Advances in Applied Mathematics
Vol.
09
No.
02
(
2020
), Article ID:
34092
,
9
pages
10.12677/AAM.2020.92016
Hopf Bifurcation Control of DCN Pyramidal Neurons
Zihan Wang, Shuai Wu
College of Mathematics and Information Science, Guangxi University, Nanning Guangxi

Received: Jan. 9th, 2020; accepted: Jan. 27th, 2020; published: Feb. 3rd, 2020

ABSTRACT
Pyramidal cell in the dorsal cochlear nucleus (DCN) is one of the first neurons in the central auditory system to process acoustic signals, which plays an important role in the processing and integration of acoustic information. The KM model based on voltage dependent ion channel variables can be established by the in vitro experimental data of DCN pyramidal neurons. We use hf as a bifurcation parameter to analyze the fast and slow dynamics of DCN pyramidal neurons. Then, a dynamic state-feedback control law incorporating a washout filter is used to control the Hopf bifurcation position of KM model, and our control task is realized.
Keywords:KM Model, Hopf Bifurcation, Washout Filter, Bifurcation Control

DCN椎体神经元的Hopf分岔控制
王子汉,伍帅
广西大学数学与信息科学学院,广西 南宁

收稿日期:2020年1月9日;录用日期:2020年1月27日;发布日期:2020年2月3日

摘 要
耳蜗背侧核(DCN)椎体神经元是听觉中枢系统中首先对声波电信号进行处理的神经元之一,其对声波信息的处理、整合起了非常重要的作用。通过DCN椎体神经元的离体实验数据可以建立基于电压依赖型离子通道变量的数学模型KM模型。我们将hf作为分岔参数对DCN椎体神经元模型进行快慢动力学分析。然后使用冲洗滤波器辅助动态反馈控制器的方法来控制KM模型的Hopf分岔位置,实现我们的控制任务。
关键词 :KM模型,Hopf分岔,冲洗滤波器,分岔控制

Copyright © 2020 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
众所周知,神经系统是复杂的非线性动力系统。从非线性动力系统的角度来看,许多精神疾病可以被认为是由描述神经元系统的相关非线性方程中一个或多个调节参数值的变化引起的分岔所引起的 [1]。因此,理解导致神经元系统分岔的机制和可以修改这种分岔特征的反馈控制法,对神经疾病的新诊断和治疗有着重要意义。
局部分岔分析是非线性系统理论的一个重要分支,非线性系统平衡点的性质因单参数改变而改变 [2]。其中最常见的分岔类型之一是Hopf分岔,平衡点由于参数改变引起的一对纯复特征值而失去稳定性。Chunli Huang等 [3] 利用由线性项和非线性立方项组成的冲洗滤波辅助动态反馈控制器来控制Hopf分岔的发生。Yong Xie等 [4] 运用冲洗滤波辅助动态反馈控制器将不变圆(snic)分岔改变为Hopf分岔,模型神经元的兴奋性由I型兴奋性转变为II型兴奋性。使用冲洗滤波器的主要优点是它保留了原始平衡点。但是,由于一个Hopf分岔点的重新定位会影响另一个Hopf分岔点的位置,因此滤波器不能应用于具有两个Hopf分岔的系统。Le Hoa Nguyen等 [5] 利用动态状态反馈控制,将具有II型兴奋性的ML神经元模型中的两个Hopf分岔点同时重新定位到新的期望位置。动态状态反馈控制律的方法不仅能够将两个不同的Hopf分岔点同时重新定位到n维非线性系统中的任何所需位置,而且能够保持系统的平衡结构。
冲洗滤波辅助动态反馈控制器方法在高维系统的分岔控制上有一定难度,本文通过对一个9维的神经元模型进行分析,运用冲洗滤波辅助动态反馈控制器方法对系统的Hopf分岔进行了控制。本篇文章的结构如下。在第2节介绍了基于耳蜗背侧核(DCN)椎体神经元建立的9维数学模型KM模型。第3节我们引入快慢动力学分析,通过膜电位V的时间历程图分析我们将系统的快钾电流的失活变量 作为快子系统的分岔参数,得到模型的分岔图。第4节引入冲洗滤波器的动态状态反馈控制律对两个模型的分岔进行控制。在第5节中给出结论。
2. Kanold-Manis DCN椎体神经元模型(KM模型)
基于H-H模型的思想,2001年P. Kanold和P. Manis根据大鼠的耳蜗背侧椎体神经元的体外膜片钳记录结果建立了一个9维的基于电压依赖型离子通道变量的数学模型 [6],在这里我们称之为KM模型。在KM模型中包含钠电流(
),漏电流(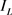 ),h电流(
),h电流(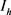 )和3种类型钾电流,这三种钾电流分别是快钾电流(
)和3种类型钾电流,这三种钾电流分别是快钾电流(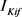 ),慢钾电流(
),慢钾电流(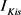 )和永不失活的钾电流(
)和永不失活的钾电流(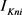 )。
)。 与
与 的主要区别在于
的主要区别在于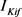 的失活通道变量的时间尺度(大约50 ms)快于
的失活通道变量的时间尺度(大约50 ms)快于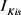 的失活变量时间尺度(大约200 ms)。
的失活变量时间尺度(大约200 ms)。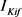 的失活变量对DCN神经元的放电模型调控上起来决定性作用。方程如下:
的失活变量对DCN神经元的放电模型调控上起来决定性作用。方程如下:
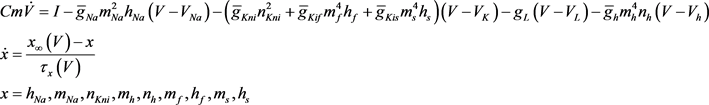
电压依赖的通道变量的稳态方程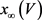 的一般形式是
的一般形式是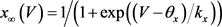 。电压依赖的通道变量的弛豫时间为
。电压依赖的通道变量的弛豫时间为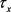 。
。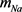 ,
,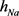 表示钠离子通道的开关概率;
表示钠离子通道的开关概率;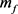 ,
,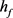 和
和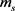 ,
, 分别表示快钾离子通道和慢钾离子通道的开关概率;
分别表示快钾离子通道和慢钾离子通道的开关概率;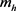 ,
, 表示h电流的开关概率;
表示h电流的开关概率; 为永不失活钾离子通道的打开概率。
为永不失活钾离子通道的打开概率。
DCN椎体神经元模型的具体通道开关概率参数如下: ,
, ,
,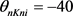 ,
, ,
,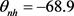 ,
,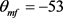 ,
,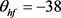 ,
, ,
, ,
, ,
,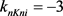 ,
,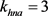 ,
, ,
,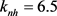 ,
, ,
,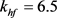 ,
,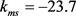 ,
, 。膜电容
。膜电容 。最大通道电导常数(单位:nS)为:
。最大通道电导常数(单位:nS)为: ,
,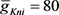 ,
,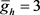 ,
,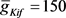 ,
, ,
,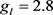 。各离子电流的可逆电位(单位:mV)为:
。各离子电流的可逆电位(单位:mV)为: ,
, ,
,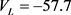 ,
, 。
。
3. KM模型的分岔分析
3.1. 快慢动力学分析
一般来说,复杂的神经系统模型大都包含两个或两个以上的时间尺度。在实验和数值模拟中经常观察到得簇放电行为通常是在慢尺度调控下的动力系统行为。考虑多时间尺度系统

 向量
向量 表示对应于放电形式的快变量。它包含膜电位V以及快电流的激活或非激活的门控变量,等等。向量
表示对应于放电形式的快变量。它包含膜电位V以及快电流的激活或非激活的门控变量,等等。向量 表示调制快放电的相关的慢变量,比如,慢钾电流的门控变量、内钙离子浓度等。小参数表示快慢时间尺度的比率,也即放电与调制的时间比率。
表示调制快放电的相关的慢变量,比如,慢钾电流的门控变量、内钙离子浓度等。小参数表示快慢时间尺度的比率,也即放电与调制的时间比率。
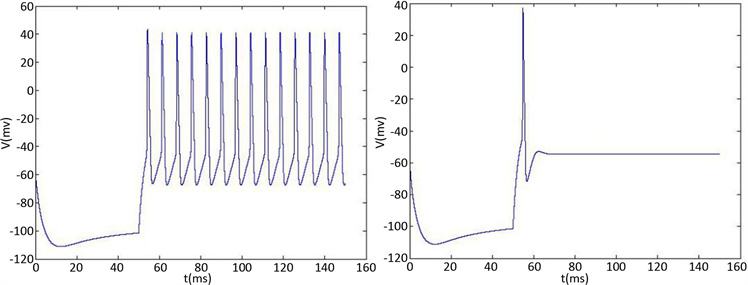
为了分析簇放电的性质和产生机理,我们假设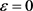 ,这样就可以分开讨论快子系统和慢子系统。快子系统可以是静息的(但是兴奋地),双稳的,或放电的,这取决于y的值。当y的值变化时,可引起簇放电的放电态与静息态的周期性地变化时,因此簇放电的许多重要行为可以通过快子系统的相图加以分析,其中,慢变量y看成是快子系统的分岔参数。
,这样就可以分开讨论快子系统和慢子系统。快子系统可以是静息的(但是兴奋地),双稳的,或放电的,这取决于y的值。当y的值变化时,可引起簇放电的放电态与静息态的周期性地变化时,因此簇放电的许多重要行为可以通过快子系统的相图加以分析,其中,慢变量y看成是快子系统的分岔参数。
 (a1) (a2)
(a1) (a2)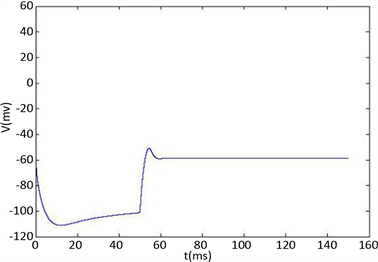 (a3)
(a3)
Figure 1. Time course of the membrane potential V stimulated by square wave current stimulated by hyperpolarization and depolarization of the KM model at different . (a1)~(a3) The current response of the KM model. The selected depolarization current is 140 pA, and the 50 ms hyperpolarization current is −200 pA. The change of
. (a1)~(a3) The current response of the KM model. The selected depolarization current is 140 pA, and the 50 ms hyperpolarization current is −200 pA. The change of  is 1, 0.5, 0.05 in turn
is 1, 0.5, 0.05 in turn
图1. KM模型在不同 下先超极化后去极化的方波电流刺激的膜电位V的时间历程图。(a1)~(a3) KM模型的电流响应,选择的去极化电流140 pA,50 ms超极化电流为−200 pA,
下先超极化后去极化的方波电流刺激的膜电位V的时间历程图。(a1)~(a3) KM模型的电流响应,选择的去极化电流140 pA,50 ms超极化电流为−200 pA,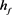 的变化依次为1,0.5,0.05
的变化依次为1,0.5,0.05
由于DCN椎体神经元同时接受兴奋型突触刺激和抑制型突触刺激,在离体实验和对模型的数值仿真时均采用先超极化后去极化的方波电流刺激,体内的放电模式可以在体外复制 [7]。由图1可知,随着快钾电流的失活变量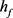 在0~1区间中的变化,KM模型膜电位V的时间历程图随着
在0~1区间中的变化,KM模型膜电位V的时间历程图随着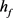 的减小(
的减小(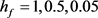 )均表现从规则簇放电到单尖峰放电再到接近静息放电的状态。再从图2中KM模型
)均表现从规则簇放电到单尖峰放电再到接近静息放电的状态。再从图2中KM模型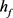 的时间序列可以看出,
的时间序列可以看出,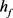 在神经元的超极化过程中不断增加,直到饱和到接近于1,在去极化电流注入时,快钾电流通道能够瞬时激活阻止由于钠通道的大量开放所导致的动作电位的发放。但经过一段时间的去极化,快钾离子通道逐渐关闭,再也不能阻止膜电位的增加,发生重复放电行为。因此可以将
在神经元的超极化过程中不断增加,直到饱和到接近于1,在去极化电流注入时,快钾电流通道能够瞬时激活阻止由于钠通道的大量开放所导致的动作电位的发放。但经过一段时间的去极化,快钾离子通道逐渐关闭,再也不能阻止膜电位的增加,发生重复放电行为。因此可以将 其视为快子系统的分岔参数,我们针对
其视为快子系统的分岔参数,我们针对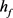 对KM模型进行分岔分析。
对KM模型进行分岔分析。
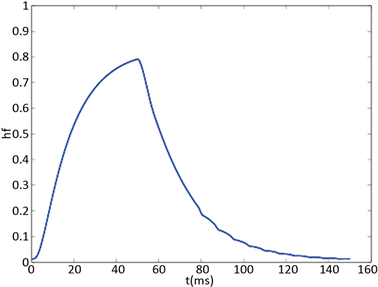
Figure 2. KM model (a) Time history of fast potassium current shutdown probability variable . In the hyperpolarized state,
. In the hyperpolarized state, 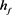 gradually increases, and in the depolarized state,
gradually increases, and in the depolarized state, 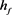 gradually decreases. The selected depolarization current is 140 pA, and the 50 ms hyperpolarization current is −200 pA
gradually decreases. The selected depolarization current is 140 pA, and the 50 ms hyperpolarization current is −200 pA
图2. KM模型(a)快钾电流关闭概率变量 的时间历程图。在超极化状态下
的时间历程图。在超极化状态下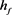 逐渐增加,在去极化状态下
逐渐增加,在去极化状态下 逐渐减小。选择的去极化电流140 pA,50 ms超极化电流为−200 pA
逐渐减小。选择的去极化电流140 pA,50 ms超极化电流为−200 pA
3.2. KM模型的快慢动力学分岔分析
通过3.1的快慢动力学分析,我们选择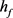 作为分岔参数。我们选定50 ms超极化电流为
作为分岔参数。我们选定50 ms超极化电流为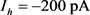 ,去极化电流
,去极化电流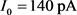 ,当参数
,当参数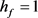 时,KM模型的快子系统处于一个稳定的平衡点(红线)。当
时,KM模型的快子系统处于一个稳定的平衡点(红线)。当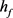 逐渐减小到0.24时,KM快子系统发生Hopf分岔,系统由一个稳定点变为稳定的极限环(蓝线)。当平衡点由于Hopf分岔消失后,神经元开始重复放电(蓝灰线)。本文中的分叉图是使用XPPAUT软件和MATLAB (ODE15S积分)生成的。如图3所示。
逐渐减小到0.24时,KM快子系统发生Hopf分岔,系统由一个稳定点变为稳定的极限环(蓝线)。当平衡点由于Hopf分岔消失后,神经元开始重复放电(蓝灰线)。本文中的分叉图是使用XPPAUT软件和MATLAB (ODE15S积分)生成的。如图3所示。
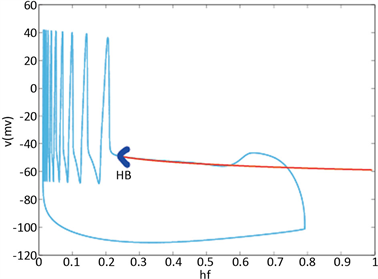
Figure 3. Analysis of fast and slow dynamic bifurcation of KM model. The slow variable  is a parameter, the red line represents the stability point of the fast subsystem, and the blue line represents the maximum and minimum values of the limit cycle. The blue-grey line indicates the trajectory of the system under the stimulation of hyperpolarized current
is a parameter, the red line represents the stability point of the fast subsystem, and the blue line represents the maximum and minimum values of the limit cycle. The blue-grey line indicates the trajectory of the system under the stimulation of hyperpolarized current 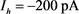 and depolarized current
and depolarized current 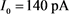
图3. KM模型的快慢动力学分岔分析。慢变量 为参数,红线表示快子系统的稳定点,蓝线表示极限环的最大值和最小值。蓝灰线表示系统在超极化电流
为参数,红线表示快子系统的稳定点,蓝线表示极限环的最大值和最小值。蓝灰线表示系统在超极化电流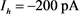 和去极化电流
和去极化电流 刺激下的轨线
刺激下的轨线
4. KM模型的Hopf分岔控制
4.1. Hopf分岔出现的条件
对于式子: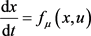 ,其中
,其中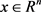 是状态向量,
是状态向量,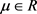 是分岔参数,
是分岔参数,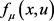 是从
是从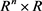 到
到 的光滑映射,u是标量输入。假设参数值
的光滑映射,u是标量输入。假设参数值 存在固定点
存在固定点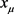 ,特征值为
,特征值为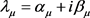 。假设
。假设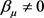 ,
, ,并且
,并且 ,则参数值
,则参数值 时发生Hopf分岔。
时发生Hopf分岔。
在一些系统中,特别是在高维系统中,不能总是获得特征值。为了避免这种情况,我们可以使用基于Routh-Hurwitz稳定性判据的Hopf分岔出现的等价条件 [8]。假设雅可比矩阵的特征多项式为:
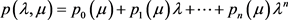
我们假设矩阵 为
为
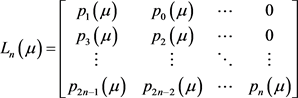
当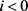 或者
或者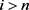 时,
时, 。我们再令
。我们再令
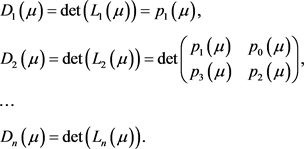
当满足以下条件时,系统在 时经历Hopf分岔:
时经历Hopf分岔:
1) 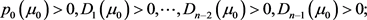
2) 
4.2. 冲洗滤波器的动态状态反馈控制律
基于冲洗滤波器的动态状态反馈控制律用于控制系统中的Hopf分岔,我们引入以下式子:

其中, ,
,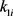 是控制收益,
是控制收益,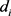 是滤波器时间常数的倒数。控制器是一个线性控制器,需要满足两个条件,一个是
是滤波器时间常数的倒数。控制器是一个线性控制器,需要满足两个条件,一个是 以保证滤波器的稳定性,还有一个是
以保证滤波器的稳定性,还有一个是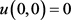 以保留初始平衡点。
以保留初始平衡点。
由于膜电位V很容易测量,我们选择它作为要控制的输入。加入滤波器后,受控制的KM模型变为:

4.3. 受控制的KM模型
我们选择将Hopf分岔推进到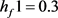 ,为了实现我们的目标,我们使用Routh-Hurwitz稳定性判据得到控制增益
,为了实现我们的目标,我们使用Routh-Hurwitz稳定性判据得到控制增益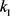 的值。令
的值。令 ,在
,在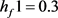 时:
时:

雅可比矩阵是:
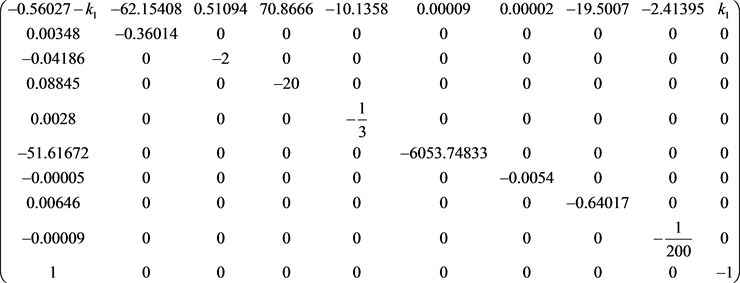
我们得到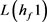 的特征多项式:
的特征多项式:
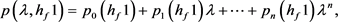
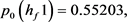
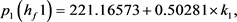
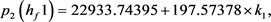

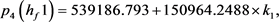
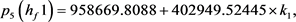
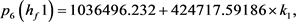
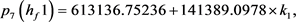



根据Hopf分岔1)和2)的等价条件,我们可以得到:
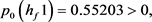
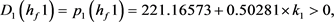
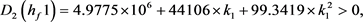

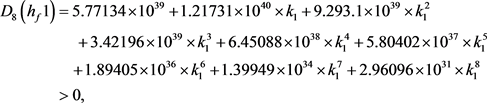

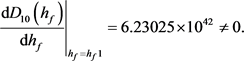
求解九个不等式和一个等式,得到 。
。
图4显示Hopf分叉的位置已成功地从 控制到
控制到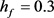 。我们重新选择合适的超极化电流为
。我们重新选择合适的超极化电流为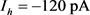 。当
。当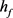 逐渐减小到0.3时,系统发生Hopf分岔,由一个稳定点(红线)变为稳定的极限环(蓝线)。当稳定的平衡点由于Hopf分岔消失后,神经元开始重复放电(蓝灰线)。
逐渐减小到0.3时,系统发生Hopf分岔,由一个稳定点(红线)变为稳定的极限环(蓝线)。当稳定的平衡点由于Hopf分岔消失后,神经元开始重复放电(蓝灰线)。
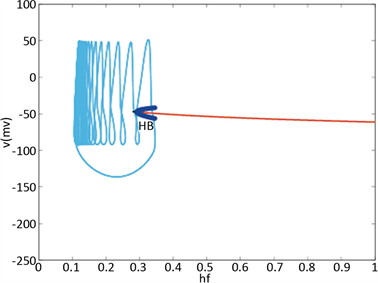
Figure 4. KM model after control. The slow variable  is a parameter, the red line represents the stability point of the fast subsystem, and the blue line represents the maximum and minimum values of the limit cycle. The blue-grey line indicates the trajectory of the system under the stimulation of hyperpolarized current
is a parameter, the red line represents the stability point of the fast subsystem, and the blue line represents the maximum and minimum values of the limit cycle. The blue-grey line indicates the trajectory of the system under the stimulation of hyperpolarized current 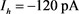 and depolarized current
and depolarized current 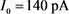
图4. 控制后的KM模型。慢变量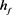 为参数,红线表示快子系统的稳定点,蓝线表示极限环的最大值和最小值。蓝灰线表示系统在超极化电流
为参数,红线表示快子系统的稳定点,蓝线表示极限环的最大值和最小值。蓝灰线表示系统在超极化电流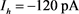 和去极化电流
和去极化电流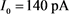 刺激下的轨线
刺激下的轨线
5. 结论
本文主要研究了DCN椎体神经元的Hopf分岔控制。文章的研究具有一定的创造性,内容表述清楚,分析清晰严谨,公式准确,研究过程及结果具有较好的应用与参考价值,值得发表。
文章引用
王 子汉,伍 帅. DCN椎体神经元的Hopf分岔控制
Hopf Bifurcation Control of DCN Pyramidal Neurons[J]. 应用数学进展, 2020, 09(02): 133-141. https://doi.org/10.12677/AAM.2020.92016
参考文献
- 1. Luo, M., Wu, Y.J. and Peng, J.H. (2009) Washout Filter Aided Mean Field Feedback Desynchronization in an En-semble of Globally Coupled Neural Oscillators. Biological Cybernetics, 101, 241-246.
https://doi.org/10.1007/s00422-009-0334-5 - 2. Crawford, J.D. (1991) Introduction to Bifurcation Theory. Review of Modern Physics, 63, 991-1038.
https://doi.org/10.1103/RevModPhys.63.991 - 3. Huang, C.L., Sun, W., Zheng, Z.G., et al. (2016) Hopf Bi-furcation Control of the M-L Neuron Model with Type I. Nonlinear Dynamics, 87, 755-766.
- 4. Xie, Y., Aihara, K. and Kang, Y.M. (2008) Change in Types of Neuronal Excitability via Bifurcation Control. Physical Review E, 77, Article ID: 021917.
https://doi.org/10.1103/PhysRevE.77.021917 - 5. Nguyen, L.H., Hong, K.S. and Park, S. (2012) Bifurcation Control of the Morris-Lecar Neuron Model via a Dynamic State-Feedback Control. Biological Cybernetics, 106, 587-594.
https://doi.org/10.1007/s00422-012-0508-4 - 6. Kanold Patrick, O. and Manis Paul, B. (2001) A Physiologically Based Model of Discharge Pattern Regulation by Transient K+ Currents in Cochlear Nucleus Pyramidal Cells. Journal of Neurophysiology, 85, 523-538.
https://doi.org/10.1152/jn.2001.85.2.523 - 7. Augustine, G.J. (1990) Regulation of Transmitter Release at the Squid Giant Synapse by Presynaptic Delayed Rectifier Potassium Current. The Journal of Physiology, 431, 343-364.
https://doi.org/10.1113/jphysiol.1990.sp018333 - 8. Liu, W.M. (1994) Criterion of Hopf Bifurcations with-out Using Eigenvalues. Journal of Mathematical Analysis and Applications, 182, 250-256.
https://doi.org/10.1006/jmaa.1994.1079