Advances in Social Sciences
Vol.07 No.07(2018), Article ID:26001,12
pages
10.12677/ASS.2018.77146
The Influence of Positive Emotion on the Implicit Multi-Finality Choice: Evidence from ERPs
Ronghua Cai1,2*, Mengmeng Ren1,2, Wenjie Zhang1,2
1School of Educational Science, Hunan Normal University, Changsha Hunan
2Cognition and Human Behavior Key Laboratory of Hunan Province, Changsha Hunan

Received: Jun. 29th, 2018; accepted: Jul. 13th, 2018; published: Jul. 20th, 2018

ABSTRACT
In this study, Event related potential technology (ERP) was used to investigate the effect of positive emotion on implicit pleiotropic selection and its neural mechanism. The experiment uses the picture start paradigm, first presents the emotional picture, and then presents the choice stimulus. The task of the subjects is to carefully look at the stimulus and carry on the classification key response, and the requirement is fast and accurate. The results of this study showed that on P2, the selection of neutral conditions stimulated larger P2 amplitude than positive emotional conditions, and the positive start stimulation induced greater amplitude of P2 than the control stimulus. On N2, the neutral mood conditions stimulated smaller N2 amplitude than the selective processing under positive emotional conditions, and the positive picture stimulation induced smaller amplitude of N2 than the control stimulus. On P3, the neutral condition excites larger P3 amplitude than the positive emotional condition, and the positive picture processing under the neutral condition results in larger P3 wave amplitude. These findings may indicate that positive emotions may affect the processing of multiple validity choices. The two may have a cross processing system, and the implicit pleiotropic effect may show unstable characteristics once the positive emotion is affected.
Keywords:Event-Related Potential, Positive Emotion, Implicit Multi-Finality Choice, N2, P3
正性情绪对内隐多效性选择的影响:来自ERP的证据
蔡荣华1,2*,任梦梦1,2,张文洁1,2
1湖南师范大学,教育科学学院,湖南 长沙
2认知与人类行为湖南省重点实验室,湖南 长沙

收稿日期:2018年6月29日;录用日期:2018年7月13日;发布日期:2018年7月20日

摘 要
本研究采用事件相关电位技术(Event related potential technology, ERP),目的是考察正性情绪对内隐多效性选择的影响及其神经机制。实验使用图片启动范式,先呈现情绪图片,然后再呈现选择刺激,被试的任务是认真看刺激并进行分类按键反应,且要求又快又准。本研究的结果发现,在P2上,对比正性情绪条件,中性条件的选择激发了更大的P2波幅;积极启动刺激比对照刺激诱发了更大的P2波幅。在N2上,中性情绪条件比正性情绪条件下的选择加工激发了更小的N2波幅;在正性情绪启动条件下,积极图片刺激比对照刺激诱发了更小的N2的波幅。在P3上,中性条件比正性情绪条件下的选择激发了更大的P3波幅;对比正性条件,中性条件下的积极图片加工得到了更大的P3波幅。这些研究结果可能表明正性情绪会影响到多效性选择的加工,二者可能有着相互交叉的加工系统,而内隐多效性一旦受到正性情绪的影响可能会表现出不稳定的特征。
关键词 :事件相关电位,正性情绪,内隐多效性选择,N2,P3

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
人们所做出的选择可能被两种性质的目标驱动,一种是有意识的外显目标,另一种则是不被意识到的内隐目标 [1] 。无意识目标的激活已被证明影响了个体的认知、行为和情感 [2] [3] 。还有学者的研究表明人们的选择,可能是因为外显目标和内隐目标共同作用,同时这些目标都可能影响个体的选择,我们都知道在个体存在明确的外显目标的情况下,内隐目标同样可能影响个体的选择 [4] [5] ,这就是内隐多效性选择。总而言之,内隐多效性选择是指选择的目标可以带来多种效果,能同时满足意识到的外显需要和未意识到的内隐需要 [6] 。
情绪在决策过程的作用才得到越来越广泛的关注。通过对一些研究文献的回顾,在决策过程的神经机制研究中,研究者发现情绪会对个体的抉择产生非常重要的影响。例如,在决策行为的研究过程中,有研究者认为情绪和认知的脑部回路在其中扮演着重要的角色,他们的研究表明在不同的脑区分布着不同的认知与情绪,这些脑区的区域在不同阶段有不同的卷入程度 [7] 。还有来自关于后悔的研究,发现眶额皮层(OFC)整合了决策过程中的认知与情绪成分,并且它们自上而下地影响决策 [8] 。有研究者发现在对选项的情绪属性加工亦有重要作用的有来自杏仁核的影响 [9] ,他们在IGT (Iowa gambling task)任务中的研究发现杏仁核对决策的作用可能是因为它的损伤,直接导致了被试无法加工刺激的情绪属性,这样的研究从而就间接导致了决策的受损 [10] 。Miettinen等人的研究发现,人们只有在自己背叛,但是在别人合作的情况下才会体验到内疚情绪,并且如果博弈前双方已达成合作的承诺,单方背叛者的内疚程度会显著增加。另外,我们还发现,人们对单方背叛者的惩罚,会减轻背叛者自身的内疚感 [11] 。
这些与情绪相关的决策的研究忽略了社会选择中的感激、快乐、满足等正性情绪。而且很多的研究主要采用主观报告形式研究情绪,但是并没有考虑情绪的其他子成分,更没有把这些子成分当作为整体来研究了。再次是对情绪的神经机制方面的研究也偏少。对于社会选择类型的决策的情绪反应的神经机制研究非常地少,我们考察内隐多效性选择神经机制的研究更是一片空白,而且目前还没有严格操纵情绪以观察神经活动变化的研究,现在大多数是通过脑区激活逆推个体可能有的情绪反应,由于脑区与情绪并非严格地一一对应,所以这种逆推法本身就存在逻辑谬误。所以,探索正性情绪刺激与内隐多效性选择的关系更有意义,这有助于从内隐层面上了解选择与情绪的复杂关系,揭示不同情绪状态下内隐多效性选择的性质与特征。
2. 方法
2.1. 实验目的与实验假设
实验目的:采用颜色选择实验,探讨正性情绪对内隐多效性选择的影响及其神经机制。
实验假设:对比中性图片启动,正性情绪图片启动会增强被试内隐多效性选择。积极启动图片相比于控制图片在正性情绪诱导下会诱发更大N2和更小的P2和P3波幅。
2.2. 被试
我们选择了70名大学生参加了这个评定,对颜色与国家的相关程度进行了一个排序评定,评定的任务是对“红、橙、黄、绿、青、蓝、紫”七种颜色进行排序,与国家相关程度最高的排在最左边,与国家相关程度最低的排在最右边,从左至右相关程度依次递减。结果表明有84.29%的被试把红色排在了最左边,认为红色与国家的相关程度最高。我们将邀请30位将红色排在最左边的被试参加脑电实验。
实验收录30名在校大学生的脑电数据,这30位为将红色排在最左边的被试,身体健康无神经系统疾病。实验完成后付给一定报酬。
2.3. 实验材料
采用正性图片与中性图片做启动刺激,不同颜色的纯色图片作为选择行为的目标刺激。120张正性与中性图片(中国情绪图片系统,CAPS)组成启动刺激。实验时,将两种图片都重复使用三次,共计360张图片次,然后将正性情绪和中性图片平都均分配成3组,每组有120张图片次,包括取60张正性情绪图片次和60张中性情绪图片次。匹配好各组图片的效价强度、唤醒度和熟悉度,使其没有显著的差异。统计结果显示启动刺激平衡控制较好,3组正性图片之间的效价强度F(1, 59) = 0.31,p > 0.05,熟悉度F(1, 59) = 0.39,p > 0.05,和唤醒度F(1, 59) = 0.69,p > 0.05,都不存在显著的差异,三组中性图片之间的效价强度F(1, 59) = 0.27,p > 0.05,熟悉度F(1, 59) = 0.19,p > 0.05,和唤醒度F(1, 59) = 0.11,p > 0.05,也不存在显著的差异。
进行实验时,红色图片与其他图片配对呈现作为目标刺激,共计6种呈现方式,在实验中每种显现方式重复20次,平衡左右位置后,如“红、橙”,“橙、红”,共计呈现240次。除红色以外的其他六种颜色自由组合配对呈现作为对照刺激,共计15种呈现方式,在实验中每种显现方式重复2次,平衡左右位置,如“蓝、橙”,“橙、蓝”等,共计呈现120次。然后将360张图片分为三组,每组包含积极启动图片80张次和对照图片40张次。
2.4. 实验设计
实验为2 (情绪类型:正性情绪、中性) × 2 (图片类型:积极启动刺激、对照刺激)两因素被试内设计,因变量为行为数据(反应时)和ERP数据(P2、N2、P300波幅)。
2.5. 实验程序
实验开始前我们先对被试进行积极认同启动。在呈现配对的纯色图片刺激时,被试的任务是认真看刺激并进行分类按键反应,要求又快又准。如果被试认为左边的图片颜色更纯一些,则按“1”键;如果被试认为右边的图片颜色更纯一些,则按“2”键;练习实验之后,后跟360trails的正式实验(图1)。
2.6. 脑电记录
使用NeuroScan ERP记录与分析系统, 按国际10~20系统扩展的64导电极帽记录EEG。
2.7. ERP数据处理与统计
本研究完成连续记录EEG后,采用离线(off-line)处理数据。对不同相关程度的自我参照刺激呈现后的脑电数据进行分析与EEG叠加。我们的实验发现了明显的N1 (120~200 ms)、P2 (200~280 ms)、N2 (280~360 ms)、和P3 (500~700 ms)波幅,所以我们进行了四因素重复测量方差分析,2 (图片类型:积极启动刺激、对照刺激) × 2 (情绪效价:正、中性) × 3单侧化(左脑:FC3,C3,CP3;中脑:FCz,Cz,CPz;右脑:FC4,C4,CP4) × 3电极(头皮前中部:FC3,FCz,FC4;头皮中部:C3,Cz,C4;头皮后中部:CP3,CPz,CP4)。最后方差分析的P值采用Greenhouse-Geisser法校正。
3. 结果
3.1. 行为结果
对反应时进行重复测量方差分析,本研究结果表明情绪刺激与目标刺激存在着一个显著的交互作用,F(1, 29) = 5.03,p < 0.05, = 0.13;简单效应分析表明对比对照图片,被试对正性情绪图片条件下的积极启动图片的反应时间更快。
3.2. ERPs结果
见图2,在四类刺激中都发现了N1,P2,N2和P3成分。重复测量方差分析表明,N1在潜伏期和波幅上都没有发现显著的差异。
3.2.1. P2
使用重复测量方差分析在P2的波幅进行分析,情绪类型主效应显著,中性情绪条件比正性情绪条件下的选择加工激发了更大的P2波幅,F (1, 29) = 18.42,p < 0.001, = 0.49。重复测量方差分析还发现,情绪类型与电极位置交互作用显著,F (3, 87) = 14.35,p < 0.001, = 0.41。由此进行简单效应分析,在电极前中部,中部,中后部位置,中性情绪条件比正性情绪条件可以诱发的P2波幅有显著差异,all F(1,29) > 3.82,all p < 0.05。另外,图片类型与脑区位置交互作用显著,F(2,58) = 3.32,p < 0.05, = 0.17;简单效应分析表明,在右脑上积极启动刺激比对照刺激诱发了更大的P2波幅,F(1,29) = 3.56,p < 0.05。
3.2.2. N2
使用重复测量方差分析在N2的波幅进行分析,结果表明情绪类型主效应显著。相比于正性情绪条件,中性情绪条件选择加工激发更小的N2波幅,F (1, 29) = 4.12,p < 0.05, = 0.13。然后进行重复测量方差分析结果还显示,情绪类型与图片刺激交互作用显著,F (1, 29) = 4.98,p < 0.05, = 0.15。

Figure 1. Schematic representation in Experiment
图1. 实验中一个试次的实验流程
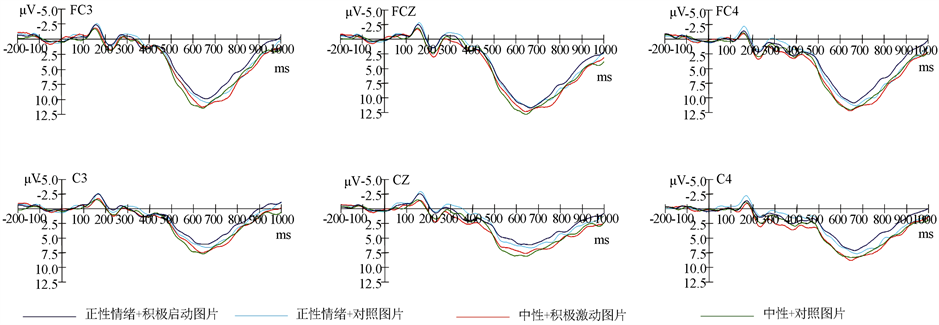
Figure 2. ERPs total mean map of two types of pictures at FC3, FCZ, FC4, C3, CZ and C4 electrodes initiated by positive and neutral emotions
图2. 正性与中性情绪启动下两类图片在FC3、FCZ、FC4、C3、CZ和C4电极点上诱发的ERPs总平均图
3.2.3. P3 (350~500 ms)
使用重复测量方差分析在P3平均波幅上进行分析,结果发现情绪类型主效应显著,相比于正性情绪条件,中性情绪条件的选择加工激发了更大的P3波幅,F (1, 29) = 10.36,p < 0.001, = 0.27。重复测量方差分析还发现,图片类型与脑区交互作用显著,F (2, 58) = 4.75,p < 0.05,
= 0.19。由此进行简单效应分析,结果表明在大脑中部和右部,相比于对照图片,积极启动图片所诱发的P3波幅有显著的差异,all F(1, 29) > 4.11,all p < 0.05。另外,图片类型与电极交互作用显著, F (2, 58) = 4.69,p < 0.05,
= 0.18。简单效应分析表明在大脑前中部和中部,积极启动图片所诱发的P3波幅显著高于对照图片,all F(1, 29) > 3.89,all p < 0.05。重复测量方差分析还显示,情绪类型与图片刺激交互作用显著,F (1, 29) = 4.37,p < 0.05,
= 0.14。简单效应分析表明对比正性图片,中性图片条件下的积极图片加工得到了更大的P3波幅。
= 0.27。重复测量方差分析还发现,图片类型与脑区交互作用显著,F (2, 58) = 4.75,p < 0.05,
= 0.19。由此进行简单效应分析,结果表明在大脑中部和右部,相比于对照图片,积极启动图片所诱发的P3波幅有显著的差异,all F(1, 29) > 4.11,all p < 0.05。另外,图片类型与电极交互作用显著, F (2, 58) = 4.69,p < 0.05,
= 0.18。简单效应分析表明在大脑前中部和中部,积极启动图片所诱发的P3波幅显著高于对照图片,all F(1, 29) > 3.89,all p < 0.05。重复测量方差分析还显示,情绪类型与图片刺激交互作用显著,F (1, 29) = 4.37,p < 0.05,
= 0.14。简单效应分析表明对比正性图片,中性图片条件下的积极图片加工得到了更大的P3波幅。
3.3. 溯源分析结果
将不同分类的总平均ERPs导入具有标准MRI头像的Curry6.0系统,用sLORETA方法在三壳球模型中重建各任务在不同时段的头皮活动源。根据实验2的ERPs成分,将正性和中性情绪诱导下积极启动减去对照图片所得差异波取P300 (350~500 ms)成分的数据进行定位分析(见图3)。
4. 讨论
目前的研究,其中的研究结果发现在N1的波幅上,四类选择加工没有发现显著的差异,这样的结果可能是由于对刺激类型的尺寸大小、长度和复杂性等物理属性上做到了均衡处理。在这一时间阶段,正性与中性情绪图片启动也没有影响到被试对多效性选择的加工,我们估计这种影响可能会出现在稍晚一点的阶段。

Figure 3. Traceability map of the differential wave P300 under the condition of positive priming and stimulation minus the contrast stimulus
图3. 实验中性与正性情绪条件下积极启动刺激减去对照刺激所得差异波P300溯源图
我们在刺激呈现后的研究结果,发现在额区电极观察到了明显的P2成分。中性图片条件下相对比正性图片多效性选择加工得到了更大的P2波幅。有研究表明,在额区位置发现的P2成分可能是对刺激特征的快速侦查,易敏感的刺激会得更多的关注 [12] [13] 。因此,我们的研究结果表明,中性图片条件下的多效性选择加工诱发更大P2,可能是由于正性图片呈现时,吸引了大量的注意资源,而人脑对中性图片的加工消耗的认知资源较小,对后面呈现的多效性选择加工影响也较小,积极启动刺激得到了充分的加工。在中性图片条件下,对积极启动刺激的加工得到了更大的P2波幅。这可能是由于对被试积极启动后,积极启动刺激的呈现会诱发被试的正性情绪,从而吸引了大量的注意资源。而人脑对控制图片的加工消耗的认知资源较小,所以诱发的P2波幅也更小。这一阶段发现差异可能与多效性选择行为无关,这是因为人脑对多效性刺激的选择与决策是晚期加工,可能发生在稍晚的阶段。
在额区位置的电极还发现了比较明显的N2成分。对比中性,正性情绪条件下的多效性选择加工诱发了更大N2波幅。这可能是由于正性情绪图片的出现从而吸引了大量的注意资源,从而导致人脑对随后出现的多效性选择加工的认知资源不足,加工困难变大,所以诱发的更大N2波幅。对比控制刺激,积极启动刺激诱发了更小的N2波幅。另外,还有研究表明,病态赌博成瘾者在决策任务中比控制组被试诱发了更小的N2 [14] 。因此,这一阶段发现差异也可能与多效性选择行为有关,人脑对多效性刺激的选择与决策有可能在这一阶段就开始了。
我们的研究发现,在两种情绪图片启动下,内隐多效性选择加工都能观察到明显的P3成分。对比正性图片,中性图片条件下的多效性选择加工得到了更大的P3波幅。因此,中性图片条件下的多效性选择加工诱发更大P3,也可能是由于正性图片呈现时,吸引了大量的注意资源,而人脑对中性图片的加工消耗的认知资源较小,对后面呈现的多效性选择加工影响也较小,积极启动刺激得到了充分的加工。另外,研究结果还表明,无论在正性情绪图片还是中性图片的影响下,多效性选择加工的优势效应都不明显。这可能是因为内隐多效性选择行为容易受到情境的干扰,一旦受到影响可能会表现出了不稳定的特征。
本研究的结果还发现了,本研究的目的:明显的内隐多效性选择效应,相比于控制刺激,积极启动刺激会诱发了更大的P2和P3波幅,本研究这个效应在中部和中后部脑区最为明显。而且偶极子溯源结果表明积极启动刺激减去对照刺激所得差异波P300成分显著激活了额上回和梭状回。这些实验结果与前人的研究相似,前人的研究发现扣带前回(ACC),腹内侧前额皮质(vmPFC),眶额皮层(OFC)等都可能是个体决策的神经机制 [15] [16] 。这种额脑区的优势效应可能表明了额区是加工正性情绪启动下多效性刺激的核心和关键区域。这也为进一步的研究提供了一个方向,未来的研究可以通过具有精细定位功能的fMRI技术来探讨这一部分脑区是不是内隐多效性选择加工的关键的神经机制。
5. 结语
总而言之,我们的研究是借助高时间进程的精密仪器ERPs,这样的研究重复了内隐多效性选择行为效应,并且拓展了前人的研究,这个研究的结果揭示了正性情绪对多效性选择加工程度效应的影响。正性情绪会影响到多效性选择的加工,二者可能有着相互交叉的加工系统。内隐多效性选择行为容易受到干扰,一旦受到影响可能会表现出不稳定的特征。
基金项目
教育部人文社会科学研究规划基金项目(13YJA190001)、湖南省自然科学基金项目(2015JJ2097, 2018JJ2254)、湖南省社会科学成果评审委员会重点课题(XSP17ZDI019)、湖南省哲学社会科学基金青年资助项目(16YBQ047)、湖南省教科规划青年资助项目(XJK016QXL002)和湖南省博物馆新馆儿童教育合作项目(20131231)。
文章引用
蔡荣华,任梦梦,张文洁. 正性情绪对内隐多效性选择的影响:来自ERP的证据
The Influence of Positive Emotion on the Implicit Multi-Finality Choice: Evidence from ERPs[J]. 社会科学前沿, 2018, 07(07): 968-979. https://doi.org/10.12677/ASS.2018.77146
参考文献
- 1. Bagozzi, R.P. and Dholakia, U. (1999) Goal Setting and Goal Striving in Consumer Behavior. The Journal of Marketing, 63, 19-32. https://doi.org/10.2307/1252098
- 2. Fitzsimons, G.M. and Bargh, J.A. (2003) Thinking of You: Nonconscious Pursuit of Interpersonal Goals Associated with Relationship Partners. Journal of Personality and Social Psychology, 84, 148. https://doi.org/10.1037/0022-3514.84.1.148
- 3. Kruglanski, A.W., Shah, J.Y., Fishbach, A., Friedman, R., Chun, W.Y. and Sleeth-Keppler, D. (2002) A Theory of Goal Systems. Advances in Experimental Social Psychology, 34, 331-378. https://doi.org/10.1016/S0065-2601(02)80008-9
- 4. Chun, W.Y. and Kruglanski, A.W. (2005) Consumption as a Mul-tiple-Goal Pursuit without Awareness. Applying Social Cognition to Consumer-Focused Strategy, 201-217.
- 5. Simonson, I. (2005) In Defense of Consciousness: The Role of Conscious and Unconscious Inputs in Consumer Choice. Stanford GSB Working Paper No. 1883.
- 6. 蔡荣华, 钟毅平. 外显与内隐选择的多效性原则[J]. 心理科学, 2014, 37(1): 66-72.
- 7. Ernst, M. and Paulus, M.P. (2005) Neurobiology of Decision Making: A Selective Review from a Neurocognitive and Clinical Perspective. Biological Psychiatry, 58, 597-604. https://doi.org/10.1016/j.biopsych.2005.06.004
- 8. Coricelli, G. and Nagel, R. (2009) Neural Correlates of Depth of Strategic Reasoning in Medial Prefrontal Cortex. Proceedings of the National Academy of Sciences, 106, 9163-9168. https://doi.org/10.1073/pnas.0807721106
- 9. Breiter, H.C., Aharon, I., Kahneman, D., Dale, A. and Shizgal, P. (2001) Functional Imaging of Neural Responses to Expectancy and Experience of Monetary Gains and Losses. Neuron, 30, 619-639. https://doi.org/10.1016/S0896-6273(01)00303-8
- 10. Bechara, A., Damasio, H., Damasio, A.R. and Lee, G.P. (1999) Different Contributions of the Human Amygdala and Ventromedial Prefrontal Cortex to Decision-Making. The Journal of Neuroscience, 19, 5473-5481. https://doi.org/10.1523/JNEUROSCI.19-13-05473.1999
- 11. Miettinen, T. and Suetens, S. (2008) Communication and Guilt in a Prisoner’s Dilemma. Journal of Conflict Resolution, 52, 945-960. https://doi.org/10.1177/0022002708322325
- 12. Karayanidis, F. and Michie, P. (1996) Frontal Processing Negativity in a Visual Selective Attention Task. Electroence-phalography and Clinical Neurophysiology, 99, 38-56. https://doi.org/10.1016/0921-884X(96)95116-4
- 13. Thorpe, S., Fize, D. and Marlot, C. (1996) Speed of Processing in the Human Visual System. Nature, 381, 520-522.
- 14. Torres, A., Catena, A., Megías, A., Maldonado, A., Cándido, A., Verdejo-García, A. and Perales, J.C. (2013) Emotional and Non-Emotional Pathways to Impulsive Behavior and Addiction. Frontiers in Human Neuroscience, 7. https://doi.org/10.3389/fnhum.2013.00043
- 15. Bechara, A., Damasio, H. and Damasio, A.R. (2003) Role of the Amygdala in Decision-Making. Annals of the New York Academy of Sciences, 985, 356-369. https://doi.org/10.1111/j.1749-6632.2003.tb07094.x
- 16. Kenning, P. and Plassmann, H. (2005) NeuroEconomics: An Overview from an Economic Perspective. Brain Research Bulletin, 67, 343-354. https://doi.org/10.1016/j.brainresbull.2005.07.006
附件
部分实验材料
1、纯色图片(部分)






2、情绪图片(部分)




NOTES
*通讯作者。