International Journal of Ecology
Vol.06 No.02(2017), Article ID:20820,12
pages
10.12677/IJE.2017.62013
Plant Diversity Characteristics of Wetland in the Middle Reaches of the Yellow River
Tianliang Zhao
Shanxi Institute of Forest Inventory and Planning, Taiyuan Shanxi

Received: May 11th, 2017; accepted: May 26th, 2017; published: May 31st, 2017

ABSTRACT
Taking the wetland plants in the middle reaches of the Yellow River as the research object, the phylogenetic tree of the wetland plants was constructed. Pedigree diversity index (PD), community lineage structure index (net genetic relationship index, NRI and recent interspecific relationship index, NTI), functional diversity index (FAD1, FDp, FDc, FRic, FEve, Rao and Dis) were used to analyze the phylogenetic diversity, pedigree structure and functional diversity of wetland plants in the middle reaches of the Yellow River were analyzed, and the species diversity index was also calculated. The main results are as follows: (1) Based on the APGIII classification system, the range of α diversity index (PD) of the 13 plots was calculated from the phylogenetic software Phylocom in the range of 706.894 - 2289.17, and the PD values were in the order of S1 > S12 > S3 > S6 > S2 > S4 > S5 > S13 > S9 > S7 > S11 > S8 > S10. By studying the structure of the community, we found that in the plots S1, S9, S12 and S13, the community lineage structure was the aggregation pattern, and the species had a tendency to gather. In the other plots, the relationship between NRI and NTI was positive and negative, which led to the inability to determine whether the community lineage was aggregated or divergent. The functional richness of plot S2 was the lowest, and plot S1 was the highest. The functional evenness of sample S2 was the lowest, and plot S4 was the highest. Since Rao’s formula is not revised, the larger the value, the smaller the dispersion is, indicating that the likelihood of low dispersion of the plot S2 was greater. (2) PD showed significantly positive correlation with NRI (p = 0.045), negative correlation with NTI (p = 0.015), and positive correlation with species diversity index (p = 0.000). There was a significantly positive correlation between NRI and species diversity index (p = 0.021). There was no significant correlation between the other indices.
Keywords:Middle Reaches of the Yellow River, Wetland, Phylogenetic Diversity, Pedigree Structure, Functional Diversity
黄河中游湿地植物多样性特征研究
赵天梁
山西省林业调查规划院,山西 太原
收稿日期:2017年5月11日;录用日期:2017年5月26日;发布日期:2017年5月31日

摘 要
以黄河中游湿地植物为研究对象,构建了黄河中游湿地植物谱系树。采用谱系多样性指数(PD)、群落谱系结构指数(净谱系亲缘关系指数NRI和最近种间亲缘关系指数NTI)和7个功能多样性指数(功能丰富度指数FAD1、FDp、FDc、FRic;功能均匀度指数FEve;功能离散度指数Rao、FDis),分析了黄河中游湿地植物的谱系多样性、谱系结构与功能多样性,并计算了物种多样性指数(物种丰富度指数Patrick、物种均匀度指数Pielou和综合表示物种丰富度与物种均匀度的Shannon-Wiener以及Simpson指数)。结果表明:(1) 13个样地谱系多样性指数的范围在706.894~2289.170,PD值从大到小依次为:S1 > S12 > S3 > S6 > S2 > S4 > S5 > S13 > S9 > S7 > S11 > S8 > S10;样地S1、S9、S12和S13中,群落谱系结构为聚集模式,物种之间有聚集的趋势;其他样地中的NRI指数与NTI指数结果正负不一致;样地S2的功能丰富度最低,样地S1的功能丰富度最高,样地S2的功能均匀度最低,样地S4的功能均匀度最高,由于Rao的计算公式未修订,其值越大说明离散度越小,说明样地S2的功能离散度低的可能性较大。(2) PD与NRI、NTI、Patrick指数之间分别呈显著正相关(p = 0.045)、显著负相关(p = 0.015)和显著正相关(p = 0.000);NRI与Patrick指数之间呈显著正相关(p = 0.021),其余指数之间均无显著相关性。
关键词 :黄河中游,湿地,谱系多样性,谱系结构,功能多样性

Copyright © 2017 by author and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
生物多样性是描述群落构建和生态系统功能的重要指标,与群落和生态系统的构建、生产力和稳定性等生态过程密切相关 [1] ,一直是生态学研究的核心问题。生物多样性不仅仅是物种多样性,还包括系统发育多样性和功能多样性 [2] 。事实上,物种多样性甚至可能是所有这些生物多样性维度中信息最少的,与之相比,系统发育多样性和功能多样性更能揭示群落和生态系统的构建、维持与功能 [3] 。研究表明,具有相同物种多样性的不同区域,它们的系统发育多样性和功能多样性可以不同;具有不同物种多样性的区域之间,也可以具有相似的水平的系统发育多样性和功能多样性 [4] [5] [6] [7] 。近100年来,群落生态学的研究非常关注系统发育多样性,早期的研究分析了群落中物种和属的比率,了解生物与非生物的相互作用是否在群落构建中起重要作用 [8] [9] [10] 。具体来说,物种与属的比率低表示系统过度分散,物种与属的比率高则表示系统发育聚集。这种研究方法持续了几十年,同样盛极一时的还有群落构建规则和20世纪60、70年代的零模型辩论 [11] [12] [13] [14] 。物种与属比率方法的基础是假设密切相关的物种更可能具有类似的生态位 [15] [16] [17] 。如果密切相关的物种倾向于共享类似的生态位,则通过非生物过滤的群落构建应该导致系统发育聚集,而通过生物相互作用导致的群落构建应该造成系统发育过度分散 [18] [19] 。Charles Darwin考虑到共同后裔的影响,最先提出生态位保守主义。具体来说,拥有最近的共同祖先的物种平均来说往往比其他物种更相似。物种与属比率方法的问题超出了生态位保守主义的假设,属于分类学等级,由于两个物种偏离,不能传达关于时间的详细信息。解决这个问题方案是使用具有分支长度的系统发育树,分支长度可以用于提供对分类和系统发育之间的亲缘关系更精确的测量。直到21世纪初,人们普遍认为生成群落系统发育树是不可能的。2005年,Webb和他的同事解决了这一问题,他们开发了用于构建植物群落的系统发育树的软件工具Phylomatic,量化了群落的系统发育多样性,并探究了系统发育多样性与随机预期给定的物种多样性的高低 [20] 。这一发现引发了热带地区对于共存的植物物种间系统发育亲缘关系的大量研究 [21] 。功能多样性是指生物多样性中影响生态系统的过程及其功能的组分,具有较高功能多样性的生态系统往往具有较强的功能,其生产力、抗干扰能力和稳定性较强 [22] 。为了更深入地解释物种多样性对于生态系统过程与功能的真实影响,Mouillot等 [23] 提出将功能多样性的内涵划分为功能丰富度(functional richness)、功能均匀度(functional evenness)和功能离散度(functional divergence)三个维度。因此,计算功能多样性进行时,应采用相对有效的指数对每个维度进行评价,以综合衡量功能多样性的大小。
黄河是我国的第二大河流,也是我国河流湿地资源的重要组成部分。对于黄河中游湿地植物多样性的研究涉及到资源可持续利用 [24] 、景观格局变化 [25] 、分类学多样性 [26] 、植被数量生态 [27] 、植物区系 [28] 、优势种种间关系 [29] 等方面。有关黄河中游湿地植物谱系多样性和功能多样性方面的研究尚属空白。
本研究以黄河中游湿地为研究对象,对黄河中游湿地的植被谱系多样性进行分析,试图阐明以期阐明2个科学问题:(1)黄河中游湿地植物的谱系多样性、谱系结构和功能多样性如何?(2)黄河中游湿地植物的物种多样性、谱系多样性、群落谱系结构和功能多样性这些指数互相之间有着怎样的关系?本研究旨在补充、完善黄河中游湿地植物多样性的研究成果,为黄河中游湿地植物群落的构建机制提供理论依据。
2. 研究区概况与研究方法
2.1. 研究区概况
研究区位于黄河中游湿地从河津市禹门口至万荣县汾河入河口处的河道及沿岸湿地,地理坐标:35˚20'~35˚39'N,110˚27'~110˚36'E,属于暖温带大陆性季风气候,夏季高温多雨,冬季寒冷干燥;年平均气温12℃~13℃,1月平均气温−4℃,7月平均气温27℃。年平均降水量500~550 mm,无霜期180~200 d。
2.2. 样地设置与调查
2013年9月,以黄河中游龙门至汾河入黄口段湿地植被为研究对象,在全面踏查的基础上,尽量选取人为干扰较小的地方设置样方并对其进行植物生态学调查。样方设置具体方案为:在研究区域内沿黄河东岸且平行于黄河河流方向设置一条样带,并沿样带每隔1 km设置一个样地,避开农田与人为干扰的区域,共设样地13个(图1)。在每个样地内,根据样地现有植被类型的数量,采用典型取样与随机取样相结合的方法进行样方设置,每样地设置样方10~30个,其中草本植物样方面积为1 m × 1 m,灌木样方面积为5 m × 4 m;研究区域内共设置调查样方177个,其中灌木样方6个,草本样方171个。在植被调查中,主要记录样方内植物的种名、高度、盖度等,同时记录样带外植物种类,样带海拔、经纬度、生境特征、土壤类型及样地周围的人为干扰情况等。
2.3. 数据分析
2.3.1. 谱系树构建
应用Phylomatic在线进化树生成工具获得基于被子植物分类系统Ⅲ(APGⅢ)的进化树。
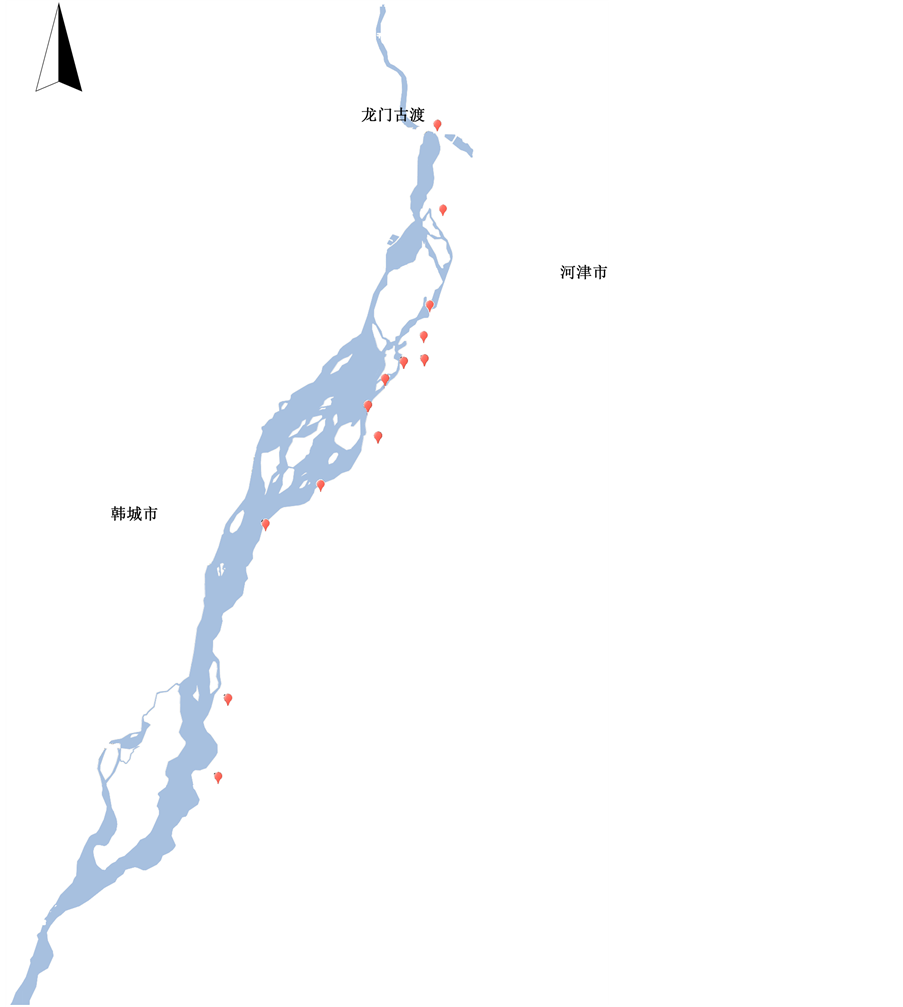
Figure 1. Location of the sample plots
图1. 样地位置示意图
2.3.2. 谱系指数
(1) 谱系多样性
利用Phylocom软件中的pd模块计算每块样地的谱系多样性(PD)。
(2) 群落谱系结构
将每个样地的植物视为一个群落,通过净谱系亲缘关系指数(net relatedness index, NRI)和最近种间亲缘关系指数(nearest taxon index, NTI)分析各样地的群落谱系结构。
NRI和NTI的计算公式如下 [30] :
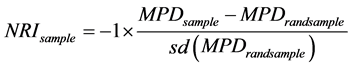 (1)
(1)
 (2)
(2)
上式中,NRIsample、NTIsample、MPDsample和MNTDsample代表群落中实际的观测值,MPDrandsample和MNTDrandsample代表物种在构建的谱系树上随机999次组合计算后获得的平均值,Sd为标准差。MPD (mean phylogenetic distance,种间平均进化距离)是群落中所有物种对间平均的进化距离;MNTD (mean nearest phylogenetic taxon distance,最近种间平均进化距离)则代表任意一个物种与群落中与自己亲缘关系最近的一个物种的平均进化距离。可以看出,它们的值越小,物种间的亲缘关系越近。
当NRI和NTI指数 > 0时,代表样地中的物种比从物种库中随机抽样组成的群落具有更高的谱系聚集度,表明群落结构趋向于亲缘相似的物种聚集在一起;当NRI和NTI指数 < 0时,代表物种间亲缘关系比从物种库中随机抽样组成的群落更分散,表明群落结构趋向于亲缘关系疏远的物种聚集在一起;当NRI和NTI指数 = 0时,表示群落谱系结构是随机的。可以看出,NRI量度所有物种整体的相似度,而NTI则更关心最相似的物种间的影响 [30] 。
2.3.3. 功能多样性
根据研究区的实际情况,本研究共选定了10种功能特征指标:种子扩散方式、传粉方式、固氮类型、生活史、生活型、植物叶型、盖度、株高、频度、始花期。为定量研究功能多样性,将研究方法所对应的指数分为功能丰富度、功能均匀度和功能离散度3个维度进行研究,本研究在功能多样性研究时,功能丰富度选取了FAD1、FDp、FDc、FRic 4个指数;功能均匀度选取FEve指数;功能离散度选取了Rao、FDis 2个指数,上述7个指数均采用FDiversity软件进行计算。
2.3.4. 物种多样性指数
采用的4个物种多样性指数是:反映物种丰富度的Patrick指数、反映物种均匀度的Pielou指数、综合反映物种丰富度及均匀度的Shannon-Wiener指数和Simpson指数。各指数的计算公式如下 [26] :
Patrick指数为:
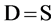 (3)
(3)
Shannon-Wiener指数为:
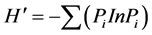 (4)
(4)
Pielou均匀度指数为:
 (5)
(5)
Simpson指数为:
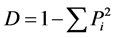 (6)
(6)
式中,S为每一样方的物种总数;Pi = Ni/N,其中Ni为第i个种的综合优势比指标,N为S个种的全部综合优势比之和。
2.3.5. 谱系多样性与传统多样性指数之间的关系
为进一步了解谱系多样性与传统多样性指数之间的关系,利用SPSS17.0对PD、NTI、NRI和物种多样性指数做相关性分析,并在此基础上对有显著相关关系的指数做回归分析。
3. 结果与分析
3.1. 黄河中游湿地植物谱系树构建
利用谱系软件包Phylocom 4.2,对研究区域的72个非被子植物物种构建了谱系树,并利用Figtree v1.4.3软件生成谱系树(图2)。我们在13个样地中调查所得的非被子植物物种数目分别为38、15、14、13、11、15、9、9、10、8、9、28、13 (表1)。
3.2. 黄河中游湿地植物谱系多样性
黄河中游湿地13个样地的谱系多样性如表1,ntaxa为物种数,PD是指包含且仅包含群落中物种的系统发育树形图中所有分枝长度之和。13个样地的谱系多样性指数的范围在706.894~2289.170,PD值从大到小依次为:S1 > S12 > S3 > S6 > S2 > S4 > S5 > S13 > S9 > S7 > S11 > S8 > S10。可以看出,物种数多的样地如S1、S2、S6和S12,它们的PD值也较大,而物种数少的样地如S7、S8、S10和S11,它们的PD值也较小。
3.3. 黄河中游湿地植物的群落谱系结构
在黄河中游湿地的13个样地中,存在不同的谱系结构(表2、表3)。样地S1、S9、S12和S13中,种间平均进化距离MPD分别是256.7111、244.7538、262.9594、264.491百万年,最近种间平均进化距离MNTD分别是80.4883、99.196、90.8059、92.5331百万年,MPD与MNTD小于随机谱系距离和随机种间最近谱系距离,即净谱系亲缘关系指数(net relatedness index, NRI)和最近种间亲缘关系指数(nearest taxon index, NTI) > 0,由此得出,样地S1、S9、S12和S13中,群落谱系结构为聚集模式,物种之间有聚集的趋势。其他的样地中,NRI指数与NTI指数结果正负不一致,无法判定群落谱系结构是聚集还是发散。
3.4. 黄河中游湿地植物的功能多样性
由7个功能多样性指数的计算公式可知,功能多样性的计算与物种数量和物种自身的功能特征有关。由表4可知,FAD1指数的变化范围为1.11~6.50;FDp指数的变化范围为3.391~16.370;FDc指数的变化范围为3.687~17.166,上述3个指数值的最大值和最小值分别对应样地S1和样地S2。FRic指数的变化范围为0.080~2.557,最大值和最小值分别对应样地S10和样地S13。FEve指数的变化范围为0.057~0.740,最大值和最小值分别对应样地S4和样地S2。Rao指数的变化范围为0.813~3.494,最大值和最小值分别对应样地S13和样地S2。FDis指数的变化范围为0.504~1.640,最大值和最小值分别对应样地S4和样地S2。
综合各个指数的定义以及上述计算结果,样地S2的功能丰富度最低,样地S1的功能丰富度最高,样地S2的功能均匀度最低,样地S4的功能均匀度最高,由于Rao的计算公式未修订,其值越大说明离散度越小,说明样地S2的功能离散度低的可能性较大。
3.5. 黄河中游湿地植物谱系多样性与传统多样性指数之间的关系
通过相关性分析可知,谱系多样性(PD)与净谱系亲缘关系指数(NRI)之间在0.05水平上呈显著正相关(r = 0.564, p = 0.045),谱系多样性(PD)与最近种间亲缘关系指数(NTI)之间在0.05水平上呈显著负相关(r = −0.656, p = 0.015),谱系多样性(PD)与物种丰富度(Patrick)之间在0.01水平上呈显著正相关(r = 0.982, p = 0.000),净谱系亲缘关系指数(NRI)与物种丰富度(Patrick)之间在0.05水平上呈显著正相关(r = 0.630, p = 0.021)。其余指数之间均无显著相关关系(p > 0.05)。

Figure 2. Summary of the phylogenetic tree assembled for the plant at the wetland of middle Yellow River
图2. 黄河中游湿地植物谱系树构建

Table 1. The phylogenetic diversity of the plant in the wetland of middle Yellow River
表1. 黄河中游湿地植物谱系多样性
注:S1:河津市禹门口河漫滩;S2:河津市黄河河道间滩地;S3:河津市汾河2号坝;S4:河津市汾河7号坝;S5:河津市连伯滩西;S6:河津市汾河9号坝;S7:河津市汾河14号坝;S8:河津市汾河25号坝;S9:闸西侧;S10:万荣县西范控导工程西侧;S11:万荣县上信王村西侧;S12:汾河入河口;S13:南谢控导工程南侧。下同。
Table 2. Mean phylogenetic distance (MPD), random phylogenetic distance (MPDrandsample), standard deviation of the MPDrandsample (sd.MPDrandsample) and the net relatedness index (NRl) of the plant in the wetland of middle Yellow River
表2. 黄河中游湿地植物种间平均进化距离、随机谱系距离、随机谱系距离标准偏差和净谱系亲缘关系指数
注:种间平均进化距离、随机谱系距离、随机谱系距离标准偏差的单位均为百万年。
Table 3. Mean nearest phylogenetic taxon distance (MNTD), random nearest phylogenetic taxon distance (MNTDrandsample), standard deviation of the MNTDrandsample (sd.MNTDrandsample) and the nearest taxon index (NTI) of the plant in the wetland of middle Yellow River
表3. 黄河中游湿地植物最近种间平均进化距离、随机种间最近谱系距离、随机种间最近谱系距离标准偏差和最近种间亲缘关系指数
注:最近种间平均进化距离、随机种间最近谱系距离、随机种间最近谱系距离标准偏差的单位均为百万年。
Table 4. Functional diversity index of 13 sample plots in the wetland of middle Yellow River
表4. 黄河中游湿地13个样地的功能多样性指数
黄河中游湿地植物物种多样性如表5,通过计算谱系多样性与传统多样性指数之间的关系(表6)可知,物种丰富度指数Patrick与净谱系亲缘关系指数(NRI)之间的相关性最强R2高达0.985,说明黄河中游湿地植物群落物种具有生态位保守性。二者之间的最优拟合函数为线性关系(y = 49.656x + 362.202),其余指数之间的相关性较弱(R2 < 0.7)。
4. 结论与讨论
生境过滤作用会导致群落内的物种适应能力相似、亲缘关系偏近,表现为谱系的聚集;相反,竞争排斥作用会导致群落内物种亲缘关系较远,表现为谱系的发散 [31] 。本研究中,样地S1、S9、S12和S13的物种之间有聚集的趋势,说明在这几个样地中以生境过滤作用为主导;其他样地内的NRI与NTI值正负不一致,这种情况与群落的健康稳定程度有关,群落中的脆弱物种如果消失,就会导致其生态系统功能下降 [32] 。
群落的构建会受到研究尺度、环境因子和人类活动的影响和控制。随研究尺度的变化, 群落聚集程度也随之发生改变 [33] 。在小尺度上,中国东北温带森林的群落构建中,竞争作用非常重要 [34] 。对黑龙江省的兴安落叶松的研究结果表明,在小空间尺度上以密度制约效应为主,谱系结构为发散;空间尺度越大,密度制约效应越弱,谱系结构则变为聚集 [35] 。对长白山植物的调查研究结果显示,海拔、林分郁闭度、坡度及土壤腐殖质厚度都会显著影响群落的谱系结构 [36] 。梅里雪山针阔混交林群落乔木层物种谱系结构于较低海拔呈现谱系聚集于较高海拔地区则为谱系发散 [37] 。不同采伐干扰方式、干扰程度与不同的放牧强度都会影响植物群落谱系结构 [38] [39] [40] 。
本研究着重从谱系多样性与功能多样性两个维度来研究黄河中游湿地植物的多样性,并分析了各多样性指数之间的相关性,发现功能多样性与各多样性指数之间无显著相关关系。其原因在于它们的定义与计算公式不同,各自所表征的生态学意义也不相同。湿地植物会受到湿地特殊的水文与土壤条件影响,本研究只是黄河中游湿地谱系多样性与群落构建研究的一个开端,在今后的研究中,还需加入环境因子与人为活动的研究,进一步揭示黄河中游湿地植物群落构建机制。
Table 5. Species diversity index of the plant in the wetland of middle Yellow River
表5. 黄河中游湿地植物物种多样性
Table 6. The regression of phylogenetic diversity index and species diversity index
表6. 黄河中游湿地植物谱系多样性与传统多样性指数之间的回归方程
基金项目
山西省基础研究计划项目(2014021032-3)。
文章引用
赵天梁. 黄河中游湿地植物多样性特征研究
Plant Diversity Characteristics of Wetland in the Middle Reaches of the Yellow River[J]. 世界生态学, 2017, 06(02): 112-123. http://dx.doi.org/10.12677/IJE.2017.62013
参考文献 (References)
- 1. Tilman, D., Reich, P.B., Knops, J., et al. (2001) Diversity and Productivity in a Long-Term Grassland Experiment. Science, 294, 843-845. https://doi.org/10.1126/science.1060391
- 2. Swenson, N.G. (2011) The Role of Evolutionary Processes in Producing Biodiversity Patterns, and the Interrelationships between Taxonomic, Functional and Phylogenetic Biodiversity. American Journal of Botany, 98, 472-480. https://doi.org/10.3732/ajb.1000289
- 3. 贾鹏, 杜国祯. 生态学的多样性指数: 功能与系统发育[J]. 生命科学, 2014, 26(2): 153-157.
- 4. Gray, A. (1846) Analogy between the Clora of Japan and That of the United States. American Journal of Sciences and Arts, 2, 175-176.
- 5. Wen, J. (2003) Evolution of Eastern Asian and Eastern North American Disjunct Distributions of Flowering Plants. Annual Review of Ecology and Systematics, 30, 421-455. https://doi.org/10.1146/annurev.ecolsys.30.1.421
- 6. Latham, R.E., Ricklefs, R.E., Ricklefs, R.E., et al. (1993) Continental Comparisons of Temperate-Zone Tree Species Diversity. In: Species Diversity in Ecological Communities: Historical and Geographical Perspectives, 294-314.
- 7. Guo, Q., Ricklefs, R.E. and Cody, M.L. (1998) Vascular Plant Diversity in Eastern Asia and North America: Historical and Ecological Explanations. Botanical Journal of the Linnean Society, 128, 123-136. https://doi.org/10.1111/j.1095-8339.1998.tb02111.x
- 8. Jaccard, P. (1926) Le coefficient generique et le coefficient de communaute dans la flore marocaine. Mémoires de la Société Vaudoise des Sciences Naturelles, 2, 385-403.
- 9. Jaccard, P. (1940) Coefficient générique réel et coefficient générique probable. Bulletin De La Societe Vaudoise Des Sciences Naturelles, 61, 117-136.
- 10. Elton, C. (1946) Competition and the Structure of Ecological Communities. Journal of Animal Ecology, 15, 54-68. https://doi.org/10.2307/1625
- 11. Simberloff, D.S. (1970) Taxonomic Diversity of Island Biotas. Evolution, 24, 23-47. https://doi.org/10.2307/2406712
- 12. Grant, P.R. and Abbott, I. (1980) Interspecific Competition, Island Biogeography and Null Hypotheses. Evolution, 34, 332-341. https://doi.org/10.2307/2407397
- 13. Harvey, P.H., Colwell, R.K., And, J.W.S., et al. (2003) Null Models in Ecology. Annual Review of Ecology and Systematics, 14, 189-211. https://doi.org/10.1146/annurev.es.14.110183.001201
- 14. Jarvinen, O. (1982) Species-to-Genus Ratios in Biogeography: A Historical Note. Journal of Biogeography, 9, 363- 370. https://doi.org/10.2307/2844723
- 15. Wiens, J.J. and Graham, C.H. (2005) Niche Conservatism: Integrating Evolution, Ecology and Conservation Biology. Annual Review of Ecology Evolution and Systematics, 36, 519-539. https://doi.org/10.1146/annurev.ecolsys.36.102803.095431
- 16. Peterson, A.T., Sobern, J. and Sanchez-Cordero, V. (1999) Conservatism of Ecological Niches in Evolutionary Time. Science, 285, 1265-1267. https://doi.org/10.1126/science.285.5431.1265
- 17. Wiens, J.J., Ackerly, D.D., Allen, A.P., et al. (2010) Niche Conservatism as an Emerging Principle in Ecology and Conservation Biology. Ecology Letters, 13, 1310-1314. https://doi.org/10.1111/j.1461-0248.2010.01515.x
- 18. Webb, C.O., Ackerly, D.D., McPeek, M.A., et al. (2002) Phylogenies and Community Ecology. Annual Review of Ecology and Systematics, 33, 475-505. https://doi.org/10.1146/annurev.ecolsys.33.010802.150448
- 19. Cavenderbares, J., Ackerly, D.D., Baum, D.A., et al. (2004) Phylogenetic over Dispersion in Floridian Oak Communities. The American Naturalist, 163, 823-843. https://doi.org/10.1086/386375
- 20. Webb, C.O. and Donoghue, M.J. (2005) Phylomatic: Tree Assembly for Applied Phylogenetics. Molecular Ecology Resources, 5, 181-183. https://doi.org/10.1111/j.1471-8286.2004.00829.x
- 21. Webb, C.O. (2000) Exploring the Phylogenetic Structure of Ecological Communities: An Example for Rain Forest Trees. The American Naturalist, 156, 145-155. https://doi.org/10.1086/303378
- 22. Petchey, O.L. and Gaston, K.J. (2006) Functional Diversity: Back to Basics and Looking Forward. Ecology Letters, 9, 741-758. https://doi.org/10.1111/j.1461-0248.2006.00924.x
- 23. Mouillot, D., Mason, W.H., Dumay, O., et al. (2005) Functional Regularity: A Neglected Aspect of Functional Diversity. Oecologia, 142, 353-359. https://doi.org/10.1007/s00442-004-1744-7
- 24. 上官铁梁, 宋伯为, 朱军, 等. 黄河中游湿地资源及可持续利用研究[J]. 干旱区资源与环境, 2005, 19(1): 7-13.
- 25. 郭东罡, 上官铁梁, 白中科, 等. 黄河中游连伯滩湿地景观格局变化[J]. 生态学报, 2011, 31(18): 5192-5198.
- 26. 李帅, 张婕, 上官铁梁, 等. 黄河中游湿地植物分类学多样性研究[J]. 植物科学学报, 2015, 33(6): 775-783.
- 27. 秦晓娟. 山西平陆黄河湿地植被数量生态研究[D]: [硕士学位论文]. 太原市: 山西大学, 2015.
- 28. 樊杰, 上官铁梁, 宋伯为. 黄河中游(禹门口-桃花峪)河漫滩种子植物区系地理研究[J]. 植物科学学报, 2003, 21(4): 332-338.
- 29. 李秋玲, 范庆安, 马晓勇, 等. 山西黄河湿地植被优势种群种间关系[J]. 生态学杂志, 2007, 26(10): 1516-1520.
- 30. 陈英. 常绿阔叶林谱系多样性对幼苗存活率的影响[J]. 植物生态学报, 2009, 33(6): 1084-1089.
- 31. 柴永福, 岳明. 植物群落构建机制研究进展[J]. 生态学报, 2016, 36(15): 4557-4572.
- 32. Mcwilliams, W., Moisen, G.G. and Czaplewski, R.L. (2009). Forest Inventory and Analysis (FIA) Symposium 2008, Park City, 21-23 October 2008, 56.
- 33. 黄建雄, 郑凤英, 米湘成. 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J]. 植物生态学报, 2010, 34(3): 309-315.
- 34. Zhang, J., Hao, Z., Song, B., et al. (2009) Fine-Scale Species Co-Occurrence Patterns in an Old-Growth Temperate Forest. Forest Ecology and Management, 257, 2115-2120.
- 35. 冷海楠, 崔福星, 张荣涛, 等. 兴安落叶松林植物群落谱系结构研究[J]. 黑龙江科学, 2014(12): 10-12.
- 36. 刘巍, 曹伟. 长白山植物群落谱系结构及环境因子对其的影响[J]. 干旱区资源与环境, 2013, 27(5): 63-68.
- 37. 李志. 梅里雪山针阔混交林沿海拔梯度的群落谱系结构、数量分类与排序[D]: [硕士学位论文]. 昆明市: 云南大学, 2015.
- 38. 卜文圣, 许涵, 臧润国, 等. 不同采伐干扰方式对热带山地雨林谱系结构的影响[J]. 林业科学, 2014, 50(4): 15-21.
- 39. 宋凯, 米湘成, 贾琪, 等. 不同程度人为干扰对古田山森林群落谱系结构的影响[J]. 生物多样性, 2011, 19(2): 190-196.
- 40. 王建勋. 基于不同放牧强度的高寒草原植物功能性状及群落谱系构建研究[D]: [博士学位论文]. 北京: 中国农业大学, 2016.