Botanical Research
Vol.
11
No.
02
(
2022
), Article ID:
49793
,
5
pages
10.12677/BR.2022.112023
生长素在拟南芥根尖干细胞中行驶两个相反的调控功能
王爽,周光蓥,张汉马*
重庆师范大学,生命科学学院,植物环境适应分子生物学重庆市重点实验室,重庆
收稿日期:2022年2月27日;录用日期:2022年3月21日;发布日期:2022年3月28日

摘要
生长素在拟南芥根尖干细胞调控中的作用是近年来植物学研究中的热点之一,受到了广泛的关注。现有实验证据表明生长素在拟南芥根尖干细胞调控中担负2个相反的功能,即既促进干细胞特性的维持,又促进干细胞的分化。生长素如何在同一细胞群中行驶这两个截然相反的调控功能的机理目前还不清楚。
关键词
拟南芥,生长素,根干细胞,PLT,mir160

Auxin Plays Two Opposite Regulatory Functions in Arabidopsis Root Meristem Stem Cells
Shuang Wang, Guangyin Zhou, Hanma Zhang*
Chongqing Key Laboratory of Molecular Adaptations of Plants, College of Life Sciences, Chongqing Normal University, Chongqing
Received: Feb. 27th, 2022; accepted: Mar. 21st, 2022; published: Mar. 28th, 2022

ABSTRACT
The regulatory role of auxin in Arabidopsis root meristem stem cells has been a hot spot of plant science research and has attracted considerable interest. Current experimental evidence shows that auxin has two opposite regulatory functions in Arabidopsis root meristems, i.e. promoting the maintenance of stem cell properties and also promoting differentiation of some stem cells. The mechanisms by which auxin executes those two regulations in the same group of cells are currently unknown.
Keywords:Arabidosis, Auxin, Root Stem Cells, PLT, mir160

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


生长素是一个非常重要的植物激素,其调控功能几乎涉及植物生命活动的各个方面 [1] [2]。由于其重要性,生长素的调控功能及其机理一直是植物学研究中的重点和热点,也是近年来植物学研究中进展最快的领域之一。经过过去20多年的研究,我们对生长素的生物合成、运输、信号传导机理以及在一些重要生长发育过程中的功能有了比较深入的了解 [1] [2] [3] [4] [5]。即便如此,目前仍然有不少生长素的调控细节尚待进一步研究,例如生长素在细胞水平的调控机理。本文将简要介绍一个我们认为没有被足够关注的与生长素细胞调控相关的问题:生长素在拟南芥根尖干细胞调控中的细胞调控机理。
在高等植物的根尖分生组织中含有一个干细胞团,它既是根器官持续生长发育的调控中心,同时也是器官发育过程中增量细胞的最终来源 [6]。在模式植物拟南芥的根尖分生组织中,干细胞团由静止中心细胞和围绕其周围的一圈组织特异性干细胞所构成(图1(a))。大量证据表明生长素在拟南芥根尖干细胞调控中担负极为重要的角色,但在细胞层面的作用机理目前并不清楚。现有实验证据显示生长素在拟南芥根尖干细胞调控中在细胞水平具有两个截然相反的调控功能,即既能促进干细胞特性的维持,又能促进干细胞分化(图1(b))。由于根尖干细胞是未分化的细胞,因此前一功能必然包含抑制干细胞分化,与后一功能在调控方向上截然相反。
生长素促进拟南芥根尖干细胞的作用最早由Sabatini等 [7] 提出。他们发现在拟南芥根尖分生组织中生长素浓度沿纵轴方向呈梯度分布,其浓度最大值所在的位置与根尖分生组织的干细胞区恰好重叠。进一步分析发现该生长素浓度的最大值对维持所覆盖区的细胞的干细胞特性,以及其远端(外侧)细胞的分裂模式与命运等有决定性的影响。改变生长素浓度最大值的浓度或位置会导致干细胞区和远端细胞区的细胞特性和分裂分化活动发生相应变化。上述结果被众多后续研究所证实 [3] [4] [8],因此生长素对根尖分生组织中干细胞的促进作用也逐步被广泛接受。Galinha等 [8] 进一步明确了生长素在促进根尖干细胞特性中的作用机理,提出生长素可能通过对PLETHORA (PLT)家族基因的表达调控来调控根尖干细胞和其它根尖分生组织中的细胞的分裂分化活动(图2(a))。PLT蛋白属于AP2-转录因子家族中的一个亚家族,在拟南芥中有4个成员 [9]。PLT蛋白在拟南芥根尖分生组织中呈现一个与生长素相类似的浓度梯度,其最大值也恰好与干细胞区重叠。PLT蛋白对根尖分生组织的细胞具有重要调控作用,且调控结果呈明显剂量效应。PLT1和PLT2功能同时缺失的突变体无法维持根尖分生组织(图2(b)),而PLT过量表达植株则具有比对照植株更长的根尖分生组织。最为关键的是生长素可诱导PLT基因的转录,且该诱导作用呈明显剂量效应 [8] [9]。基于上述研究结果,Galinha等提出在拟南芥根尖分生组织中生长素通过对PLT基因的转录调控实现对细胞活动(包括干细胞)的剂量调控假说(图2(a))。但该假说中的一些关键细节(如生长素对PLT基因转录调控的机理)仍然有待进一步验证,PLT蛋白对不同细胞(包括干细胞)的调控机理目前也不清楚。
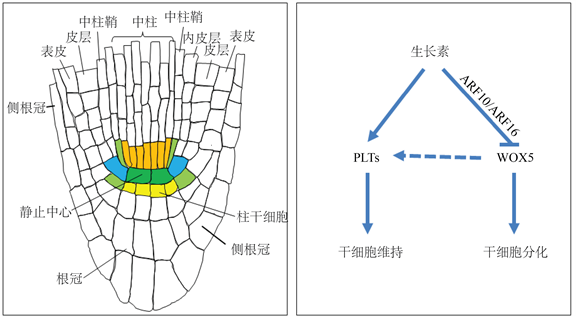 (a) (b)注:(a) 拟南芥根尖分生组织细胞结构示意图,颜色填充的细胞为干细胞;(b) 生长素在拟南芥根尖干细胞调控中的两种截然相反的调控功能。“↑”形线表示促进作用(正调控),“T”形表示抑制作用(负调控)。
(a) (b)注:(a) 拟南芥根尖分生组织细胞结构示意图,颜色填充的细胞为干细胞;(b) 生长素在拟南芥根尖干细胞调控中的两种截然相反的调控功能。“↑”形线表示促进作用(正调控),“T”形表示抑制作用(负调控)。
Figure 1. Schematic representation of the cellular structure of Arabidopsis root meristem (a) and the two opposite regulatory functions of auxin in stems cells of Arabidopsis root meristems (b)
图1. 拟南芥根尖分生组织细胞结构(a)和生长素在拟南芥根尖干细胞调控中的两种截然相反的调控功能(b)示意图
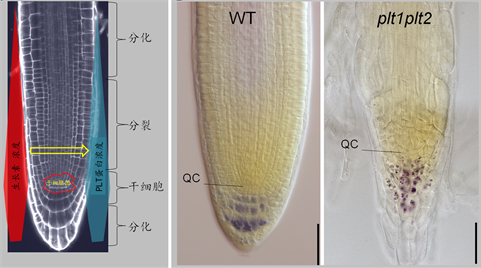 (a) (b) 注:(a) 生长素对拟南芥根尖干细胞正调控功能的作用机理假说,其依据来自文献 [7] [8] [9] 报道的实验结果。该假说的核心内容是生长素浓度在拟南芥分生组织中沿纵轴呈梯度分布,其最大值与干细胞区重叠。根尖分生组织中细胞的特性和分裂分化活动由生长素浓度所决定,生长素浓度最高值处为干细胞,中等生长素浓度区细胞快速分裂,低生长素浓度区细胞进行分化。生长素浓度梯度通过剂量调控的方式决定PLT基因的转录,由此转换成与生长素浓度梯度相类似的PLT蛋白浓度梯度,后者通过剂量调控决定细胞特性和分裂分化状况。梯度图形中以宽度代表浓度高低。(b) 野生型和plt1plt2根尖分生组织光学显微镜照片,显示plt1plt2突变体丧失了维持根尖分生组织的能力。照片拍摄于萌发后5天的幼苗,其中的标尺为10 µM。
(a) (b) 注:(a) 生长素对拟南芥根尖干细胞正调控功能的作用机理假说,其依据来自文献 [7] [8] [9] 报道的实验结果。该假说的核心内容是生长素浓度在拟南芥分生组织中沿纵轴呈梯度分布,其最大值与干细胞区重叠。根尖分生组织中细胞的特性和分裂分化活动由生长素浓度所决定,生长素浓度最高值处为干细胞,中等生长素浓度区细胞快速分裂,低生长素浓度区细胞进行分化。生长素浓度梯度通过剂量调控的方式决定PLT基因的转录,由此转换成与生长素浓度梯度相类似的PLT蛋白浓度梯度,后者通过剂量调控决定细胞特性和分裂分化状况。梯度图形中以宽度代表浓度高低。(b) 野生型和plt1plt2根尖分生组织光学显微镜照片,显示plt1plt2突变体丧失了维持根尖分生组织的能力。照片拍摄于萌发后5天的幼苗,其中的标尺为10 µM。
Figure 2. The positive regulatory function of auxin on in Arabidopsis root meristem stem cells
图2. 生长素对拟南芥根尖分生组织中干细胞的正调控功能
生长素对柱干细胞及其子细胞分化的促进作用也有充分的实验证据支持。证据之一来自于生长素响应突变体axr3-1。AXR3基因编码一个生长素信号响应元件IAA17。在axr3-1突变体中IAA17蛋白的II功能域中出现了一个氨基酸突变,导致该蛋白稳定性增加而不被生长素介导的降解系统所降解,由此导致突变体对生长素信号不敏感 [10]。axr3-1植株根尖分生组织中静止中心下方的细胞均未分化,不含淀粉粒 [7] [10] [11],表明由IAA17介导的生长素信号可促进柱干细胞及其子细胞分化。另一条支持证据来自中科院上海植物生理生态研究所陈晓亚院士领导的团队的研究结果 [12]。他们发现在过量表达mir160 (一个小RNA基因)的拟南芥植株或拟南芥arf10/arf16双基因功能缺失突变体植株中,根尖分生组织中静止中心下方细胞不分化(图3)。ARF10和ARF16为生长素信号响应通路中的核心元件,其编码基因是mir160的调控(降解)对象。上述结果同样表明生长素具有抑制柱干细胞及其后代细胞分化的功能。第3条证据来自丁兆军博士等(Ding & Friml, 2010)的研究结果 [11]。他们发现拟南芥根尖分生组织中柱干细胞的分化与细胞中生长素含量有直接关系,通过添加外源生长素NAA或生长素运输抑制剂NPA的方法增加根尖干细胞中生长素含量可促进柱干细胞的分化,而生长素合成或运输基因的功能缺失型突变体中在静止中心以下的未分化细胞层数增加。进一步分析他们还发现生长素可能作用与WOX5基因的上游,即通过下调WOX5基因的表达促进柱干细胞的分化。
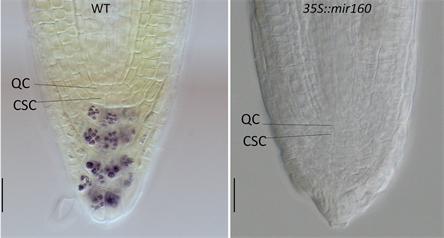 注:经Lugos染色的野生型(WT, Col-0)和35S:mir160幼苗(萌发后5天)的根尖分生组织光学照片。注意在野生型根尖分生组织中柱干细胞(CSC,静止中心下方的一层细胞)中不含淀粉粒(染色后呈棕褐色),但其下方的中央根冠细胞(又称柱细胞)中含淀粉粒;在35S:mir160根尖分生组织中,柱干细胞和中央根冠细胞都不含淀粉粒,表明没有分化。图中标尺为10 µm。
注:经Lugos染色的野生型(WT, Col-0)和35S:mir160幼苗(萌发后5天)的根尖分生组织光学照片。注意在野生型根尖分生组织中柱干细胞(CSC,静止中心下方的一层细胞)中不含淀粉粒(染色后呈棕褐色),但其下方的中央根冠细胞(又称柱细胞)中含淀粉粒;在35S:mir160根尖分生组织中,柱干细胞和中央根冠细胞都不含淀粉粒,表明没有分化。图中标尺为10 µm。
Figure 3. The differentiation promoting function of auxin in Arabidopsis root meristem
图3. 生长素在拟南芥根尖分生组织中对干细胞分化的促进作用
如上上述,有实验证据表明生长素在拟南芥根尖干细胞调控中承担了两个截然相反的细胞调控功能,但在文献中很少有人同时提及这两个功能。这可能是因为我们对它们的调控机理不是特别清楚,不太容易解释清楚生长素如何能行驶如此相反的功能。虽然目前对两个调控功能都提出了可能的作用机理,但这些机理在细胞水平的运作细节并不十分明确,有些解释甚至相互矛盾。例如Ding & Friml [11] 发现增加根尖干细胞中生长素浓度下调WOX5基因的转录,因此提出生长素作用于WOX5基因的上游,通过对后者转录的负调控促进干细胞的分化。可上述解释与生长素对干细胞特性的促进作用显然相互矛盾。此外Ding & Friml [11] 还发现WOX5的功能对PLT基因的表达具有正调控的作用,因此提出前者作用于PLT基因的上游,生长素通过下调WOX5基因降低PLT蛋白的浓度,由此促进干细胞分化。此结论与实验观察到的生长素对PLT基因表达的正调控作用也不符。总之,现有证据表明生长素在根尖干细胞中的确行驶两个相反的调控功能,但调控机理目前还不清楚,需要进一步研究。
基金项目
本研究得到了重庆师范大学人才基金(12XLR36)的资助。
文章引用
王 爽,周光蓥,张汉马. 生长素在拟南芥根尖干细胞中行驶两个相反的调控功能
Auxin Plays Two Opposite Regulatory Functions in Arabidopsis Root Meristem Stem Cells[J]. 植物学研究, 2022, 11(02): 192-196. https://doi.org/10.12677/BR.2022.112023
参考文献
- 1. Vanneste, S. and Friml, J. (2009) Auxin: A Trigger for Change in Plant Development. Cell, 136, 1005-1016. https://doi.org/10.1016/j.cell.2009.03.001
- 2. Chapman, E.J. and Estelle, M. (2009) Mechanism of Aux-in-Regulated Gene Expression in Plants. Annual Review of Genetics, 43, 265-285. https://doi.org/10.1146/annurev-genet-102108-134148
- 3. Petersson, S.V., et al. (2009) An Auxin Gradient and Maximum in the Arabidopsis Root Apex Shown by High-Resolution Cell-Specific Analysis of IAA Distribution and Synthesis. Plant Cell, 21, 1659-1668. https://doi.org/10.1105/tpc.109.066480
- 4. Blilou, I., et al. (2005) The PIN Auxin Efflux Facilitator Network Controls Growth and Patterning in Arabidopsis Roots. Nature, 433, 39-44. https://doi.org/10.1038/nature03184
- 5. Cheng, Y., Dai, X. and Zhao, Y. (2007) Auxin Synthesized by the YUCCA Flavin Monooxygenases Is Essential for Embryogenesis and Leaf Formation in Arabidopsis. Plant Cell, 19, 2430-2439. https://doi.org/10.1105/tpc.107.053009
- 6. Dolan, L., et al. (1993) Cellular Organisation of the Arabidopsis thaliana Root. Development, 119, 71-84. https://doi.org/10.1242/dev.119.1.71
- 7. Sabatini S, et al. (1999) An Auxin-Dependent Distal Organizer of Pattern and Polarity in the Arabidopsis Root. Cell, 99, 463-472. https://doi.org/10.1016/S0092-8674(00)81535-4
- 8. Galinha, C., et al. (2007) PLETHORA Proteins as Dose-Dependent Master Regulators of Arabidopsis Root Development. Nature, 449, 1053-1057. https://doi.org/10.1038/nature06206
- 9. Aida, M., et al. (2004) The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell, 119, 109-120. https://doi.org/10.1016/j.cell.2004.09.018
- 10. Rouse, D., et al. (1998) Changes in Auxin Response from Mutations in an AUX/IAA Gene. Science, 279, 1371-1373. https://doi.org/10.1126/science.279.5355.1371
- 11. Ding, Z. and Friml, J. (2010) Auxin Regulates Distal Stem Cell Differentiation in Arabidopsis Roots. PNAS, 107, 12046-12051. https://doi.org/10.1073/pnas.1000672107
- 12. Wang, J.W., et al. (2005) Control of Root Cap Formation by MicroRNA-Targeted Auxinresponse Factors in Arabidopsis. Plant Cell, 17, 2204-2216. https://doi.org/10.1105/tpc.105.033076
NOTES
*通讯作者。