草属植物多为二倍体物种,且鼠尾草属植物的染色体数目在属内具有多样性。鼠尾草属植物染色体数目多为12,14,16,22,24,44,染色体基数(x)多为7,8,9 [11] [12] 。鼠尾草属植物的染色体很小,一般属于微小型染色体或小型染色体,难以用于核型分析 [5] 。最新的研究成果认为,中国所有的鼠尾草属植物的染色体基数可能都是x = 8 [13] [14] 。
2.2. 电镜下花粉形态的比较
植物的花粉主要由遗传因素决定,受环境因素的影响较小,其成熟花粉粒的形态结构特征十分稳定,大量花粉资料显示,花粉形态是进行被子植物系统分类的重要依据 [15] - [31] 。在被子植物系统研究中,孢粉学能在短时间内,利用很少资料获得大量信息。
国内对鼠尾草属植物的花粉研究较少,除王开发 [22] 对20种鼠尾草属植物花粉的电镜显微图像进行了比较系统的分析外,大多国内研究者仅对丹参或可做丹参替代品的雪山鼠尾草(S. evanaiana)、云南鼠尾草(S. yunnanensis)、黄花鼠尾草(S. flava)等进行对比 [13] - [18] ,而对张家界鼠尾草(S. daiguii)的花粉形态学研究未见报道。杨德奎等 [17] 利用扫描电子显微镜首次对山东鼠尾草属(Salvia L.) 3种植物的花粉形态进行了观察和研究。研究结果表明,鼠尾草属花粉的萌发孔均为6沟,外壁表面均具网状雕纹,这是该属的共同特征。但从花粉的形状可分为2个类型,即长球形(如荔枝草和丹参)和扁球形(如一串红),表现出了种间差异。从花粉粒大小和网状纹饰的深浅、形状等的细微结构分析,差异也较明显。因此,花粉的形状、大小、外壁雕纹结构特征,可作为种间分类的参考依据 [17] 。另外,YL Zhang等 [32] 认为花粉粒大小可能与植物生长环境有关,花粉形态要比花粉大小更加稳定。
本文使用扫描电镜分别对4种鼠尾草属植物的花粉粒形态及其表面雕纹进行了观察研究,旨在为该属植物的分类提供花粉形态学方面的依据,也为唇形科植物花粉粒形态的研究积累资料。
3. 实验
3.1. 染色体数目与形态研究
3.1.1. 实验材料
用于染色体研究的鼠尾草属植物采集信息,如表1所示。

Table 1. Collection data of the Salvia taxa examined in this study
表1. 用于染色体研究的鼠尾草属植物采集信息
实验器材:
OLYMPUS DP73成像系统,OLYMPUS BX43细胞学显微镜,西门子KK25V61T1冰箱,一恒TU-100 恒温金属浴,0℃~100℃温度计,2 mL离心管,1.5 mL离心管,离心管架,培养皿,0.002 mol/L 8-羟基喹啉溶液,卡诺氏固定液(无水乙醇:冰醋酸 = 3:1),1 mol/L盐酸,卡宝品红染色液,胶头滴管,载玻片,盖玻片,镊子,解剖剪,解剖刀,解剖针,滤纸,铅笔,竹签,中性记号笔。
3.1.2. 实验方法
本次的方法参照胡忠红等 [10] 所使用的方法,经过预实验做了一些调整,具体如下:
1) 材料的预处理:于上午9~11点取生长旺盛的鼠尾草属植物幼嫩根尖0.5~1.0 cm,取材后用清水和软毛刷清理沙土。待根尖清理干净后,尽快放入盛有8-羟基喹啉溶液的2 mL离心管中(此操作应在通风橱进行),在15℃~20℃下预处理6至8小时。其间要注意温度的控制。
2) 固定:将预处理后的根尖直接放入盛有卡诺氏固定液的2 mL离心管中,放置在4℃冰箱10小时以上(或至少过夜)。固定后若未立即使用根尖,可继续在4℃冰箱中存放数星期。
3) 漂洗与解离:从卡诺氏固定液中取出固定好的根尖,在清水中漂洗约10 min;再将根尖放入盛有60℃、1 mol的盐酸的1.5 mL离心管中,解离10~12 min (不同种时间不同)。解离后立即取出根尖放入装有清水的培养皿漂洗,更换至少4次水,其间可用装有蒸馏水的洗瓶等用具吹打冲洗,增强漂洗效果。将漂洗后的根尖放在装有蒸馏水的培养皿中备用。
4) 制片与观察:取出待用根尖置于载玻片上,用滤纸吸去根尖周围大部分的水(不要彻底吸干,以防在操作过程中根尖失水过多,影响细胞形态),用刀或镊子将根尖分生区切下,用解剖针挑去根冠和表皮组织,留下分生区细胞。再次用滤纸吸去残留水分,之后在材料上滴一滴卡宝品红染色液,染色5 min后,盖上盖玻片,注意尽量避免产生气泡,若有气泡产生,可轻压盖玻片挤去气泡,并保持盖玻片不滑动,用滤纸吸去多余染液。用竹签敲击盖玻片,力度均匀适中,敲击方向垂直于盖玻片,施力点密集,使细胞尽可能均匀分散为一层。用OLYMPUS BX43研究级显微镜观察目标区域内的分生区细胞,寻找分裂期的染色体。若看到有染色体未分散在同一平面上的情况,应重复敲击盖玻片的步骤,直至染色体均匀分散。选取染色体分布理想的视野,用OLYMPUS DP73成像系统进行拍照,选取对应的比例尺印入照片,并保存图像。将符合要求的临时装片用加拿大中性树胶进行封片,制成永久装片,标注标本编号和制作日期。
3.2. 花粉形态学比较研究
3.2.1. 实验材料
FEI Quanta 250扫描电子显微镜(以下简称“扫描电镜”),Magnetron Ion Sputter Metal Coating Device MSP-2S离子溅射仪,样品台,日新NEM双面导电胶带(SEM专用),硫酸纸,4种鼠尾草花粉,PCR管或离心管(用于存放、晾干花粉)。
3.2.2. 实验方法
1) 用镊子取盛花期发育成熟、形态饱满的鼠尾草花粉,置于pcr管或离心管内自然风干3~4天(与环境湿度有关),周围可放变色硅胶保持合适的湿度,其间可用光学显微镜观察花粉干燥情况。
2) 剪取适当大小的导电胶带,将其粘在样品台上,再轻轻沾取微量烘干的花粉,尽量使得不同花粉在胶带上分散开,避免重叠,以便于观察与拍照,由于花粉粒较小,该步骤可借助光学显微镜操作。若花粉重叠,可用洗耳球轻轻吹开花粉,但注意在操作期间避免不同种的花粉之间互相混杂造成材料污染。粘好花粉后编号,不同编号的花粉分开存放于密封箱内。
3) 将粘好花粉的样品托与样品托架放入离子溅射仪中镀铂金膜。
4) 操作扫描电镜:A:放气3分钟;B:将样品台放入扫描电镜的样品室,执行抽真空20分钟;C:开灯丝(beam on),升样品台(按住鼠标滑轮上升至黄线1 cm处);D:自动白平衡,自动对焦,找到目标物;E:手动调节对比度、亮度、对焦,找到理想的视野,拍照。
4. 结果与讨论
4.1. 染色体研究
4.1.1. 染色体数目

Figure 1. Photographs of somatic metaphase chromosome of 4 Salvia taxa from China: (A) S. daiguii (S0418); (B) S. cavaleriei var. simplicifolia 1 (S0751); (C) S. cavaleriei var. simplicifolia 2 (S0708); (D) S. chinensis (S0171). Scale bar = 5 μm. (zooming in 100 times)
图1. (A) 张家界鼠尾草(S0418);(B) 血盆草1 (S0751);(C) 血盆草2 (S0708);(D) 华鼠尾草(S0171)。标尺长度 = 5 μm。(放大100倍)
结果显示:以上四个居群的鼠尾草属植物均为二倍体,染色体数目均为16,染色体基数x均为8。染色体数目在同一物种中具有高度的保守性,染色体数目、单倍体染色体总长度(HCL)的差异与物种之间亲缘关系的远近具有一致性,因此由染色体数目相同可推测本研究所选择的四个居群在遗传上具有较高的保守性,这也正是它们在形态上容易混淆,但彼此确实又存在一定差异的证据。染色体总长度和形态的差异又决定了彼此间的性状差异,从图1中可以明显看出,四个居群染色体的大小和形态有较明显差异,如染色体绝对长度、形态都有等都有明显差异(见表2)。
4.1.2. 染色体核型分析
Table 2. Main characteristics of the chromosome morphology in four accessions of Salvia
表2. 4种鼠尾草属植物材料染色体形态学特征
注:HCL:染色体组总长度,CL:染色体绝对长度,RL%:染色体相对长度,L/S:最长与最短染色体长度比值。
张家界鼠尾草(S0418)核型公式K(2n) = 2x = 16。
血盆草1 (S0751)核型公式K(2n) = 2x = 16。
血盆草2 (S0708)核型公式K(2n) = 2x = 16。
华鼠尾草(S0171)核型公式K(2n) = 2x = 16。
表2比较了的四种鼠尾草属植物的染色体组总长(HCL)、染色体绝对长度(CL)、染色体相对长度(RL%)、最长、最短染色体长度比(L/S)。结果显示,染色体组长度由长到短分别为张家界鼠尾草(S0418)、血盆草1 (S0751)、血盆草2 (S0708)、华鼠尾草(S0171)。张家界鼠尾草(S0418)的染色体组总长为17.629 μm,染色体绝对长度变异范围:1.514~3.028 μm;血盆草1 (S0751)的染色体组总长为11.104 μm,染色体绝对长度变异范围:1.092~1.723 μm;血盆草2 (S0708)的染色体组总长为14.714 μm,染色体绝对长度变异范围:1.553~2.471 μm;华鼠尾草(S0171)的染色体组总长为17.373 μm,染色体绝对长度变异范围:1.394~2.500 μm。
通过结果比较可以发现:两种血盆草的染色体组总长、染色体绝对长度变异范围和最长、最短染色体长度比值差异很大,染色体相对长度也有较明显的差异。该结果说明两种分布生境和形态学差异较明显的血盆草,在遗传上也有较大差异,亲缘关系较远,可能是两种不同分类群。形态上与张家界鼠尾草(S0418)极为相似的血盆草2 (S0708),其染色体组长度也与张家界鼠尾草(S0418)有一定差异,但比血盆草1 (S0751)更为接近,这也很可能是与张家界鼠尾草(S0418)和血盆草2 (S0708)形态相似的原因之一。
4.2. 花粉形态比较研究
根据王开发等 [22] 对20种鼠尾草属植物的研究可知,在扫描电镜下,当花粉外壁放大至5000倍以上时,虽然都是网状纹饰,但显示出差异性。从以往研究的鼠尾草属植物花粉的网状纹饰中,大致可以有下几种类型:
Table 3. Seven sculpture types of pollen grains of Salvia
表3. 7类鼠尾草属植物花粉纹饰类型
花粉粒是种子植物雄蕊花粉囊内的粉状体,属雄配子体,遗传物质的携带者,在植物系统发育中比其他性状更为稳定 [20] 。
本研究对4种花粉的扫描电镜图像进行对比,将四种花粉的形态特征进行列表(见表4)。对该表的每一项进行对照分析,结果显示:4种鼠尾草属植物的花粉大小、形状和外壁网状纹饰等都有差异。其中花粉长宽比最小的为1.118 (张家界鼠尾草,S0418),大多呈近似圆球形;最大的为1.393 (血盆草2,S0708),大多呈饼状或长球形。萌发沟宽度最小的为0.48 μm (血盆草2,S0708),最大的为4.2 μm (张家界鼠尾草,S0418);4个居群鼠尾草属植物的沟底突起情况均不同:沟壑状突起(张家界鼠尾草,S0418),点状突起与块疣状突起(血盆草1,S0751),无突起(华鼠尾草,S0171),点状突起(血盆草2)。综合本研究4种鼠尾草的形态特征,可将它们大致归为表3中7种常见鼠尾草纹饰类型中的3类:上下两层网组成的网状纹饰(张家界鼠尾草,S0418)和(血盆草2,S0708);不规则网状纹饰(血盆草1,S0751);浅网状纹饰(华鼠尾草,S0171)。其中华鼠尾草(S0171)花粉的结构描述与电镜图像与王开发等 [20] 。
有一定差异,这种差异有可能是花粉发育时期不同导致的,有待进一步研究。本研究为鼠尾草属植物的花粉形态学研究积累了一定资料,同时可作为上述几种鼠尾草属植物物种间鉴定的可靠依据。
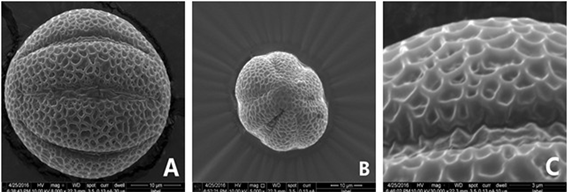



Figure 2. SEM micrographs of pollen grains in the Salvia: (A)~(C) S. daiguii (S0418); (D)~(F) S. cavaleriei var. simplicifolia1 (S0751); (G)~(I) S. cavaleriei var. simplicifolia 2 (S0708); (J)~(L) S. chinensis (S0171)
图2. (A)~(C) 张家界鼠尾草(S0418);(D)~(F)血盆草1 (S0751);(G)~(I)血盆草2 (S0708);(J)~(L)华鼠尾草(S0171)花粉扫描电镜图
图2中,从左至右三列分别为四种鼠尾草属植物花粉的赤道观、极观、高放大倍数下的网状结构。
Table 4. Main characteristics of the pollen morphology in four accessions of Salvia
表4. 4种鼠尾草属植物材料花粉形态学特征
4.3. 综合两个形态学研究的推测
本研究对四种鼠尾草属植物的染色体形态、核型与花粉形态进行了详细的对比。结合两个实验的结果可以看出,张家界鼠尾草和血盆草1 (S0418, S0751),其染色体核型与花粉形态都具有较大区别。而产于贵州的血盆草2 (S0708),其染色体核型与花粉形态却与张家界鼠尾草(S0418)更为接近,这让笔者有理由做如下推测:1) 根据《中国植物志》检索,编号(S00751)与(S0708)的两个鼠尾草属植物居群,其检索结果都为血盆草(S. cavaleriei var. simplicifolia) [2] 。但从染色体和花粉的比较中可以发现,这两个居群的遗传差异是很显著的,可作为今后血盆草物种种下鉴定的依据;2) 张家界鼠尾草(S0418)与血盆草2 (S0708),它们的染色体核型比较接近,花粉形态也有很多相似之处,体现出了其亲缘关系可能较近,可进一步通过聚类分析等其他分析手段来探讨它们的亲缘关系。
4.4. 鼠尾草属植物花粉的“多形性”
鼠尾草属植物的花粉具有多形性,多形性指的是同一种鼠尾草属植物的花粉具有2种或2种以上形状。本研究中,2种鼠尾草属植物(S0418,张家界鼠尾草;S0708,血盆草2)的花粉均具有2种以上的形状(花粉粒整体形状),这与李晓娟 [17] [18] 提到的花粉“二形性”相似。花粉不同形状的形成既与遗传物质有关,也与花粉发育时期有关 [20] 。而花粉的极轴长与赤道轴长的比值与花粉形状最为密切相关,这是微观分类上很重要的划分依据。同时,同一物种中花粉的“多形性”也提醒我们,不能仅凭借花粉的大小和形状来确定不同鼠尾草属植物的种间差异,应当结合花粉外壁的纹饰来作比较,尤其是通过丰富的网状结构与不同形态、不同密度的突起对鼠尾草属植物的花粉进行有效的分类。
5. 结论
鼠尾草属植物染色体和花粉形态研究在国内外都鲜有报道。本研究通过细胞学和孢粉学两种微观分类手段对4个鼠尾草属植物居群进行了比较,研究结果显示出4个鼠尾草属居群种中,无论是亲缘关系较远的2个血盆草(S. cavaleriei var. simplicifolia)居群与华鼠尾草居群(S. chinensis),还是形态上极其相似、容易混淆的张家界鼠尾草(S. daiguii)居群与血盆草2 (S. cavaleriei var. simplicifolia)居群,或是目前可被《中国植物志》归为同一类的血盆草1 (S. cavaleriei var. simplicifolia)和血盆草2 (S. cavaleriei var. simplicifolia),它们之间的微观形态差异显著。研究结果充分显示了这两种微观分类手段可作为鼠尾草属植物分类的可靠微观分类依据,也对药用鼠尾草的应用具有重要指导意义,同时为鼠尾草属植物原有物种的鉴定和修订作铺垫。
基金项目
本研究由1、中国科学院战略生物资源科技支撑体系运行专项(CZBZX-1);2、上海市绿化和市容局辰山专项课题(G172410, G182409)资助。
文章引用
冯 时. 4种鼠尾草属植物的染色体和花粉形态比较研究
Studies on Four Salvia Species Based on Chromosome and Pollen Morphological Comparison[J]. 植物学研究, 2019, 08(02): 107-117. https://doi.org/10.12677/BR.2019.82015
参考文献
- 1. Wester, P. and Claßen-Bockhoff, R. (2001) Pollination Syndromes of New World Salvia Species with Special Reference to Bird Pollination. Annals of the Missouri Botanical Garden, 98, 101-155. https://doi.org/10.3417/2007035
- 2. 吴征镒等. 中国植物志[M]. 北京: 科学出版社, 1977: 145-150.
- 3. 国家药典委员会. 中华人民共和国药典[M]. 北京: 化学工业出版社, 2005.
- 4. 肖培根. 新编中药志[M]. 北京: 化学工业出版社, 2002.
- 5. 赵红霞. 丹参、雪山鼠尾、云南鼠尾的显微鉴定学、细胞学及药用成分、无机元素含量研究[D]: [硕士学位论文]. 雅安: 四川农业大学, 2007.
- 6. 张德山, 赵建萍. 荔枝草的染色体数目和核型分析[J]. 山东师大学报(自然科学版), 1991, 6(3): 109-111.
- 7. 赵红霞, 张利, 凡星, 杨瑞武, 丁春邦, 周永红. 丹参、黄花鼠尾和雪山鼠尾染色体数目的研究[J]. 中国中药杂志, 2006, 31(22): 1847-1849.
- 8. 朱琨, 张璐, 陈雅君, 张楚涵, 周阳, 李雪. 37个早熟禾品种的花粉形态及其分类学意义[J]. 草地学报, 2011, 19(1): 114-121.
- 9. 张彦妮, 钱灿. 12种百合属植物花粉形态扫描电镜观察[J]. 草业学报, 2011, 20(5): 111-118.
- 10. 胡忠红, 欧鑫, 谭昕, 易自力, 蒋建雄, 陈智勇. 3 种鼠尾草属植物的核型分析[J]. 中国农学通报, 2011, 27(27): 250-254.
- 11. 杨在君. 四川和重庆的鼠尾草属植物资源调查、细胞学及无机元素研究[D]: [硕士学位论文]. 雅安: 四川农业大学, 2008.
- 12. Gill, L.S. (1971) Chromosome Studies in Salvia (Labiatae): West-Himalayan Species. Cellular and Molecular Life Sciences, 27, 596-598. https://doi.org/10.1007/BF02147623
- 13. Hu, G.X., Xiang, C.L., Liu, E.D., et al. (2016) Karyotypic Study of Eighteen Taxa of Salvia (Lamiaceae) from China. Caryologia, 69, 50-57. https://doi.org/10.1080/00087114.2015.1109955
- 14. Wang, M., Zhang, L., Ding, C.B., et al. (2009) Meiotic Observations of Eight Taxa in the Genus Salvia. Caryologia, 62, 334-340.
- 15. 曹丽敏. 鼠尾草属植物研究进展[J]. 湖南农业科学, 2009, 1(3): 137-139, 147.
- 16. 杨向晖, 吴颖欣, 林顺权. 6种枇杷属植物花粉形态扫描电镜观察[J]. 果树学报, 2009, 26(4): 572-576.
- 17. 胡彦, 丁友芳, 温春秀, 谢晓亮, 刘玉军. 唇形科三个属植物花粉粒形态的比较研究[J]. 北方园艺, 2012, 1(11): 60-64.
- 18. 刘文婷. 丹参的生物学特性研究[D]: [硕士学位论文]. 杨凌: 西北农林科技大学, 2004.
- 19. 李晓娟, 李建秀, 孙经兴, 宋英杰, 张永清. 扫描电镜下山东丹参与近缘种花粉形态特征比较观察[J]. 广西植物, 2013, 33(1): 20-24.
- 20. 李晓娟. 山东丹参与其近缘种的分类鉴定研究[D]: [硕士学位论文]. 济南: 山东中医药大学, 2013.
- 21. 杨德奎, 孙京田, 王丙全. 山东鼠尾草属花粉形态的研究及其在分类上的意义[J]. 山东科学, 2003, 16(1): 14-16.
- 22. 王开发, 黄秀兰. 我国鼠尾草属花粉形态研究[J]. 植物研究, 1986, 6(4): 55-71.
- 23. 蔡秀珍, 刘克明, 丛义艳, 陈薇. 凤仙花属(Impatiens L.)10种植物花粉形态的扫描电镜观察[J]. 植物研究, 2007, 27(3): 279-283.
- 24. 徐柏森, 杨静. 植物花粉扫描电镜技术研究与观察[J]. 西南林学院学报, 2007, 27(5): 21-24.
- 25. 贾子瑞, 王军辉, 张守攻, 马建伟, 杨桂娟. 云杉属花粉形态的电镜扫描研究[J]. 林业科学, 2014, 50(5): 49-61.
- 26. 刘鹏, 马利超, 王宇, 刘志鹏. 野豌豆属牧草种质花粉形态的扫描电镜观察[J]. 草业科学, 2013, 30(11): 1755-1761.
- 27. 韩柏明, 赵密珍, 王静, 于红梅. 草莓属及其近缘属植物的花粉形态电镜观察[J]. 东北农业大学学报, 2013, 44(10): 104-110.
- 28. 王延秀, 陈佰鸿, 王淑华, 胡紫璟, 党兆霞, 魏晓燕. 11个观赏海棠品种花粉形态扫描电镜观察[J]. 植物研究, 2014, 34(6): 751-757.
- 29. 刘鹏, 马利超, 王宇, 刘志鹏. 16 种野豌豆属植物花粉形态的扫描电镜观察[J]. 草业学报, 2015, 24(1): 107-114.
- 30. 刘鹏. 野豌豆属牧草的遗传多样性研究[D]: [硕士学位论文]. 兰州: 兰州大学, 2015.
- 31. 王亚敏, 汪洪捷, 张卓勇. 松花粉的红外光谱、扫描电镜和X射线能谱仪分析[J]. 光谱学与光谱分析, 2005, 25(11): 55-58.
- 32. 王湘南, 陈永忠, 蒋丽娟, 刘志玲, 彭邵锋, 王瑞, 马力, 杨小胡. 油茶优良无性系花粉形态结构的扫描电镜观测[J]. 中南林业科技大学学报, 2010, 30(5): 67-71.