Botanical Research
Vol.
10
No.
04
(
2021
), Article ID:
44254
,
11
pages
10.12677/BR.2021.104072
海水胁迫下黄秋葵Na+、K+含量变化规律研究
刘岩1,程玉静2,翟彩娇2,王小秋2,刘畅1,谢冬微1,孙健1*,戴志刚3*
1南通大学生命科学学院,江苏 南通
2江苏沿江地区农业科学研究所,江苏 南通
3中国农业科学院麻类研究所,湖南 长沙

收稿日期:2021年6月18日;录用日期:2021年7月22日;发布日期:2021年7月29日

摘要
黄秋葵(Abelmoschus esculentus L.)为一年生草本植物,具有很高的营养价值。我国有大面积沿海滩涂盐渍化土地,但有关海水胁迫对黄秋葵体内Na+和K+含量变化规律的研究较少。本研究以耐盐黄秋葵品种无角香蕉和盐敏感品种非洲秋葵为材料,采用50%和100%浓度的海水进行胁迫处理,于第0、4、8、12 d进行取样,分析Na+和K+含量在黄秋葵体内的变化规律。结果表明,海水胁迫后,黄秋葵体内Na+含量和Na+/K+呈逐渐增加的趋势,K+含量呈逐渐下降的趋势。耐盐品种无角香蕉在海水胁迫后,根中积累了较多的Na+,而茎和叶中的Na+含量则明显低于非洲秋葵;无角香蕉茎和叶中K+含量的降低幅度明显小于非洲秋葵;无角香蕉根的Na+/K+高于非洲秋葵,而茎和叶的Na+/K+则明显低于非洲秋葵。耐盐品种无角香蕉在受到海水胁迫后根系对Na+具有较强的存储功能,叶片则具有较强的Na+外排功能;无角香蕉能较好地控制茎和叶中的K+含量,并维持其占整株比率的相对稳定。本研究为明确黄秋葵的海水盐胁迫耐受机理以及耐盐黄秋葵新品种选育提供了理论依据。
关键词
黄秋葵,海水胁迫,Na+含量,K+含量

Study of the Change Regulation of Na+ and K+ Contents in Okra under Seawater Stress
Yan Liu1, Yujing Cheng2, Caijiao Zhai2, Xiaoqiu Wang2, Chang Liu1, Dongwei Xie1, Jian Sun1*, Zhigang Dai3*
1School of Life Sciences, Nantong University, Nantong Jiangsu
2Jiangsu Yanjiang Institute of Agricultural Sciences, Nantong Jiangsu
3Institute of Bast Fiber Crops, Chinese Academy of Agricultural Sciences, Changsha Hunan

Received: Jun. 18th, 2021; accepted: Jul. 22nd, 2021; published: Jul. 29th, 2021

ABSTRACT
Okra (Abelmoschus esculentus L.) is an annual herb with high nutritional value. There is a large area of salinized land along the beach in China, but there are few studies on the changes of Na+ and K+ contents in okra under seawater stress. In this study, 50% and 100% concentrations of seawater were used to treat the salt-tolerant okra cultivar Wujiaoxiangjiao and salt-sensitive cultivar Feizhouqiukui. Samples were taken at 0, 4, 8 and 12 days to analyze the changes of Na+ and K+ contents in okra. The results showed that after seawater stress, Na+ content and Na+/K+ in okra increased gradually, while K+ content decreased gradually. After seawater stress, more Na+ was accumulated in the root of salt-tolerant cultivar Wujiaoxiangjiao, while the content of Na+ in the stem and leaf was significantly lower than that of Feizhouqiukui. The decrease of K+ content in the stem and leaf of Wujiaoxiangjiao was less than that of Feizhouqiukui. The Na+/K+ in the root of Wujiaoxiangjiao was higher than that of Feizhouqiukui, while the Na+/K+ of stem and leaf was lower than that of Feizhouqiukui. After seawater stress, the root of salt-tolerant cultivar Wujiaoxiangjiao has a strong storage function for Na+, while the leaf has a strong Na+ efflux function. The content of K+ in stem and leaf of Wujiaoxiangjiao can be well controlled, and the proportion of K+ in the whole plant is relatively stable. This study provides a theoretical basis for understanding the mechanism of salt tolerance of okra and the breeding of new salt-tolerant okra varieties.
Keywords:Okra, Seawater Stress, Na+ Content, K+ Content

Copyright © 2021 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
土壤盐渍化是限制植物生长发育和产量增长的重要环境因素。中国盐渍化土地面积已经超过1亿公顷,致使耕地不足,农业生产力下降 [1]。海洋植物在长期的进化过程中,产生了对高盐的耐受性,而大部分陆地植物则无法适应土壤中的盐分而只能在甜土中生活 [2]。土壤中的盐分主要以两种方式胁迫植物,一是盐分中过量的Na+和Cl−会对植物造成毒害,导致生理生化代谢紊乱,进而对形态建成和生长发育造成不良影响;二是使根系难以吸取水分,引起渗透胁迫导致膜透性改变 [3] [4]。
盐胁迫环境下,植物细胞中积累的过量Na+会打破细胞的离子平衡,特别是Na+与K+之间的平衡。K+是细胞中60多种酶的活化剂,在蛋白质和糖类的合成与运输以及细胞渗透势调节等方面发挥着重要作用 [5]。Na+大量进入细胞,竞争执行新陈代谢所必需酶的K+结合位点,引起K+外渗,同时抑制K+及其它离子的吸收,严重影响植物细胞的正常代谢和生长 [6]。为了适应盐环境,植物需通过Na+外排或区隔化以降低细胞内的Na+浓度,进而建立新的离子稳态 [7]。耐盐基因型除了需要在盐环境条件中保持低浓度的Na+外,低水平的Na+/K+对其维持离子平衡和较高的生物量也极为重要,因此,低Na+含量和Na+/K+被认为是耐盐品种筛选的重要指标 [8]。王学征等 [9] 在研究番茄耐盐性时表明,耐盐番茄品种体内的Na+总积累量低于不耐盐品种,且Na+/K+也低于不耐盐品种。姜静涵等 [10] 对不同耐盐性的大豆品种进行了苗期盐胁迫处理,结果表明,盐胁迫使盐敏感品种叶片的Na+含量显著升高,且随着盐浓度的增加,耐盐品种叶片Na+含量显著低于盐敏感品种,并建立了以Na+含量为核心的简便大豆耐盐性鉴定方法。王旭明等 [11] 研究不同水稻品种的耐盐性结果表明,耐盐品种根系的聚Na+能力较强,可以缓解细胞渗透压,减少盐分对地上部的伤害。耐盐品种叶片K+积累量随盐浓度的增加而逐渐升高,以置换出过多的Na+,维持叶细胞的Na+、K+平衡,保持正常的叶片功能。
黄秋葵(Abelmoschus esculentus L.),又称秋葵、补肾草、咖啡葵、羊角豆等,原产地为非洲,是药食两用型植物 [12]。黄秋葵在美国被称为植物伟哥,在印度被称为神奇的乡土菜,日韩称之为绿色人参,具有很高的营养保健价值 [13] [14]。黄秋葵果实中富含类黄酮、多糖、脂肪酸、维生素、矿质元素和多种氨基酸等功能营养保健成分,是老幼皆宜的蔬菜 [15]。目前,关于黄秋葵的研究主要集中在其营养品质的开发和利用以及栽培技术等方面,逆境特别是盐胁迫下黄秋葵的研究相对较少。王永慧等 [16] [17] [18] 研究了NaCl盐胁迫对黄秋葵种子萌发期和幼苗期生长发育及生理特性的影响。刘雅辉等 [19] 对黄秋葵苗期的盐胁迫反应及耐盐阈值进行了分析,认为黄秋葵耐盐性较强,适宜在轻、中度盐碱土地区种植。中国有漫长的海岸线,沿海滩涂面积约217万hm2,滩涂盐渍土壤以复盐为主,以往用NaCl或Na2CO3模拟盐胁迫来研究盐分对植物的影响会与实际有一定的差距 [20]。本研究采用海水处理耐盐性不同的黄秋葵品种,分析海水胁迫下黄秋葵体内Na+、K+含量的变化规律,探讨黄秋葵的海水盐胁迫耐受机理,旨在为耐盐黄秋葵新品种选育及开发利用沿海滩涂地区盐渍土地提供参考。
2. 材料与方法
2.1. 试验材料
耐盐型黄秋葵品种无角香蕉和盐敏感型品种非洲秋葵,由中国农业科学院麻类研究所(中国农业科学院南方经济作物研究中心)提供。海水取自江苏省南通市东灶港镇,盐度为3.12%。
2.2. 方法
2.2.1. 黄秋葵幼苗的培养
挑选大小一致、饱满的黄秋葵种子,用浓度为3%的次氯酸钠消毒10 min,再用蒸馏水冲洗3次。将黄秋葵种子播于装有营养土的育苗盘内,育苗盘的规格为52 cm × 26 cm × 10 cm,每个育苗盘含50穴,穴的规格为5 cm × 5 cm。每个品种播3盘,每穴播3粒种子,待子叶展平后间苗,每穴留1棵苗。黄秋葵幼苗在温室内培养,待苗长至两叶一心时进行海水处理。
2.2.2. 海水胁迫处理
设置50% (T1)和100% (T2) 2个海水处理浓度,自来水浇灌为对照(CK)。开始处理时T1、T2的浓度均为50%,第2天T2的浓度提高为100%,以防止幼苗突然遭受高浓度盐胁迫而难以适应。
2.2.3. 测定指标及方法
分别于处理的第0、4、8、12 d进行性状测定,包括苗高、Na+和K+含量,三次重复。
测量苗高时每个处理随机选取5颗苗,挂牌标记,苗高为茎基部到茎顶端的距离。
每次取样时每个处理选取3棵苗,缓慢从育苗盘中带土拔出,用自来水将根部冲洗干净,尽量保证根系完整,再用蒸馏水冲洗,吸水纸吸取根系表面水分。将苗的根、茎、叶剪开,迅速置于105℃烘箱中杀青,80℃烘干至恒重。将样品磨成粉末,过100目筛,准确称取0.1000 g,放入具塞试管内,加入10 ml浓度为100 mmol/L的乙酸在90℃条件下提取2 h [21],用火焰光度计测定Na+和K+含量。Na+含量和K+含量的比值为Na+/K+。
调查T1和T2从开始海水处理到植株全部死亡的天数,计为存活天数。
2.3. 数据分析
用Microsoft Excel 2019进行数据整理,SPSS 21.0软件进行数据统计分析。
3. 结果与分析
3.1. 黄秋葵品种耐盐比较
表1为不同浓度海水处理和对照条件下黄秋葵苗高的变化规律和海水处理后黄秋葵的存活天数。从表1可以看出,开始进行海水处理时,无角香蕉和非洲秋葵的苗高接近,无角香蕉略高于非洲秋葵。对照条件下,两个品种的苗高生长速度均较快,无角香蕉从第0 d到第12 d的苗高增长量为6.42 cm,非洲秋葵为5.72 cm,无角香蕉比非洲秋葵的增长量多0.70 cm。在50%海水处理(T1)条件下,两品种的苗高生长开始放缓,两品种处理后第4 d的苗高与0 d相比差异均不显著。无角香蕉从第0 d到第12 d的苗高增长量为3.94 cm,非洲秋葵为2.22 cm,无角香蕉比非洲秋葵的增长量多1.72 cm。在100%海水处理(T2)条件下,两品种的苗高生长受到严重抑制,增长极为缓慢,且不同天数的数值差异大多不显著。无角香蕉从第0 d到第12 d的苗高仅增长2.02 cm,非洲秋葵仅为0.96 cm,无角香蕉比非洲秋葵的苗高增长量多1.06 cm。
无角香蕉从开始50%海水处理后存活了47天,非洲秋葵存活了33天,无角香蕉比非洲秋葵多存活14天。无角香蕉从开始100%海水处理后存活了18天,非洲秋葵仅存活了15天,无角香蕉比非洲秋葵多存活3天。从苗高的变化和存活天数可以看出,无角香蕉在海水处理后比非洲秋葵具有较多的苗高增长量和存活天数,说明无角香蕉比非洲秋葵具有较好的海水耐受能力,特别是在50%海水处理条件下,这种耐受能力的差异更大。

Table 1. Comparison of seedling height and survival days between two okra cultivars under the seawater stress
表1. 海水胁迫下两个黄秋葵品种苗高和存活天数的比较
同列数字不同字母表示处理间在P < 0.05水平差异显著。
3.2. 海水胁迫对黄秋葵体内Na+含量的影响
为了分析黄秋葵幼苗在海水处理后体内Na+和K+含量的变化规律,在海水处理的第0、4、8、12 d对黄秋葵根、茎、叶的Na+和K+含量进行测定,结果如图1所示。黄秋葵受到海水胁迫后,体内Na+含量呈逐渐增加的趋势,各器官Na+含量总趋势为根 > 茎 > 叶。50%海水处理的Na+含量相比对照迅速增加,100%海水处理的Na+含量最高。从处理时间上来看,Na+含量在第4 d和8 d均有较大增长,第12 d含量达到最高值,但第12 d的增长量与第8 d相比,差距已经很小。耐盐品种无角香蕉在海水胁迫后,根中积累了较多的Na+,盐敏感品种非洲秋葵的积累量相对较少。而茎和叶的趋势则相反,无角香蕉茎和叶中的Na+含量明显低于非洲秋葵,这种差异在50%海水处理时表现的比较明显。
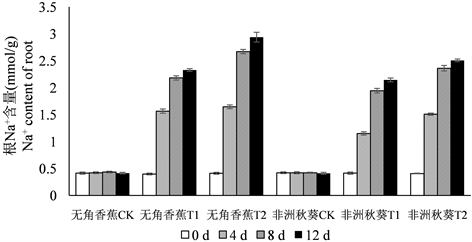
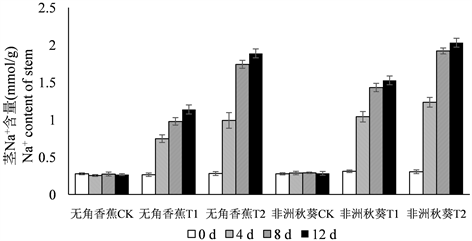
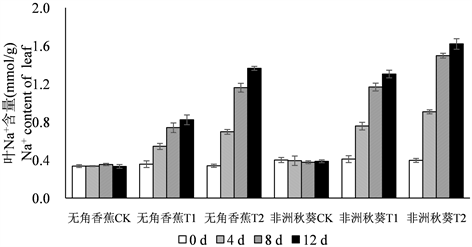
Figure 1. Dynamic change of Na+ content in root, shoot, leaf of Wujiaoxiangjiao and Feizhouqiukuiokra under seawater stress
图1. 海水胁迫下黄秋葵无角香蕉和非洲秋葵根、茎、叶中Na+含量的动态变化
3.3. 海水胁迫对黄秋葵体内K+含量的影响
海水胁迫下,黄秋葵根、茎、叶中的K+含量呈逐渐下降的趋势,K+含量总的趋势为根 > 茎 > 叶(图2)。各器官50%海水处理的K+含量与对照相比下降幅度较大,100%浓度下继续下降,但下降幅度明显放缓。在处理时间上,两个品种大部分处理的K+含量在第4 d做出迅速下降的反应,第8 d持续下降,第12天降幅减小。无角香蕉与非洲秋葵相比,根K+含量在处理后各时间点的变化没有明显的规律,可见根K+含量并不能反映出不同黄秋葵品种的耐盐性。海水处理后,无角香蕉茎和叶中K+含量的降低幅度明显小于非洲秋葵,说明海水胁迫对无角香蕉地上部K+含量的影响相对较小。
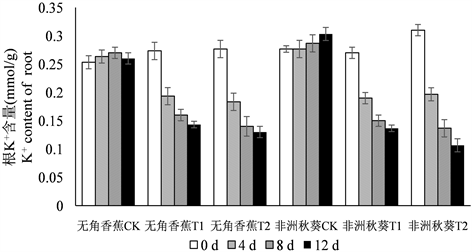

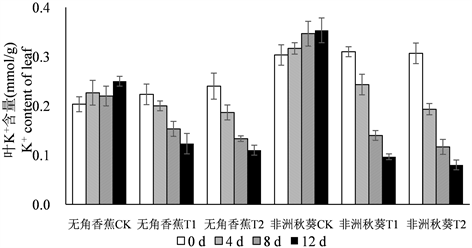
Figure 2. Dynamic change of K+ content in root, shoot, leaf of Wujiaoxiangjiao and Feizhouqiukuiokra under seawater stress
图2. 海水胁迫下黄秋葵无角香蕉和非洲秋葵根、茎、叶中K+含量的动态变化
3.4. 海水胁迫对黄秋葵体内Na+/K+的影响
海水胁迫下,由于黄秋葵根、茎、叶中的Na+含量上升,K+含量下降,因此,Na+/K+呈大幅度升高趋势(图3)。由于50%浓度海水处理的Na+含量比100%浓度低,而K+含量高,因此,两个品种各器官50%浓度海水处理的Na+/K+明显低于100%浓度。在处理时间上,仍然表现出第4、8 d大幅升高,第12 d升高幅度减缓的总趋势,说明海水处理的第12 d已经接近黄秋葵Na+积累的上限和K+外排的下限。无角香蕉和非洲秋葵两个耐盐性不同的品种在Na+/K+上的差异更加明显,特别是茎和叶的Na+/K+。在不同处理的各个时间点,无角香蕉根的Na+/K+高于非洲秋葵,而茎和叶的Na+/K+则明显低于非洲秋葵。

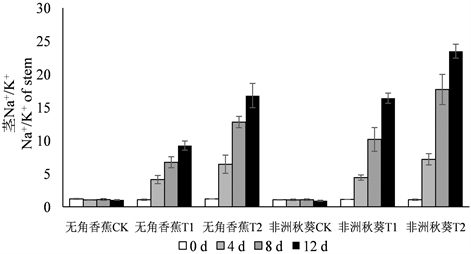
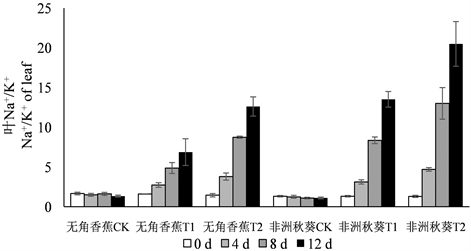
Figure 3. Dynamic change of Na+/K+ in root, shoot, leaf of Wujiaoxiangjiao and Feizhouqiukuiokra under seawater stress
图3. 海水胁迫下黄秋葵无角香蕉和非洲秋葵根、茎、叶中Na+/K+的动态变化
3.5. 海水胁迫对黄秋葵体内Na+、K+在各器官分布的影响
为了研究海水胁迫后Na+和K+在黄秋葵根、茎、叶中的变化规律,将各器官某一处理12 d的Na+含量及K+含量值分别相加,得出整株Na+含量和K+含量总和,再用各器官Na+和K+含量占整株的百分比来分析其在海水胁迫后的分布情况。由图4可知,无角香蕉在受到50%浓度海水胁迫后,根系内Na+含量占整株的比重从对照的40.79%提高到54.28%,而叶片中Na+含量则由32.89%降低到19.21%,100%浓度海水胁迫后Na+含量在各器官中的变化也有类似规律。非洲秋葵在受到50%浓度海水胁迫后,根系内Na+含量占整株的比重从对照的37.96%提高到43.10%,叶片中Na+含量由35.80%降低到26.21%,根内Na+含量升高的幅度和叶片Na+含量降低的幅度明显不及无角香蕉。说明耐盐品种在受到盐胁迫后根系对Na+具有较强的存储功能,叶片则具有较强的Na+外排功能。
海水胁迫后,无角香蕉根系K+含量所占比重由对照的33.91%分别提高到50%海水的36.75%和100%海水的36.79%,茎和叶K+含量占比较对照均有所降低,但降幅很小。非洲秋葵根系K+含量所占比重由对照的31.27%分别提高到50%海水的41.84%和100%海水的42.53%,提高幅度明显高于无角香蕉,而茎和叶K+含量占比的降低幅度要也大于无角香蕉。说明耐盐品种在盐胁迫后能较好的控制茎和叶中的K+含量,并维持它们占整株比率的相对稳定。
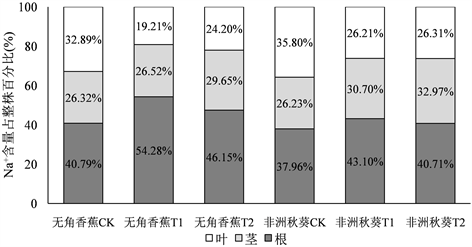
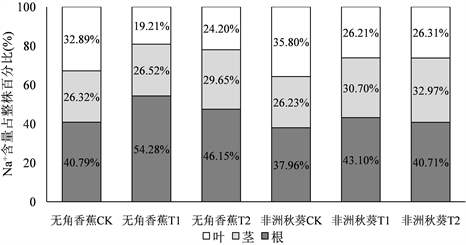
Figure 4. Distribution of Na+, K+ in root, shoot, leaf of Wujiaoxiangjiao and Feizhouqiukuiokra under seawater stress
图4. 海水胁迫下黄秋葵无角香蕉和非洲秋葵Na+、K+在根、茎、叶中的分布情况
4. 讨论与结论
盐分对植物的伤害主要包括离子毒害和渗透胁迫,以及由此而产生的营养和氧化胁迫。以Na+为代表的离子毒害是原初胁迫,不但使植物造成严重的毒害作用,还对植物生长发育必须的其他阳离子产生排斥或竞争作用。因此,研究盐胁迫下植物对Na+和K+的含量变化、区域化分布等特征,对明确植物的耐盐机理至关重要 [22]。本研究对耐盐黄秋葵品种无角香蕉和盐敏感黄秋葵品种非洲秋葵进行50%和100%浓度的海水胁迫处理,分析两个耐盐性不同的品种对海水耐受程度的差异及Na+、K+含量在黄秋葵各器官的变化规律,以明确黄秋葵的耐盐机理及不同耐盐性品种在海水胁迫下的生理相应差异。
植物的生长量,包括幼苗高度、芽长度、根长度、鲜重、干重等,常被作为品种资源耐盐性筛选鉴定的指标 [23] [24] [25] [26]。努尔帕提曼·买买提热依木等 [27] 研究4种十字花科短命植物的耐盐性时发现,在相同盐浓度胁迫下,4种植物的存活天数有明显的不同。Liu等 [28] 通过幼苗存活天数和盐害级别鉴定了708份水稻品种资源的耐盐性,且这两个指标在供试材料中均表现出丰富的变异,说明存活天数也可以作为植物耐盐性鉴定的重要指标。本研究以苗高和存活天数为指标,验证耐盐黄秋葵品种无角香蕉和盐敏感品种非洲秋葵对海水胁迫的耐受性差异,结果表明,50%和100%浓度的海水处理均使无角香蕉和非洲秋葵的苗高生长受到了严重的抑制。50%海水处理下,无角香蕉从第0 d到第12 d的苗高增长量比非洲秋葵多1.72 cm,在100%海水处理下多出1.06 cm。无角香蕉在50%海水处理后的存活天数比非洲秋葵多14天,在100%海水处理后多了3天。因此,从苗高和存活天数上可以反映出无角香蕉比非洲秋葵具有更好的海水耐受能力。另外,两个性状均在处理后两个品种间产生了明显的差异,特别是50%海水处理,这也为海水处理后不同耐性品种Na+和K+含量变化规律的研究奠定了基础。
选用合适浓度的海水处理使不同材料产生明显的性状差异是进行植物耐盐性研究的重要前提。不同植物、同一植物不同品种的耐盐性不同,因此,需要根据自身特点设置不同的处理浓度。李洪燕等 [29] 采用浓度为10%、20%、30%、40%、50%的海水处理苦荬菜幼苗,结果表明,苦荬菜根在小于30%海水处理时与对照差异不显著,叶绿素含量与对照差异也不显著;在10%海水处理下,K+含量与对照差异不显著。王蕾等 [30] 设置浓度为0、10%、20%、30%、40%、50%、60%、70%的海水对番杏幼苗进行胁迫处理,结果显示,番杏能在0%~70%海水胁迫下完成生活史,且10%~40%海水浓度促进了其生物量的增加。唐宁等 [31] 利用浓度为5%、10%、20%、30%、50%、100%的海水处理黄秋葵幼苗,结果表明,在0%~20%低浓度海水处理下,黄秋葵的叶绿素含量和净光合速率均能保存在较高的水平,且丙二醛浓度升高不显著。还有研究显示,黄秋葵能在轻度盐渍化环境中生长 [32]。以上研究均表明,30%以下低浓度的海水处理可能会造成植物受盐害较轻,难以产生明显的性状差异;而如番杏等植物则可以在70%浓度海水下完成生活史,所以可以不设置低浓度海水而有必要设置最高海水浓度100%来探索黄秋葵在其胁迫下的耐受性情况。因此,本研究设置了50%和100%两个海水浓度。50%海水既在每个取样时间点获得了具有较大差异的性状值,在短时间内又没有对黄秋葵植株造成很大的伤害,从而方便了取样和调查。本研究结果也表明,所选黄秋葵品种很难适应100%海水处理,在短时间内会造成植株死亡。
在海水胁迫后,耐盐黄秋葵品种无角香蕉根中的Na+含量高于盐敏感品种非洲秋葵,而茎和叶中的Na+含量要低于非洲秋葵。这与王学征等 [9]、丁同楼等 [33]、王旭明等 [11] 研究不同耐盐性的番茄、小麦和水稻品种Na+含量变化规律一致。海霞等 [34] 研究小麦耐盐性和王永慧等 [18] 研究黄秋葵耐盐性的结果表明,耐盐小麦和黄秋葵品种根中的Na+含量要低于盐敏感品种,并分析根系并无明显贮存盐分的作用,也可能与植物的基因型或处于不同发育时期有关,或者耐盐性较强的品种在盐胁迫下具有更强的Na+外排能力。同样的黄秋葵耐盐实验却与本研究结果并不一致,可能是因为所选黄秋葵不同基因型的原因,或是因为王永慧等 [18] 采用单一盐NaCl而本研究采用海水处理造成的差异。
基金项目
江苏省大学生创新创业训练计划项目(202110304082Y);南通市科技计划项目(JC2020156);中国农业科学院麻类研究所“特色资源功能基因挖掘及功能验证”课题(IBFC-KYSY-080)。
文章引用
刘 岩,程玉静,翟彩娇,王小秋,刘 畅,谢冬微,孙 健,戴志刚. 海水胁迫下黄秋葵Na+、K+含量变化规律研究
Study of the Change Regulation of Na+ and K+ Contents in Okra under Seawater Stress[J]. 植物学研究, 2021, 10(04): 576-586. https://doi.org/10.12677/BR.2021.104072
参考文献
- 1. 云雪雪, 陈雨生. 国际盐碱地开发动态及其对我国的启示[J]. 国土与自然资源研究, 2020(1): 84-87.
- 2. Zhang, J.L. and Shi, H.Z. (2013) Physiological and Molecular Mechanisms of Plant Salt Tolerance. Photosynthesis Research, 115, 1-22. https://doi.org/10.1007/s11120-013-9813-6
- 3. Munns, R. and Tester, M. (2008) Mechanisms of Salinity Tolerance. Annual Review of Plant Biology, 59, 651-681. https://doi.org/10.1146/annurev.arplant.59.032607.092911
- 4. 王俊丽, 高晨旭, 王辉, 朱晔荣, 孙玥, 王勇. 盐胁迫对不同水稻品种幼苗的影响[J]. 植物学研究, 2020, 9(1): 52-58.
- 5. Liang, Y.C. (1999) Effects of Silicon on Enzyme Activity and Sodium, Potassium and Calcium Concentration in Barley Under Salt Stress. Plant and Soil, 209, Article No. 217. https://doi.org/10.1023/A:1004526604913
- 6. Kronzucker, H.J. and Britto, D.T. (2011) Sodium Transport in Plants: A Critical Review. New Phytologist, 189, 54-81. https://doi.org/10.1111/j.1469-8137.2010.03540.x
- 7. 李晓院, 解莉楠. 盐胁迫下植物Na+调节机制的研究进展[J]. 生物技术通报, 2019, 35(7): 148-155.
- 8. Reddy, I.N.B.L., Kim, B.K., Yoon, I.S., Kim, K.-H. and Kwon, T.-R. (2017) Salt Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Science, 24, 123-144. https://doi.org/10.1016/j.rsci.2016.09.004
- 9. 王学征, 李秋红, 吴凤芝. NaCl胁迫下栽培型番茄Na+、K+吸收、分配和转运特性[J]. 中国农业科学, 2010, 43(7): 1423-1432.
- 10. 姜静涵, 关荣霞, 郭勇, 常汝镇, 邱丽娟. 大豆苗期耐盐性的简便鉴定方法[J]. 作物学报, 2013, 39(7): 1248-1256.
- 11. 王旭明, 赵夏夏, 黄露莎, 陈景阳, 莫俊杰, 叶昌辉, 周鸿凯, 谢平. 盐胁迫下4个不同耐盐基因型水稻Na+、K+积累效应[J]. 热带作物学报, 2018, 39(11): 2140-2146.
- 12. 练冬梅, 姚运法, 赖正锋, 洪建基. 黄秋葵果实加工利用研究进展[J]. 中国农学通报, 2016, 32(27): 161-164.
- 13. 张少平, 邱珊莲, 张帅, 郑云云, 吴松海, 何炎森, 郑开斌. 黄秋葵种质资源及相关品种选育研究进展[J]. 农学学报, 2017, 7(6): 49-55.
- 14. Gemede, H.F., Haki, G.D., Beyene, F., Woldegiorgis, A.Z. and Rakshit, S.K. (2015) Proximate, Mineral, and Antinutrient Compositions of Indigenous Okra (Abelmoschus esculentus) Pod Accessions: Implications for Mineral Bioavailability. Food Science & Nutrition, 4, 223-233. https://doi.org/10.1002/fsn3.282
- 15. 邓爱华, 王云, 谢鹏, 彭友林, 易梦媛, 刘康珂. 黄秋葵营养价值研究进展[J]. 农产品加工, 2020(7): 81-84.
- 16. 王永慧, 陈建平, 张培通, 蔡立旺, 施庆华, 王海洋. 黄秋葵耐盐材料的筛选及萌发期耐盐性相关分析[J]. 西南农业学报, 2014, 27(2): 788-792.
- 17. 王永慧, 陈建平, 张培通, 高进, 周春霖, 施庆华, 王海洋. NaCl胁迫对黄秋葵种子萌发和幼苗生理特性的影响[J]. 扬州大学学报(农业与生命科学版), 2015, 36(1): 78-82, 91.
- 18. 王永慧, 陈建平, 张培通, 高进, 周春霖, 施庆华, 周汝琴. 盐胁迫对不同基因型黄秋葵苗期生长及生理生态特征的影响[J]. 华北农学报, 2016, 31(6): 105 -110.
- 19. 刘雅辉, 杨雅华, 王秀萍, 张国新, 鲁雪林. 黄秋葵苗期的盐胁迫反应及耐盐阈值分析[J]. 安徽农业科学, 2017, 45(23): 27-28, 57.
- 20. 吴纪中, 刘妍妍, 王冲, 沈振国, 蔡士宾, 张巧凤, 夏妍, 王桂萍, 陈亚华. 人工海水胁迫下小麦种质资源的耐盐性筛选与鉴定[J]. 植物遗传资源学报, 2014, 15(5): 948-953.
- 21. Lin, H.X., Zhu, M.Z., Yano, M., Gao, J.P., Liang, Z.W., Su, W.A., et al. (2004) QTLs for Na+ and K+ Uptake of the Shoots and Roots Controlling Rice Salt Tolerance. Theoretical and Applied Genetics, 108, 253-260. https://doi.org/10.1007/s00122-003-1421-y
- 22. 刘友良, 汪良驹. 植物对盐胁迫的反应和耐盐性[M]. 北京: 科学出版社, 1999: 767-752.
- 23. 董志刚, 程智慧. 番茄品种资源芽苗期和幼苗期的耐盐性及耐盐指标评价[J]. 生态学报, 2009, 29(3): 1348-1355
- 24. 张士超, 袁芳, 郭建荣, 韩国良, 孙利珍, 王帅, 王宝山. 利用隶属函数值法对甜高粱苗期耐盐性的综合评价[J]. 植物生理学报, 2015(6): 893-902.
- 25. 张玉娟, 游均, 刘爱丽, 黎冬华, 于景印, 王燕燕, 周瑢, 宫慧慧, 张秀荣. 芝麻发芽期耐盐性鉴定方法研究及耐盐候选基因的挖掘[J]. 中国农业科学, 2018, 51(12): 2235-2247.
- 26. 马帅国, 田蓉蓉, 胡慧, 吕建东, 田蕾, 罗成科, 张银霞, 李培富. 粳稻种质资源苗期耐盐性综合评价与筛选[J]. 植物遗传资源学报, 2020, 21(5): 1089-1101.
- 27. 努尔帕提曼•买买提热依木, 齐曼•尤努斯, 谭敦炎. 盐胁迫对4种短命植物种子萌发及植株生长的影响[J]. 西北植物学报, 2011, 31(8): 1618-1627.
- 28. Liu, C., Chen, K., Zhao, X., Wang, X.Q., Shen, C.C., Zhu, Y.J., Dai, M.L., Qiu, X.J., Yang, R.W., Xing, D.Y., Pang, Y.L. and Xu, J.L. (2019) Identification of Genes for Salt Tolerance and Yield-Related Traits in Rice Plants Grown Hydroponically and under Saline Field Conditions by Genome-Wide Association Study. Rice, 12, Article No. 88. https://doi.org/10.1186/s12284-019-0349-z
- 29. 李洪燕, 郑青松, 刘兆普, 李青. 海水胁迫对苦荬菜幼苗生长及生理特性的影响[J]. 植物学报, 2010, 45(1): 73-78.
- 30. 王蕾, 吴朝波, 徐微风, 郭建春, 符少萍, 刘娇, 李瑞梅, 段瑞军. 海水胁迫对番杏生长、渗透调节物质和细胞膜透性的影响[J]. 江苏农业科学, 2016, 44(7): 248-251.
- 31. 唐宁, 张边江, 王立科, 杨平, 陈全战. 海水胁迫对黄秋葵光合及抗氧化生理特性的影响[J]. 分子植物育种, 2019, 17(8): 2639-2645.
- 32. Habib, N., Ashraf, M., Ali, Q. and Perveen, R. (2012) Response of Salt Stressed Okra (Abelmoschus esculentus Moench) Plants to Foliar-Applied Glycine Betaine and Glycine Betaine Containing Sugarbeet Extract, South African. Journal of Botany, 83, 151-158. https://doi.org/10.1016/j.sajb.2012.08.005
- 33. 丁同楼, 贾玉辉, 鲍敬, 王宝山. 不同耐盐性小麦根Na+和K+的吸收特性[J]. 植物生理学报, 2013(1): 34-40.
- 34. 海霞, 刘景辉, 杨彦明, 赵洲, 陈晓晶, 贾硕. 盐胁迫对燕麦幼苗Na+、K+吸收和离子积累的影响[J]. 麦类作物学报, 2019, 39(5): 613-620.