Climate Change Research Letters
Vol.
11
No.
04
(
2022
), Article ID:
53741
,
16
pages
10.12677/CCRL.2022.114051
基于通量观测的温带森林碳交换变化特征及影响因素研究
沈戈弋1,柴美琪2,吴博文2
1阿拉山口市气象局,新疆 博州
2博州气象局,新疆 博州
收稿日期:2022年6月13日;录用日期:2022年7月11日;发布日期:2022年7月18日

摘要
随着全球经济的高速发展,二氧化碳总排放量越来越高,在这个环境问题日益严重的紧要关头,森林在植物光合固碳这一过程中扮演着关键的角色;中国温带森林面积大、蓄积量高,因此作为我国研究森林固碳的重要对象。本论文研究对象为长白山阔叶红松林,利用公式GPP = −(Fc − Re)拆分为呼吸作用与光合作用两种不同通量,并在不同时间尺度上分析其变化特征,从而得出生态系统净二氧化碳交换量(Fc)在不同时间尺度上的变化趋势,可得知Fc日平均尺度与月尺度基本保持一致。日尺度上森林生态系统在日间由于光合作用多表现为碳汇,夜晚因呼吸作用表现为碳源;月尺度上,Fc的季节尺度变化总体呈负值,则表示除了程度区分之外所有月份都处于吸收CO2,及碳汇的状态;提取环境因子数据与Re、GPP做相关性分析,检测相关性是否显著;结果表明:太阳辐射及土壤温度为主要相关因子,达到显著相关水平。2007年长白山阔叶红松林处于吸收二氧化碳的“碳汇”状态。
关键词
温带森林,碳交换,生态系统总初级生产力,呼吸作用,光合作用

The Study on Temporal Variation and Impact Factors of Carbon Exchange in Temperate Forest Based on Eddy Flux
Geyi Shen1, Meiqi Chai2, Bowen Wu2
1Alashankou Meteorological Bureau, Bozhou Xinjiang
2Bozhou Meteorological Bureau, Bozhou Xinjiang
Received: Jun. 13th, 2022; accepted: Jul. 11th, 2022; published: Jul. 18th, 2022

ABSTRACT
With the rapid development of the global economy, the total carbon dioxide emissions are getting higher and higher, at this critical moment when the environmental problem is becoming more and more serious. Forests play a key role in the process of plant photosynthesis carbon sequestration, and China’s temperate forests have a large area and high accumulation, so they are important objects for the study of forest carbon sequestration in China. The object of this paper is the broad-leaved red pine forest of Changbai Mountain, which is divided into two different fluxes of respiratory action and photosynthesis by using the formula GPP-Re, and analyzing its change characteristics on different time scales so as to obtain the net carbon dioxide of the ecosystem. The changing trend of exchange quantity (Fc) on different time scales shows that the average scale of Fc day is basically consistent with the monthly scale, and those forest ecosystems on the daily scale are represented as carbon sinks during the day due to photosynthesis, and nighttime as carbon sources due to respiratory action. On the monthly scale, Fc’s seasonal scale change was negative overall, indicating that all months except the degree distinction were in the state of absorbing CO2 and carbon sinks. The correlation between the extraction of environmental factor data and Re and GPP was analyzed to detect whether the correlation was significant, and the results showed that solar radiation and soil temperature were the main correlation factors. In 2007, Changbaishan broad-leaved red pine forest was in the “carbon sink” state of absorbing carbon dioxide.
Keywords:Temperate Forest, Carbon Exchange, Gross Primary Productivity, Photosynthesis, Ecosystem Respiration

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
近年来,人们对于大气中二氧化碳浓度增加可能导致全球变暖的问题日益关注,这主要是化石燃料燃烧而导致的,从而也影响着陆地生态系统,包括森林、农作物与水资源等等,因此在1997年12月制定了《京都议定书》。根据该议定书,各国可以通过限制排放来减少排放量 [1]。全球碳收支表明,大量的二氧化碳被陆地的生态系统所吸收,北半球的温带和寒带森林可能是最重要的陆地汇。对森林冠层范围内的二氧化碳和水蒸气通量的长期测量最近才成为可能 [2]。因此,到目前为止对温带森林的二氧化碳交换情况的研究,还不够深入。生态系统总初级生产力,简称为GPP,它是指在单位时间内,植物在光合作用下所产生的有机碳总量,在地球下垫面–大气系统的碳交换中,GPP所表现出的作用可谓相当明显和突出。为对全球碳循环过程展开积极研究,加大对森林、草地生态系统碳循环过程的深入研究,对于全球气候问题的解决具有非常重要的作用。森林是全球三大生态系统之一,在陆地生态系统中,更是如同碳库 [3],基于全球气候不断发生改变的背景下,对于不同森林森林生态系统碳交换动态特征加以量化,同时分析其环境影响因子,对全球温室效应以及更好地了解生物地球化学过程具有重要的实践和理论意义 [4]。
中国温带森林的总面积、蓄积量在全国森林中的占比分别为35.3%、34.5%,由此也体现出了中国温带森林的重要性,甚至还在一定程度上影响着国家以及世界碳估算的情况,对其加以量化分析,客观评价显得十分重要和必要。因相关人为因素的影响,导致CO2生态系统净交换量变化差异较大 [5] [6] [7] [8]。全球变化生态系统碳循环的重要过程之一就是生态系统光合作用,其中,绿色植物的光合作用就是在可见光的照射影响下,体内的叶绿素等各种光合色素将水与二氧化碳转化为有机物,最终成功释放出氧气,所体现出的是一个生物化学过程。整个生物圈与大气被植物的光合固碳连接在一起,每年约有120 Pg的碳通过光合作用被陆地生态系统固定,这些被固定的碳为人类提供了食物和材料,同时很大程度上也推动了全球碳循环。食物链中的生产者是植物,在光合作用下,无机物产生有机物之后从而达到储存能量的目的。总之,植物想要实现生存就必须要依赖光合作用 [9]。
光合作用的意义不言而喻,比如:提供物质来源;稳定大气中的氧气含量等等,除此之外,还对生物的进化起到了非常明显的作用。特别是从工业革命之后,人类燃烧化石燃料释放的二氧化碳等温室气体引发了各种环境问题,所以,靠植物提高光合固碳就对缓解如全球变暖等一系列环境问题来说十分重要。
CO2通量主要是指CO2流通的量,但一定是在单位时间单位面积内所实现的,单位一般表示为μmol·m−2·s−1。与大气和海洋部分相比,陆地生态系统碳循环仍然是全球碳收支中受约束最少的部分 [10]。全球碳收支和过程模型预测的准确性在很大程度上取决于生态系统层面对二氧化碳(CO2)流的准确测量 [11]。各种各样的方法都被用来估算陆地二氧化碳通量,如涡度相关测量(EC)、生物测量、静态箱测量法等。
2. 研究地点和方法介绍
2.1. 研究地点简介
将长白山作为此次研究的主要地点,它位于中朝两国的交界处,地理位置比较特殊。从地质属性进行分析,它属于休火山,而海拔为2691 m的白云峰,则是东北地区的第一高峰,山势陡峭,直矗云霄, 1800米以下为熔岩高原和台地,地势倾斜平缓。土壤、植被主要呈现出了垂直分布的态势,且受到地势高程变化因素的影响,从而也构成了温带大陆东岸典型的山地垂直景观带谱,即便是从全球自然生态系统角度分析,长白山作为亚洲东部典型的山地森林生态系统,保存可谓非常的完整。
该地区由于地处中纬度且濒临太平洋,且容易受到季风的影响,所以属于温带大陆性山地气候,一年四季,冬季时间最长,几乎接近7个月,尤其是海拔较高的地区,几乎没有夏天,全年平均气温多在4.9℃~−7.3℃之间。由于全年气温都相对比较低,降水较为充足,加上森林本身所具有的涵养作用,从而使得长白山的湿度高达70%左右,并呈现出了水量丰富、热量不足的水热状况,整体上的湿润状况显得非常优良。
2.2. 观测仪器概述
为了对垂直尺度上森林微气象进行科学观测,还专门在长白山阔叶红松林内,建设了一个高度为62 m的微气象观测塔,从冠下到冠上还配备了专门的观测平台,多达9个。不仅如此,在森林冠上和冠下还都安装有一套开路式涡度相关系统,目的是对森林、大气和土壤、大气间CO2能量彼此之间的交换情况,进行科学观测。而为了实现对森林气象因子的连续性测定,技术人员还在观测塔上安装了七层常规气象观测系统。
安装在微气象观测塔上的小气候梯度观测系统,所发挥的作用可谓相当多样,比如:对7个不同层次的大气温度、湿度等因子,3个不同层次的土壤湿度进行观测等等。所有的传感器分别同3个CR10X,1个CR23X型数据采集器有效连接在一起,将采样频率设置为0.5 Hz,对于平均值加以自动化记录,间隔时间为半个小时。
2.3. 研究方法
在计算生态系统净二氧化碳交换量(Fc)时,因观测数据存在少数异常或缺测数据,为计算碳通量变化的日总量、月总量和年总量,需要对异常或缺测数据进行数据插补。文献分析呼吸作用与温度有较高相关性,光合作用手=受太阳辐射强度的影响较大,所以我们利用夜间的温度与生态系统净二氧化碳交换量进行数据插补,首先利用白天现有太阳辐射(Rn)的观测数据建立白天Fc~Rn的拟合方程:
(1)
之后利用拟合方程计算出白天Fc的数据,以完成数据插补。
依据相同的操作,实现对夜间的数据插补,在建立拟合方程的时候,应对夜间夜间Fc与气温的观测数据进行思考和分析:
(2)
然后利用该拟合方程计算夜间Fc的缺测数据,完成数据插补。
为进一步分析碳通量变化特征,我们将Fc拆分为总初级生产力(GPP)及生态系统呼吸(Re)两个通量,并利用Fc与气温的拟合方程:
(3)
代入白天的气温数据,结果作为白天Re的数据。
随后利用公式:
(4)
代入Fc正常观测数据与插补后的数据,从而计算出GPP的数据。
3. 结果分析
3.1. 生态系统碳交换量(Fc、Re、GPP)的日变化
3.1.1. 碳通量(Fc、Re、GPP)平均日变化
在分析碳交换量的日变化时,为较为准确的了解不同时间尺度内的变化,我们将日变化按照季节尺度划分为四个图例(如下若干图),将12~2月的数据列为冬季(d),3~5月的数据列为春季(a),6~8月的数据列为夏季(b),9~11月的数据列为秋季(c);长白山混交林的碳通量交换的变化趋势基本较为稳定,但是变化趋势的幅度在夏季有较大差异,夏季(图1(b)所示),在中午12时的碳交换量最低,Fc的值约为−0.52 mg·m−2·s−1,说明此时该混交林此时期的碳吸收较强。而到了冬季,碳交换基本呈一条直线,并与其他季节相比有较为明显的下降,说明与其他季节相比,该地区冬季的混交林对碳的吸收为最弱的时期,Fc的值约为−0.08 mg·m−2·s−1。由图1所示,该地区混交林在四个季节都是在日出后(早晨7时)开始吸收CO2,因此可得出,当Fc的值小于0时说明植被在吸收CO2,当Fc为正值时表示生态系统处于释放CO2的状态,森林生态系统在日间由于光合作用多表现为碳汇,夜晚因呼吸作用表现为碳源 [12]。
在对碳交换量日变化分析时,由于Fc的相关变化包含该地区植物的光合作用以及生态系统呼吸作用,所以将Fc拆分为生态系统呼吸(Re)与生态系统总初级生产力(GPP),并分析这两个通量各个季节的变化趋势(如图2(a)~(d)所示);首先可以看出,该地区混交林的光合作用效率日变化在夏季最为显著:Re的变化范围为0.09~0.14 mg·m−2·s−1;GPP的变化范围为0~0.67 mg·m−2·s−1冬季的变化趋势最为平缓:Re的变化范围为−0.01~0.01 mg·m−2·s−1;GPP的变化范围为0~0.06 mg·m−2·s−1;呼吸速率(Re)在四个季节变化较小,其范围为春季:0.02~0.06 mg·m−2·s−1,夏季:0.09~0.14 mg·m−2·s−1,秋季:0.02~0.07 mg·m−2·s−1,
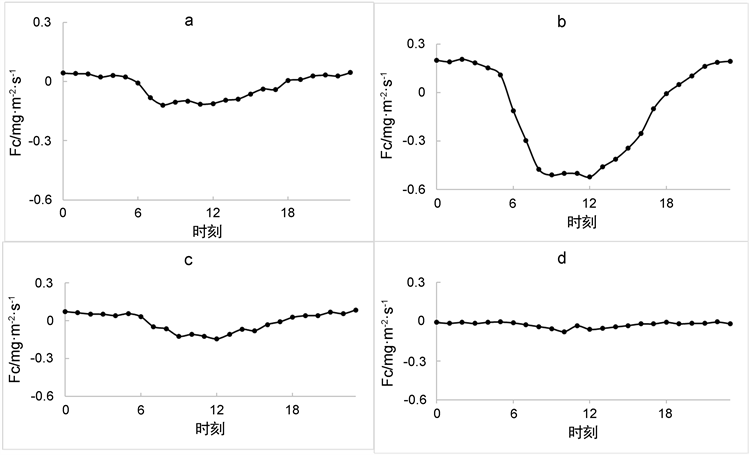
Figure 1. Average daily change of Fc. ((a) For spring, (b) For summer, (c) For autumn, (d) For winter)
图1. Fc平均日变化。((a) 为春季,(b) 为夏季,(c) 为秋季,(d) 为冬季)
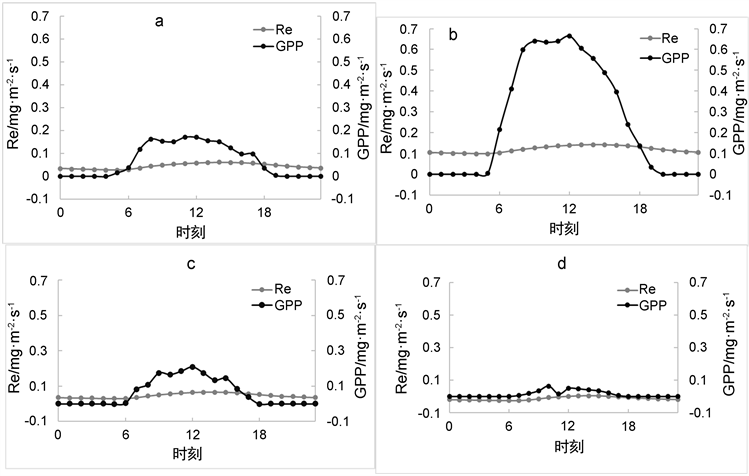
Figure 2. Average daily change of Re and GPP ((a) For spring, (b) For summer, (c) For autumn, (d) For winter)
图2. Re、GPP平均日变化((a) 为春季,(b) 为夏季,(c) 为秋季,(d) 为冬季)
冬季:−0.01~0.01 mg·m−2·s−1。对于每日的Re与GPP变化来看,日出(早晨7时)后光合作用速率开始渐渐大于呼吸作用的速率,并在中午12时~下午13时之间达到顶峰,此结果也对应了Fc相关数据的变化特征,呼吸作用在夏季下午14时达到顶峰,峰值为0.14 mg·m−2·s−1,且夏季的变化量 > 秋季的变化量 > 春季的变化量 > 冬季的变化量。由图2中趋势线可得出,从一月(冬季)至七月(夏季) GPP增长率在持续升高,从八月(秋季)开始逐渐降低。所以该地区混交林的光合作用在夏季处于最强且较为稳定的时期。
气温以及太阳辐射也对碳通量的交换的日变化产生了一定的影响,光合作用的完成需要一定的太阳辐射以及特定范围的温度,而呼吸作用也与气温相关。由图3(a)、图3(c)可得,春秋两季的气温相差不大,太阳辐射春天较强,其范围为−60~423 W·m−2,从而导致与秋季(太阳辐射值范围为:−55~330 W·m−2)相比,春季的GPP值要稍高于秋季GPP的值,;夏季气温与太阳辐射明显是最高的其范围为:夏季气温值为14℃~23℃,太阳辐射值为−47~487 W·m−2,于是也影响夏季光合作用强于其他三个季节。
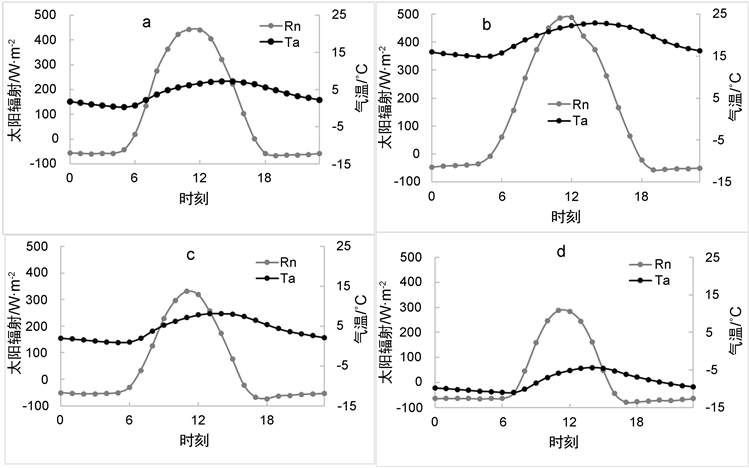
Figure 3. Daily average changes in solar radiation and temperature ((a) For spring, (b) For summer, (c) For autumn, (d) For winter)
图3. 太阳辐射及气温的日平均变化((a) 为春季,(b) 为夏季,(c) 为秋季,(d) 为冬季)
日出半小时左右,碳通量的值由正值逐渐转变成为负值,这也意味着从之前的释放变成了吸收。太阳辐射受到太阳高度角逐渐增大的影响,随之有所增强,一般在中午12时左右,Fc不断增大并达到顶峰,Fc的值因为太阳辐射量的逐渐降低,也将趋于逐渐减小,傍晚19时左右由负转正。
不同季节的GPP值都呈“U”型曲线变化趋势,这种变化趋势的主要原因就是早晨随着太阳辐射及气温的增加,该生态系统的光合作用的强度开始不断增大,由图2及图3可知,GPP的变化趋势在11时至13至到达峰值(0.67 mg·m−2·s−1),然后伴随着太阳辐射减少,气温降低,光合作用的强度也开始慢慢减弱,导致该生态系统的生产力降低,在晚上20时降至最低(0 mg·m−2·s−1);而Re的变化并没有很显著的特征,由于日夜交替,生态系统的呼吸值会大幅下降,主要是由于夜间生态系统只有呼吸作用;日出之后,由于太阳辐射以及气温不断增大,该生态系统开始吸收大气中的CO2,导致生态系统的呼吸作用急剧减弱,但是当太阳辐射以及气温逐渐升高,呼吸作用也开始增强,日落时间之后,由于太阳辐射及气温减小,呼吸作用也随之减弱,但由于夜间生态系统不吸收空气中的CO2,则呼吸作用也会逐渐增强。但由于本文所用数据为一个季度的日平均值,所以呼吸作用的变化趋势并不明显。王春林等人所做的鼎湖山常绿针阔叶混交林碳交易表明:将该地区2003~2004年期间的森林生态系统总体表现为碳汇,并认为在大气相对稳定的夜间,仅仅只是通过传感器并不能对在森林冠层呼吸作用下所产生的CO2进行有效探测,甚至还可能会对其通量观测值的探测造成巨大影响 [13]。所以Re与GPP的图示变化规律均符合物候变化规则。
相对湿度一天内的变化为:下午13时到15时为相对湿度最低的时刻,其变化范围为:春季44%~70%,夏季56%~84%,秋季46%~72%,冬季45%~68% (如图4所示);不同季节的日变化曲线并没有太大的区别。
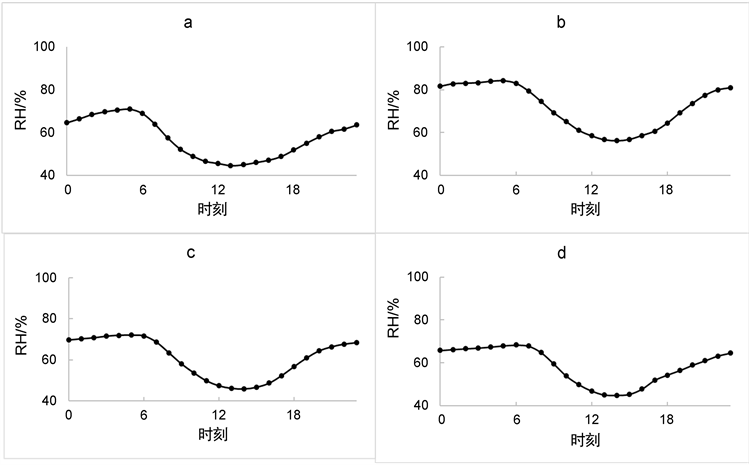
Figure 4. Average daily change in relative humidity ((a) For spring, (b) For summer, (c) For autumn, (d) For winter)
图4. 相对湿度平均日变化((a) 为春季,(b) 为夏季,(c) 为秋季,(d) 为冬季)
3.1.2. 典型天气碳通量(Fc、Re、GPP)日变化
由于上述数据皆为季度数据日平均数值,其中某些特殊天气情况可能需要单独描述,例如2007年6月27日及28日两天,分别为晴天与多云天气,由图5可以看出,是否晴天对于太阳辐射的影响还是较为明显的,例如图5中6月27日太阳辐射峰值在上午11时达到了687.2 W·m−2,而6月28日多云天气情况下的太阳辐射峰值在下午14时,为273.8 W·m−2。此时还处于该森林生态系统的生长期,而在2007年11月9日与12日分别处于多云天气与晴天,此时处于该森林生态系统的非生长期;11月12日上午11时,太阳辐射达到峰值为378.9 W·m−2,11月9日为多云天气,上午9时为太阳辐射峰值,仅仅只达到了187.4 W·m−2,由此可见不同天气情况对太阳辐射的会产生较为直观的影响,从而间接影响碳交换量。除此之外由图5、图6还可以看出植物生长期所接受的太阳辐射要强于非生长期。

Figure 5. Changes in solar radiation in typical weather (sunny and cloudy days during the growing season)
图5. 典型天气太阳辐射变化(生长期晴天和多云天)
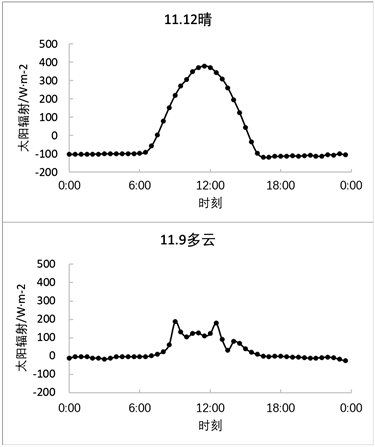
Figure 6. Solar radiation diurnal variations in typical weather conditions (sunny and cloudy days during the non-growing season)
图6. 典型天气情况太阳辐射日变化(非生长期晴天和多云天)
对于Fc在两种天气情况下的日变化趋势进行分析可知,该森林生态系统处于生长期时Fc的日变化较为明显(如图7、图8所示),生长期晴天的Fc日变化范围为−0.9~0.5 mg·m−2·s−1,非生长期晴天的Fc日变化范围为−0.5~0.2 mg·m−2·s−1。同一时期的森林在不同天气情况下的Fc变化值也不同,晴天的Fc变化值与多云天气相比趋势更加平滑,多云天Fc的变化范围在生长期为−1.2~0.9 mg·m−2·s−1且包含很多急升及骤降的情况;在非生长期的变化范围为−1.3~0.7 mg·m−2·s−1,数据间差异范围较大,说明天气情况对太阳辐射产生的影响间接影响了Fc的变化趋势。
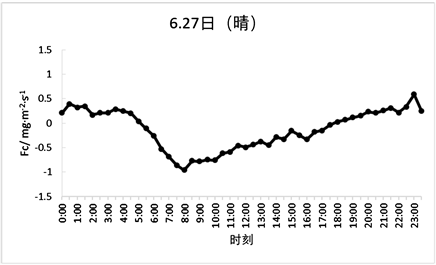

Figure 7. Typical weather Fc day variation (sunny and cloudy days during the growing season)
图7. 典型天气Fc日变化(生长期晴天和多云天)

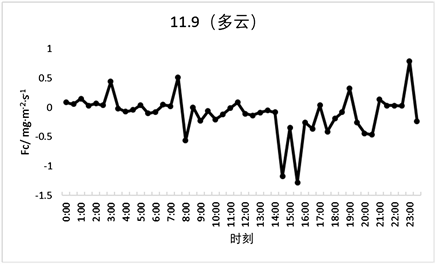
Figure 8. Typical weather Fc day variation (sunny and cloudy days during non-growing period)
图8. 典型天气Fc日变化(非生长期晴天和多云天)
Re在生长期与非生长期的变化范围基本持平,但呼吸速率有较大差异,如图9与图10所示,非生长期Re的变化范围为−0.01~0.06 mg·m−2·s−1,生长期的变化范围为0.12~0.18 mg·m−2·s−1,由此可得该森林生态系统处于生长期时的呼吸作用要强于非生长期。GPP也有同样的变化状态,在非生长期时GPP的变化范围是0~0.5 mg·m−2·s−1,非生长期的变化范围是0~1.1 mg·m−2·s−1,由此可得,处于生长期的森林生态系统的光合作用强于非生长期,因此该森林生态系统处于生长期时的固碳能力要高于非生长期。
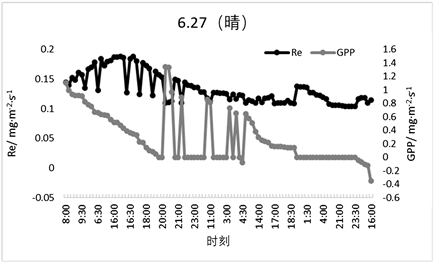

Figure 9. Typical weather Re and GPP daily variations (sunny and cloudy days during the growing season)
图9. 典型天气Re、GPP日变化(生长期晴天和多云天)
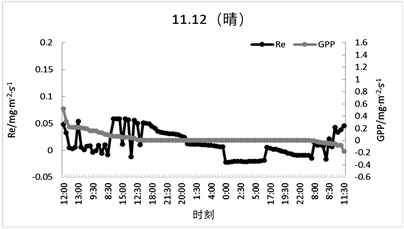

Figure 10. Typical weather Re and GPP diurnal variations (sunny and cloudy days during the non-growing season)
图10. 典型天气Re、GPP日变化(非生长期晴天和多云天)
3.2. 生态系统碳交换量(Fc、Re、GPP)的季节变化
图11表明:Fc的季节尺度变化总体呈负值,则表示除了程度区分之外所有月份都处于吸收CO2,及“碳汇”的状态,Fc的变化范围为−0.14~0.01 mg·m−2·s−1,10月份的数据可能因湍流运动或其他观测偏差导致了Fc呈现正值,即“碳源”的状态。由此可得该生态系统于2007年整体扮演着“碳汇”的角色,相对湿度在冬季还是处于缓慢增长的状态(52%~79%),在进入春季时陡降,直到秋季结束基本处于平稳增长的状态,在8月份达到峰值79%。

Figure 11. The monthly average change of Fc
图11. Fc的月平均值变化
由图12可以看出,直到1~4月及10~12月GPP与Re两个通量的差距并不大,而在5~9月光合作用的效率明显高于呼吸作用,而GPP在7月达到峰值0.29 mg·m−2·s−1,其变化范围为0.01~0.28 mg·m−2·s−1,可以代表植物光合作用增强幅度最大的时期,Re在1~8月处于平稳增长的状态,达到峰值0.12 mg·m−2·s−1,从8月之后缓慢降低,其变化范围为−0.02~0.12 mg·m−2·s−1。
太阳辐射、气温的季节性变化,都会对生态系统呼吸作用变化造成不同程度的影响,由图12可以看出,2007年该地区月平均最高气温为8月,达到了19.2℃,气温总体呈先增长,达到峰值后开始下降的趋势,符合物候规律。
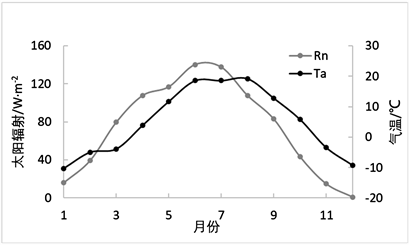
Figure 12. Monthly average change of Re and GPP
图12. Re、GPP的月平均值变化
图13表明:Fc的季节尺度变化总体呈负值,则表示除了程度区分之外所有月份都处于吸收CO2,及“碳汇”的状态,Fc的变化范围为−0.14~0.01 mg·m−2·s−1,10月份的数据可能因湍流运动或其他观测偏差导致了Fc呈现正值,即“碳源”的状态。由此可得该生态系统于2007年整体扮演着“碳汇”的角色,相对湿度在冬季还是处于缓慢增长的状态(52%~79%),在进入春季时陡降,直到秋季结束基本处于平稳增长的状态,在8月份达到峰值79%。

Figure 13. Monthly mean change of relative humidity (RH)
图13. 相对湿度(RH)的月平均值变化
3.3. 环境因子对生态系统碳交换量的影响
由下图(图14~17)可得,与该生态系统的碳交换量变化相关系数较高的环境因子为太阳辐射以及土壤温度(利用线性关系拟合方程,随后可得拟合方程以及其相关系数P < 0.01),随着太阳辐射值的增大,碳交换量也随之逐渐增大,土壤温度同理,该结果与张涛等(2018)研究所得出的结论一致——土壤温度为影响碳交换量变化的主要因素之一 [14];相反风速及相对湿度对于碳交换的影响并不显著,由于降雨对于碳交换的影响具有滞后性,因此影响也不直观显著。
之所以会影响到土壤的温度,在于那些同土壤有关的植物和微生物在呼吸作用下,新陈代谢过程中不断将CO2释放到大气,高温保持在一定程度,可以促进微生物及植物的新陈代谢,从而增加碳排放 [15]。除了土壤温度以外,由图2、图3也可以得出气温对该生态系统碳交换量有一定的相关性,即为随着空气温度升高时间点的提前或推迟,也会造成生态系统碳交换量变化的提前或推迟。

Figure 14. Effects of solar radiation on carbon exchange
图14. 太阳辐射对碳交换量的影响

Figure 15. Effect of 5 cm soil temperature on carbon exchange
图15. 5 cm土壤温度对碳交换量的影响

Figure 16. Effect of relative humidity on carbon exchange
图16. 相对湿度对碳交换量的影响
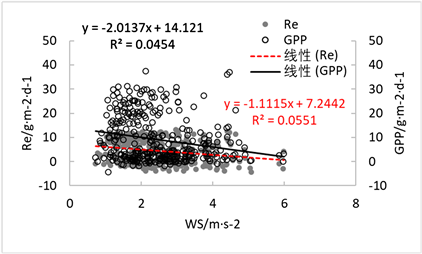
Figure 17. Effect of average wind speed on carbon exchange
图17. 平均风速对碳交换量的影响
4. 讨论
对于温带森林生态系统碳交换量的不同时间尺度的分析,可以得知该生态系统的碳交换存在很明显的日变化与季度变化,导致这种变化的原因除了周围环境因子对于植物的影响以外也包括植物本身是否处于生长期;与日平均尺度相比,月平均尺度(季度尺度)的GPP变化更为明显(其变化范围为0.01~0.28 mg·m−2·s−1),整年的变化趋势呈先增长后减小的状态,其中还存在波动范围较大的数据,主要原因为是否被特殊天气状况影响。Re与GPP在日尺度、季度尺度上会发生变化,主要是受到了“空气温度”这一因素的巨大影响。如图18,在日尺度上,环境因子变化对于Re产生的影响要大于GPP,而在季节尺度上则刚好相反。Re与GPP两种通量均会因为土壤温度的升高而增大,且与土壤温度相关显著。

Figure 18. The full-year trend of the monthly Average of Re and GPP
图18. Re与GPP月平均值的整年变化趋势
综合所有图表可以得出,该地区的此森林生态系统在2007年全年都处于“碳汇”的状态。
由于下垫面与大气的物质交换主要靠湍流运动完成,所以涡动相关法就成了测量陆地生态系统中碳交换的主要方法,这个方法要求的观测数据精确度较高,本文所用的数据因观测失误会存在一定偏差,观测数据时湍流运动以及某些植物冠层的呼吸作用不易被观测到,可能某些数据产生一定误差 [16] [17] [18]。目前对于森林冠层上平流作用及湍流运动对二氧化碳观测的影响还需要较为长久精细的研究 [18]。
相比较而言,学者们对于森林碳通量交换所展开的研究关注度明显不够,对于农田、森林生态系统所存在的差异进行分析:1) 农作物的不同时期对于环境因子的影响要更加敏感,变化也较为显著。2) 农田除了农作物自身的呼吸作用之外还存在土壤的呼吸作用影响,而森林冠层的碳通量检测就与土壤的呼吸作用没有明显的相关性。
生态系统最大的特征即是多样性,以致于关于森林碳交换的相关研究结论存在较大差异性,不仅如此,在碳循环研究过程中还存在很多因素,且存在诸多的不确定性。但不可否认,全球植被碳库、土壤碳库,在森林生态系统中重要存在,其占比分别高达86%,73%。学者们对于这一观点也表示认同。各种森林碳循环的研究方法中,较为有效的研究方法为模型方法 [19]。
除了研究我国温带自然生长的森林之外,有研究发现人造林的固碳价值也不可以忽略;张悦等人在对人造林的固碳能力进行研究时发现,在人造林生长季时表现为较强的“碳汇”状态,而在非生长季呈现较弱的“碳源”状态,从而可得人造林也有较强的固碳能力 [20]。
5. 结论
本文基于涡度相关系统的观测数据,将净生态系统碳通量Fc拆分为生态系统呼吸作用速率Re和总初级生产力GPP,并分析了其时间变化特征,以及环境因子的影响,得出以下结论:
1) Fc的值在日平均尺度上的变化要比月平均尺度更加显著,其主要由于植物呼吸作用在一定的温度范围内是一直进行的,而光合作用一般开始于日出,结束于日落,所以日平均尺度的变化会有较为明显的趋势线。
2) 生态系统碳交换量除了被太阳辐射与气温影响之外还会被其他环境因子所影响,如土壤温度等。同时还受植物本身所处的生长周期影响,处于生长期的植物呼吸作用与光合作用的作用强度比非生长期更高;且生长期的温度与太阳辐射都更适合光合作用更有效地进行,所以处于生长期的植物固碳能力也相对较好。
3) Re、GPP的变化量会直接导致Fc的变化,从而影响植物的固碳能力,对于森林生态系统而言,固碳能力不只是对该生态系统生命力的体现,也对全球环境问题有重大意义,目前对于森林的碳交换研究还不够全面,需要更加深入与细致的探索。
文章引用
沈戈弋,柴美琪,吴博文. 基于通量观测的温带森林碳交换变化特征及影响因素研究
The Study on Temporal Variation and Impact Factors of Carbon Exchange in Temperate Forest Based on Eddy Flux[J]. 气候变化研究快报, 2022, 11(04): 481-496. https://doi.org/10.12677/CCRL.2022.114051
参考文献
- 1. Pilegaard, K., Hummelshøj, P., Jensen, N.O. and Chen, Z. (2001) Two Years of Continuous CO2 Eddy-Flux Measure-ments over a Danish Beech Forest. Agricultural and Forest Meteorology, 107, 29-41. https://doi.org/10.1016/S0168-1923(00)00227-6
- 2. Baldocchi, D.D., Hicks, B.B. and Meyers, T.P. (1988). Measuring Biosphere-Atmosphere Exchanges of Biologically Related Gases with Micrometeorological Methods. Ecolo-gy, 69, 1331-1340. https://doi.org/10.2307/1941631
- 3. Kramer, P.J. (1981) Carbon Dioxide Concentration, Photosynthesis, and Dry Matter Production. BioScience, 31, 29-33. https://doi.org/10.2307/1308175
- 4. 李慧. 福建省森林生态系统NPP和NEP时空模拟研究[D]: [博士学位论文]. 福州: 福建师范大学, 2008.
- 5. Soegaard, H., Jensen, N.O., Boegh, E., Bay Hasager, C., Schelde, K. and Thomsen, A. (2003) Carbon Dioxide Exchange Overagricultural Landscape Using Eddy Correlation and Footprint Mod-eling. Agricultural and Forest Meteorology, 114, 153-173. https://doi.org/10.1016/S0168-1923(02)00177-6
- 6. 梁涛, 李荣平, 吴航, 刘景利, 史奎桥, 张兵兵, 等. 玉米农田生态系统CO2通量的动态变化[J]. 气象与环境学报, 2012, 28(3): 49-53.
- 7. 薛红喜, 李峰, 李琪, 王连喜, 王云龙, 胡正华. 基于涡度相关法的中国农田生态系统碳通量研究进展[J]. 南京信息工程大学学报: 自然科学版, 2012, 4(3): 226-232.
- 8. 李俊, 于强, 孙晓敏, 同小娟, 任传友, 王靖, 等. 华北平原农田生态系统碳交换及其环境调控机制[J]. 中国科学D辑: 地球科学, 2006, 36(S1): 210-223.
- 9. 朱苑. 帽儿山落叶阔叶林日间净生态系统碳交换特征[D]: [硕士学位论文]. 哈尔滨: 东北林业大学, 2021. https://doi.org/10.27009/d.cnki.gdblu.2020.000508
- 10. Le Quéré, C., Andrew, R.M., Canadell, J.G., Sitch, S., Korsbakken, J.I., Friedlingstein, P., Peters, G.P., Manning, A.C., Boden, T.A., Tans, P.P., Houghton, R.A., et al. (2016) Global Carbon Budget 2016. Earth System Science Data, 8, 605-694. https://doi.org/10.5194/essd-8-605-2016
- 11. Luyssaert, S., Reichstein, M., Schulze, E.D., Janssens, I.A., Law, B.E., Papale, D., Dragoni, D., Goulden, M.L., Granier, A. and Kutsch, W.L. (2009) Toward a Consistency Cross-Check of Eddy Covariance Flux-Based and Biometric Estimates of Ecosystem Carbon Balance. Global Biogeochemical Cycles, 23, Article No. GB3009. https://doi.org/10.1029/2008GB003377
- 12. 龚元, 纪小芳, 花雨婷, 张银龙, 李楠. 基于涡动相关技术的森林生态系统二氧化碳通量研究进展[J]. 浙江农林大学学报, 2020, 37(3): 593-604.
- 13. 王春林, 于贵瑞, 周国逸, 闫俊华, 张雷明, 王旭, 等. 鼎湖山常绿针阔叶混交林CO2通量估算[J]. 中国科学D辑: 地球科学, 2006, 36(S1): 119-129.
- 14. 张涛, 王根绪, 杨燕, 毛天旭. 青藏高原多年冻土区不同草地类型生态系统呼吸季节差异性[J]. 冰川冻土, 2018. 40(6): 1255-1264.
- 15. 魏甲彬, 徐华勤, 周玲红, 成小琳, 唐先亮, 傅志强, 等. “双季稻-冬闲田”生态系统碳交换动态变化及其影响因素[J]. 农业环境科学学报, 2018, 37(5): 1035-1044.
- 16. Xu, L.K. and Baldocchi, D.B. (2004) Seasonal Variation in Carbon Dioxide Exchange over a Mediterranean Annual Grassland in Cal-ifornia. Agricultural and Forest Meteorology, 123, 79-96. https://doi.org/10.1016/j.agrformet.2003.10.004
- 17. Katul, G., Lai, C.T., Schäfer, K., Vidakovic, B., Albertson, J., Ellsworth, D., et al. (2001) Multiscale Analysis of Vegetation Surface Fluxes: From Second Stoyears. Advances in Water Resources, 24, 1119-1132. https://doi.org/10.1016/S0309-1708(01)00029-X
- 18. 张弥, 温学发, 于贵瑞, 张雷明, 伏玉玲, 孙晓敏, 等. 二氧化碳储存通量对森林生态系统碳收支的影响[J]. 应用生态学报, 2010, 21(5): 1201-1209.
- 19. 范洪,胡军国, 赵明水, 毛国平. 基于WSN的天目山森林碳通量估算[J]. 环境科学与技术, 2019, 42(12): 230-236.
- 20. 唐祥. 华北典型人工针阔混交林碳通量研究——以北京八达岭林场为例[D]: [硕士学位论文]. 北京: 北京林业大学, 2014.