Advances in Geosciences
Vol.08 No.01(2018), Article ID:23712,10
pages
10.12677/AG.2018.81002
The Evidence from the Biomarker in Lake Sediments for the Solar Activity Influences on the Plateau Ecosystem
Yang Pu1, Jinglian Ge1, Tianhao He1, Fengxia Gao1, Hanjie Diao2, Yingying Ding2
1School of Geographical Sciences, Nanjing University of Information Science & Technology, Nanjing Jiangsu
2School of Remote Sensing & Geomatics Engineering, Nanjing University of Information Science & Technology, Nanjing Jiangsu
Received: Jan. 16th, 2018; accepted: Feb. 2nd, 2018; published: Feb. 9th, 2018

ABSTRACT
The relationship between solar activity and the terrestrial ecosystem is an important topic but less mentioned in previous studies. Herein, we applied the biomarker index deduced from the n-alkanes and n-alkanols in lake sediments which are believed to be derived from higher plant exploring the relationship between the solar activity and plateau ecosystem. The results show that the conventional biomarker indexes including carbon preference index (CPI) and average chain length (ACL) sensitively respond to the total solar irradiation (TSI) variation and faithfully recorded four solar minimum periods during the last 900 a. So it is considered that the driving force behind the fluctuations of plateau lacustrine ecosystem is essentially the change in solar radiation intensity.
Keywords:Lake Sediment, Biomarker, n-Alkane, n-Alkanol, Total Solar Irradiation
太阳辐射影响高原生态系统的湖泊沉积生物标志物证据
蒲阳1,葛井莲1,何天豪1,高凤霞1,刁汉婕2,丁莹莹2
1南京信息工程大学,地理科学学院,江苏 南京
2南京信息工程大学,遥感与测绘工程学院,江苏 南京
收稿日期:2018年1月16日;录用日期:2018年2月2日;发布日期:2018年2月9日

摘 要
太阳活动与地球生态系统之间的联系是学术界较少谈及但却十分重要的论题。本文尝试利用青藏高原东部湖泊沉积物中高等植物生物标志物指标探讨高原生态系统与太阳活动之间的关系。结果表明高等植物来源正构烷烃和正构脂肪醇的碳优势指数(CPI)及平均碳链长度(ACL)敏感地响应了总太阳辐照度(TSI)的变化,忠实地记录了青藏高原东部近900 a来四次太阳活动极小期。因此我们认为高原湖泊生态系统波动背后的驱动力实质上是太阳辐射强度的变化。
关键词 :湖泊沉积物,生物标志物,正构烷烃,正构脂肪醇,总太阳辐照度

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
在过去数十年间,已有大量研究证实较长期的气候变化与太阳活动之间存在着密切的联系 [1] 。但已有的研究强调太阳活动对气候因素的驱动,而对于太阳辐射驱动陆生生态系统变化的研究还十分缺乏,主要原因为太阳活动并不是直接作用于生态系统,而是通过气候变化制约生态系统,其作用机制复杂;其次较难寻找到一种客观评估生态系统变化的地球化学指标。
来源于高等植物的类脂物生物标志物为我们认识过去生态系统变化提供了独立的指标体系。它们主要来自各种高等植物的叶片、茎及花朵等器官 [2] 。由于高等植物的寿命相对较长,且种子植物对外界环境有着良好的适应力,因此在其较长的生活史中不断地产生各种生物标志物并保存在沉积物中。而高等植物在多数陆生生态系统中占据着生物量的绝对优势,所产生的生物标志物总量远多于其它类型的生物;另一方面,类脂物生物标志物不易被成岩/成土作用及微生物活动降解,因此无论是相对含量还是分布特征在沉积物及土壤中均保持了良好的稳定性,并且对周围环境条件的改变有着敏感地响应 [3] 。其中长链正构烷烃以及正构脂肪醇是两类最常见的来源于高等植物的类脂物生物标志物 [4] ,在各类沉积物中均能很方便地检测到并开展相关的定量研究。
本文选取青藏高原东部年保玉则地区典型的冰蚀湖希门错为研究区域,对该湖沉积物中来源于高等植物的正构烷烃和正构脂肪醇类生物标志物进行了提取和分析,利用常规的碳优势指数(CPI)及平均碳链长度(ACL)指标表征研究区内高等植物生态系统的波动,并通过生物标志物指标与总太阳辐照度(TSI)的对比,进而探讨太阳活动对高原生态系统的影响。
2. 材料和方法
2.1. 样品的采集及前处理
年保玉则位于青海省果洛州久治县境内(又称果洛山),现代雪线海拔约为5100 m,在最高峰的周围覆盖有现代冰川 [5] (图1)。希门错是该地区的冰川槽谷在冰后期被冰融水充填而成的冰蚀湖。湖面平均海拔约为4020 m,面积3.8 km2,平均水深约为40 m,最深处水深接近65 m。湖水补给主要靠湖区降水
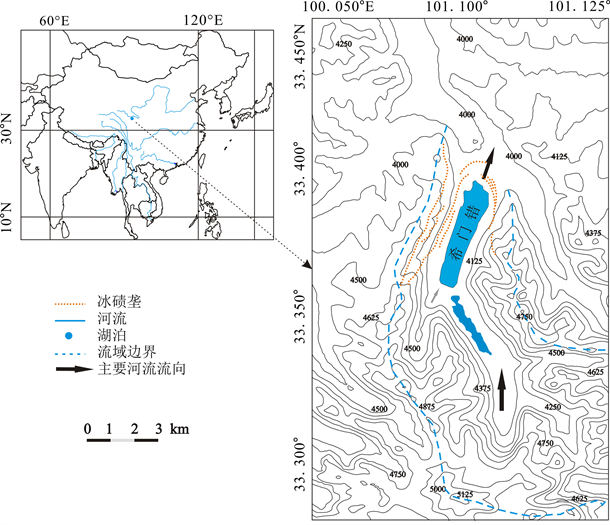
Figure 1. Geographical location and contour map of Lake Ximencuo which is located in Nianbaoye zearea, eastern Qinghai-Tibet Plateau
图1. 青藏高原东部年宝玉则地区希门错湖泊地理位置及等高线地形图
以及冰川融水。
本次研究湖泊沉积物钻孔坐标为N33˚22'40.59/E101˚06'21.78,利用重力采样器取得水深34.4 m处44 cm长的湖泊沉积物岩芯。整个岩芯上下颜色均一,为浅灰色粉砂质粘土,没有受到扰动的痕迹。按2 cm间隔采样共采集样品22个,进行生物标志物分析。然后在相邻水域取平行钻孔,按1 cm间隔采样,开展210Pb及137Cs年代分析。
将每个实验样品称取2 g,研磨至80目以下。采用溶剂-超声波萃取法,将样品用二氯甲烷:甲醇(93:7)混合溶剂浸泡一段时间,再用超声波常温萃取三次,每次20 min,合并萃取液并过滤,将滤液浓缩恒重。湖泊沉积物样品为防止样品中微量组分在分离过程中进一步流失,不再用硅胶-氧化铝色谱柱分离为族组分。自然风干后取出部分样品转移至细胞瓶,经由BSTFA衍生化后,直接进行气相色谱-质谱联用仪(GC/MS)分析。
2.2. 年代测定
图2展示了希门错湖泊沉积物210Pbexc及137Cs比活度随沉积物深度的变化趋势,可以发现210Pbexc及137Cs的比活度在6 cm以后逐渐趋于0 dpm/g,其中210Pbexc比活度随深度大致呈平滑的指数衰减,说明沉积环境相对稳定 [6] 。而Mischke and Zhang, 2010根据希门错沉积物中碱溶性组分和非碱溶性组分的14C年龄建立了希门错湖泊沉积物近16 ka以来的年代框架,14C年代显示至少在近2 ka以来湖泊沉积速率较为稳定 [7] 。故采用恒定沉积速率模型计算得出该岩芯的沉积速率约为0.5 mm/a。而137Cs在2 cm处有一

Figure 2. Excess 210Pb activity and total 137Cs activity in dpm/g vary with depth
图2. 希门错湖泊沉积物过剩210Pb及137Cs活度随深度变化图
个峰值(图2),代表1963年世界各国频繁试爆核弹的时期,据此计算出来的沉积速率与210Pb的结果基本一致。
3. 结果与讨论
3.1. 希门错湖泊沉积物中正构烷烃及正构脂肪醇分布特征及来源
图3展示了希门错表层湖泊沉积物样品(0~2 cm)中正构烷烃及正构脂肪醇生物标志物的分布特征。其中正构烷烃的碳数分布范围为n-C15~n-C33,呈单峰型分布,高碳数正构烷烃(Cn ≥ C21)具有明显的奇碳优势,n-C15~n-C20等低碳数正构烷烃含量较低,无明显的奇偶优势(图3(a))。大多数陆生维管束植物叶片表皮蜡质中含有很高比例的长链正构烷烃,以n-C27、n-C29或n-C31为主峰,具有显著的奇碳优势 [3] 。而水生维管束植物的正构烷烃以n-C21、n-C23或n-C25为主峰 [8] ,因此希门错湖泊沉积物中长链正构烷烃主要来自高等维管类植物,包括陆生以及水生维管类植物。
正构脂肪醇的碳数分布范围是C13~C32,所有样品的高碳数部分(Cn ≥ C22)具有明显的偶碳优势,且高碳数正构脂肪醇的相对含量明显高于低碳数部分的相对含量(图3(b))。已有的研究表明长链偶碳数正构脂肪醇来源于高等植物叶片表层的蜡质,而短链的正构脂肪醇(碳数 ≤ 21)来源于低等菌藻类生物 [9] ,故可认为希门错湖泊沉积物中长链正构脂肪醇主要来自高等维管类植物,其中主要为陆生高等植物,可能包括少量水生维管类植物的贡献。
3.2. 湖泊沉积物生物标志物碳优势指数与太阳活动之间的关系
正构烷烃和正构脂肪醇生物标志物是湖泊沉积物中的化学成分,属于化学种(或者称为分子种) [10] 。已有的研究证实化学种在反映气候、环境条件变化上比形态种(如孢粉)更加灵敏。一些短暂的气候事件并不能导致植物种类发生改变,但是植物的化学成分却发生了细微的变化 [11] 。因此生物标志物在指示气候变化所导致的生态系统波动具有优势。以下我们采用常规的碳优势指数(CPI)以及平均碳链长度(ACL)指标来代表希门错湖区高等植物生态系统的变化。
碳优势指数(CPI)是反映生物标志物所含奇数碳与偶数碳同系物相对丰度的参数。多用于正构烷烃系列的计算,例如来自微生物和藻类等低等生物体的正构烷烃CPI值很低,而高等维管类植物的正构烷烃具有相对较高的CPI值 [3] 。研究表明沉积物中正构烷烃系列CPI值与微生物活动有关,而微生物活动又与气候条件紧密相连,因此CPI值能间接反映气候的变化。例如Zhou et al., 2005在江西定南县泥炭沉积序列中发现正构烷烃与正构脂肪酸的CPI值受到微生物活动的影响,间接反映了研究区的气候变化 [12] 。

Figure 3. Relative abundance versus carbon distribution of n-alkane (a) and n-alkanol (b) homologues in surface sediment of lake Ximencuo
图3. 希门错湖泊表层沉积物正构烷烃(a)及正构脂肪醇(b)生物标志物碳数分布特征
此外在现代表土的地带性研究中也有类似的发现:中国东部地区62个现代表土样品中长链正构烷烃CPI值随着纬度的上升而增加,而与采样地区的植被类型没有必然联系,说明生物标志物CPI值主要受控于气候因素 [13] ;在中国东南沿海至黄土高原北缘的调查中获得了相似的结论,并进一步阐明正构烷烃CPI指标与研究区相对湿度紧密联系 [14] 。而正构脂肪醇系列的CPI指标研究的相对较少,其古环境意义在进一步探索中。为数不多的报道来自于:兰州九州台黄土-古土壤沉积剖面中长链正构脂肪醇CPI值与磁化率呈负相关关系,CPI值能够很好地指示东亚夏季风的活动 [15] ;红原泥炭中正构脂肪酸与正构脂肪醇的CPI表现出很好的一致性,相对较低的CPI值指示了气候历史的暖期,其解译方法与泥炭中正构烷烃CPI指标相同 [16] 。
但需指出的是制约沉积物中生物标志物CPI值变化的因素较多。比如沉积环境的演变 [17] 、植物群落的演替 [15] [18] 、微生物输入或降解程度的变化 [12] [19] 、物质来源的改变 [20] 以及气候变迁 [21] 均有可能改变生物标志物奇数碳与偶数碳同系物的相对丰度,因此直接利用沉积物中生物标志物CPI值来重建古环境的研究较少。本次研究中考虑到正构脂肪醇系列中C22~C28偶碳同系物相对丰度高,故在计算脂肪醇的CPI值时尝试选取了C22~C28偶碳数正构脂肪醇,用CPI22-28-ACH来表示;而正构烷烃系列则选取了C21~C33奇碳数正构烷烃来进行CPI的计算,包含了陆生以及水生高等维管植物的贡献,用CPI21-33-ALK来表示;另一方面,Bard et al., 2000综合北美及欧洲的树轮14C记录和南极冰芯10Be记录,结合数值模型重建了过去1200年以来太阳总辐射能(TSI)的变化(图4) [22] 。本次研究利用该记录中近1000 a以来的TSI记录与生物标志物指标进行对比。由于本次研究中希门错湖泊沉积生物标志物记录的分辨率约为40 a,而用于对比的近千年以来TSI的时间分辨率约为8 a [22] ,前者为后者的5倍。因此对TSI变化曲线进行5点滑动平均,降低两者之间因时间分辨率的不同而产生的误差。
通过对比发现近1000 a以来TSI的5点滑动平均线与希门错湖泊沉积物正构脂肪醇和正构烷烃CPI值有着总体一致的变化趋势,说明CPI指标敏感地响应了太阳活动的变化。特别是近900年以来的四次太阳活动极小期,包括沃尔夫极小期(Wolf Minimum)、斯玻勒极小期(Spörer Minimum)、蒙德极小期(Maunder Minimum)、道尔顿极小期(Dalton Minimum),均在CPI22-28-ACH和CPI21-33-ALK指标上有所体现。而中世纪暖期(MWP)和近代小冰期(LIA)也在CPI指标上有所体现。其中MWP期间CPI值较高,而LIA期间CPI值较低,可能的原因是MWP期间,由于较高的TSI值导致季风活动增强,降水量上升,从而导致更多的陆源碎屑随地表径流及风力搬运到湖泊中沉积下来,而陆源碎屑中大部分的长链正构烷烃及正构脂肪醇来自高等植物,它们均表现出很高的CPI值 [3] ,进而导致沉积物中CPI值升高;而在LIA期间则相反,低TSI值导致环境温度降低,湖泊每年的冰封期增长,导致陆源输入减少,湖泊自生生物

Figure 4. The comparison between the CPI22-28-ACH, CPI21-33-ALK and solar insolation (TSI, w/m2) for the last millennium. W, S, M, D represent Wolf Minimum, Spörer Minimum, Maunder Minimum, Dalton Minimum, respectively
图4. 正构脂肪醇碳优势指数(CPI22-28-ACH)和正构烷烃碳优势指数(CPI21-33-ALK)与近千年以来太阳总辐射能(TSI, w/m2)的对比,阴影部分代表四次明显的太阳活动极小期,W、S、M、D分别代表沃尔夫极小期(Wolf Minimum)、斯玻勒极小期(Spörer Minimum)、蒙德极小期(Maunder Minimum)、道尔顿极小期(Dalton Minimum)
贡献的生物标志物增加,如一些沉水植物和菌藻类生物等,而它们的CPI值一般较陆生高等植物低,因此在LIA期间CPI值呈现低值。
3.3. 湖泊沉积物生物标志物平均碳链长度与太阳活动之间的关系
Poynter引入了平均碳链长度(ACL)的概念,他研究了海洋积物中长链正构烷烃ACL值的变化,并认为该指标可以代表陆地物源区古温度的相对变化 [23] 。但近年来也有学者认为ACL指标反映了源区降水量的变化 [24] 。一般认为降水量稳定而气温上升时,植物的蒸腾作用加强,合成链长较长的类脂物来减少水分损失,温度下降时,植物蒸腾作用减弱,类脂物碳链长度降低;若温度保持稳定而降水量增加时,植物有足够的水分来源,蒸腾作用减弱,ACL值降低,当降水量减小时,蒸腾作用加快,ACL值增加。因此我们认为ACL指标实质是衡量了高等植物类脂物生物标志物的“锁水功能”,即植物对水分的保持能力。
与上一节探讨CPI与TSI之间的关系类似,将近1000 a以来的TSI变化进行5点滑动平均(如图5所示)。正构脂肪醇ACL的计算依然选用C22~C28的正构脂肪醇(表示为ACL22-28-ACH)。而正构烷烃ACL的计算主要集中于C27~C33长链正构烷烃系列(表示为ACL27-33-ALK),中碳数系列的C21至C25系列没有参与计算,主要是考虑到水生植物由于湖水的隔离,对外界环境变化的响应可能相对迟钝。

Figure 5. The comparison between the ACL22-28-ACH, ACL21-33-ALK and solar insolation (TSI, w/m2) for the last millennium. W, S, M, D have the same meaning as in the Figure 4
图5. 正构脂肪醇平均碳链长度(ACL22-28-ACH)和正构烷烃平均碳链长度(ACL21-33-ALK)与近千年以来太阳总辐射能(TSI, w/m2)的对比,阴影部分代表四次明显的太阳活动极小期,W、S、M、D所代表的意义与图4相同
从已有的希门错湖泊沉积物孢粉数据来看,希门错地区的植物群落在近2000 a以来并没有发生植物生活型的演替,但是存在植物种属间相对生物量的波动 [25] 。因此可以认为ACL22-28-ACH和ACL27-33-ALK指标反映的是高等植物生态系统波动的信号。通过对比可以发现ACL指标与TSI的变化趋势十分相似,特别是在近1000 a以来TSI最小的斯玻勒极小期(Spörer Minimum)所对应的时间范围内,ACL22-28-ACH和ACL27-33-ALK指标均呈现了极低值。因此本次研究中的ACL指标主要还是响应了研究区温度的变化:环境温度下降,植物的蒸腾作用下降,导致类脂物的ACL值减小;环境温度升高,植物蒸腾作用上升,类脂物的ACL值变大。这种吻合表明湖泊沉积物中正构烷烃和正构脂肪醇的ACL指标很好地响应了太阳辐射变化所导致的高原生态系统变化,间接指示了研究区的气候变化。
3.4. 湖泊沉积生物标志物记录滞后于太阳活动变化
综合对比生物标志物CPI和ACL指标与1000 a以来TSI之间的关系,不难发现生物标志物指标有滞后于TSI变化的趋势。特别是近900 a以来四次明显的降温过程,其发生和结束的时间均滞后于TSI记录。而生物标志物指标所反映的MWP和LIA事件也滞后于TSI所对应的这两个特殊的气候阶段(图4及图5)。由于湖泊沉积物中年代测定可能存在一定的误差,因此本次研究发现的“滞后效应”的绝对时间是无法给出的,但根据图4和图5所示的四次明显降温过程,滞后的时间范围可初步判定为30~80年。这一结果与前人对生物标志物指标与气候变化指标之间关系的研究较为一致:南美洲卡里亚科盆地沉积物中指示高等植物群落变化的长链脂肪酸ACL24-28指标和C32脂肪酸单体碳同位素值滞后于其他气候变化指标约50 ± 25 a [26] 。
通过已有的研究结论并结合希门错湖区自身的地理环境,我们认为该“滞后效应”可能是由太阳辐射的变化导致地球气候系统内部发生调整 [27] ,进而导致高原生态系统产生相应的波动,随后这些信号保存在高等植被生物标志物中并首先储存在表层土壤中 [28] ,然后随地表径流以及风力搬运等外力作用,最终保存在湖泊沉积物中 [29] ,而这一过程必然要经历一定的时间周期。类似的现象出现在很多与有机指标相关的古环境重建研究中。例如:阿拉伯海东北部2502G孔生物代用指标(Corg%、N%、AL%)表征了过去1200 a以来印度洋季风强度的变化 [30] ,将其与TSI变化进行仔细比对后可以发现文中生物代用指标明显滞后于TSI变化;Liu et al., 2006利用青藏高原东北部青海湖沉积物中长链不饱和烯酮指标(U37 k’和C37:4%)重建了湖区过去3500 a以来的湖水温度以及盐度的变化趋势,通过与同一湖泊沉积物序列中光谱指标进行对比,也可以发现生物标志物指标有明显滞后的现象 [31] ;美国加利福尼亚芭芭拉盆地沉积物序列中长链脂肪酸δD明显滞后于旱涝指数(PDSI)、表层海水温度(SST)以及氧同位素指标(δ18O),而这一滞后现象被解释为生物标志物从高等植物剥离后到被搬运到盆地的沉积中心所需的时间 [32] 。综上所述,我们认为在数百年至千年尺度上,太阳活动影响了地球气候系统的变化,进一步制约了生态系统的波动,而这一波动信号被生物标志物记录下来并最终保存在湖泊沉积物中。这一现象在青藏高原东部希门错湖泊沉积物中得到了很好的体现。因此我们在利用湖泊沉积物中生物标志物指标讨论太阳活动相关问题时,在数百年至千年尺度的研究实例中需考虑这种“滞后效应”。
4. 结论
青藏高原东部希门错湖泊沉积物中来源于高等植物的生物标志物记录了高原生态系统的波动,反映了研究区气候的变化,间接响应了太阳活动。说明太阳辐射强度可能在数百年至千年尺度上是高原湖泊生态系统波动的重要驱动力。
致谢
本项目由2016年度南京信息工程大学大学生实践创新训练计划项目(编号201610300310)及国家自然科学基金(编号41301224)联合资助。
文章引用
蒲 阳,葛井莲,何天豪,高凤霞,刁汉婕,丁莹莹. 太阳辐射影响高原生态系统的湖泊沉积生物标志物证据
The Evidence from the Biomarker in Lake Sediments for the Solar Activity Influences on the Plateau Ecosystem[J]. 地球科学前沿, 2018, 08(01): 9-18. http://dx.doi.org/10.12677/AG.2018.81002
参考文献 (References)
- 1. 王绍武: 全新世气候变化[M]. 气象出版社, 2011.
- 2. Ye, J., Su, J., Chen, K.Y., Liu, H., Yang, X., He, Y.J. and Zhang, W.D. (2014) Comparative Investigation on Chemical Constituents of Flower Bud, Stem and Leaf of Lonicera japonica Thunb. by HPLC-DAD-ESI-MS/MS (n) and GC-MS. Journal of Analytical Chemistry, 69, 777-784. https://doi.org/10.1134/S1061934814080036
- 3. Eglinton, T.I. and Eglinton, G. (2008) Molecular Proxies for Paleoclimatology. Earth and Planetary Science Letters, 275, 1-16. https://doi.org/10.1016/j.epsl.2008.07.012
- 4. Diefendorf, A.F., Freeman, K.H., Wing, S.L. and Graham, H.V. (2011) Production of n-Alkyl Lipids in Living Plants and Implications for the Geologic Past. Geochimica et Cosmochimica Acta, 75, 7472-7485. https://doi.org/10.1016/j.gca.2011.09.028
- 5. Lehmkuhl, F. (1998) Extent and Spatial Distribution of Pleistocene Glaciations in Eastern Tibet. Quaternary International, 45-46, 123-134. https://doi.org/10.1016/S1040-6182(97)00010-4
- 6. 沈吉, 薛滨, 吴敬禄, 吴艳宏, 刘兴起, 羊向东, 刘健, 王苏民. 湖泊沉积与环境演化[M]. 科学出版社, 2010.
- 7. Mischke, S. and Zhang, C.J. (2010) Holocene Cold Events on the Tibetan Plateau. Global and Planetary Change, 72, 155-163. https://doi.org/10.1016/j.gloplacha.2010.02.001
- 8. Ficken, K.J., Li, B., Swain, D.L. and Eglinton, G. (2000) An n-Alkane Proxy for the Sedimentary Input of Submerged/Floating Freshwater Aquatic Macrophytes. Organic Geochemistry, 31, 745-749. https://doi.org/10.1016/S0146-6380(00)00081-4
- 9. Baker, E.A., Ed. (1982) Chemistry and Morphology of Plant Epicuticular Waxes. London.
- 10. 谢树成, Evershed RP. 泥炭分子化石记录气候变迁和生物演替的信息[J]. 科学通报, 2001, 46(10): 863-866.
- 11. 郑艳红, 周卫健, 谢树成, 于学峰. 正构烷烃分子化石与孢粉记录的指示意义对比: 以华南地区为例[J]. 科学通报, 2009, 54(12): 1749-1755.
- 12. Zhou, W., Xie, S., Meyers, P.A. and Zheng, Y. (2005) Reconstruction of Late Glacial and Ho-locene Climate Evolution in Southern China from Geolipids and Pollen in the Dingnan Peat Sequence. Organic Geochemistry, 36, 1272-1284. https://doi.org/10.1016/j.orggeochem.2005.04.005
- 13. Rao, Z., Zhu, Z., Wang, S., Jia, G., Qiang, M. and Wu, Y. (2009) CPI Values of Terrestrial Higher Plant-Derived Long-Chain n-Alkanes: A Potential Paleoclimatic Proxy. Frontiers of Earth Science in China, 3, 266-272. https://doi.org/10.1007/s11707-009-0037-1
- 14. 罗攀, 彭平安, 吕厚远, 郑卓, 王旭. 表土长链正构烷烃CPI值的纬向变化: CPI作为气候干旱程度替代指标的证据[J]. 中国科学: 地球科学, 2012, 42(11): 1729-1741.
- 15. Xie, S., Chen, F., Wang, Z., Wang, H., Gu, Y. and Huang, Y. (2003) Lipid Distributions in Loess-Paleosol Sequences from Northwest China. Organic Geoche-mistry, 34, 1071-1079. https://doi.org/10.1016/S0146-6380(03)00083-4
- 16. Zheng, Y., Zhou, W., Meyers, P.A. and Xie, S. (2007) Lipid Biomarkers in the Zoigê-Hongyuan Peat Deposit: Indicators of Holocene Climate Changes in West China. Organic Geochemistry, 38, 1927-1940. https://doi.org/10.1016/j.orggeochem.2007.06.012
- 17. 蒲阳, 张虎才, 雷国良, 常凤琴, 杨明生, 张文翔, 类延斌, 杨伦庆, 庞有智. 青藏高原东北部柴达木盆地古湖泊沉积物正构烷烃记录的MIS3晚期气候变化[J]. 中国科学: 地球科学, 2010, 40(5): 624-631.
- 18. 张虎才, 杨明生, 张文翔, 雷国良, 常凤琴, 蒲阳, 樊红芳. 洛川黄土剖面S-4古土壤及相邻黄土层分子化石与植被变化[J]. 中国科学D辑: 地球科学, 2007, 37(12): 1634-1642.
- 19. Zheng, Y., Zhou, W. and Meyers, P.A. (2011) Proxy Value of n-alkan-2-ones in the Hongyuan Peat Sequence to Reconstruct Holocene Climate Changes on the Eastern Margin of the Tibetan Plateau. Chemical Geology, 288, 97-104. https://doi.org/10.1016/j.chemgeo.2011.06.011
- 20. Zhang, Z., Zhao, M., Yang, X., Wang, S., Jiang, X., Frank, O. and Geoffrey, E. (2004) A Hydrocarbon Biomarker Record for the Last 40 kyr of Plant Input to Lake Heqing, Southwestern China. Organic Geochemistry, 35, 595-613. https://doi.org/10.1016/j.orggeochem.2003.12.003
- 21. Xie, S., Nott, C.J., Avsejs, L.A., Maddy, D., Chambers, F.M. and Evershed, R.P. (2004) Molecular and Isotopic Stratigraphy in an Ombrotrophic Mire for Paleoclimate Reconstruction. Geochimica et Cosmochimica Acta, 68, 2849-2862. https://doi.org/10.1016/j.gca.2003.08.025
- 22. Bard, E., Raisbeck, G., Yiou, F. and Jouzel, J. (2000) Solar Irradiance during the Last 1200 Years Based on Cosmogenic Nuclides. Tellus B, 52, 985-992. https://doi.org/10.3402/tellusb.v52i3.17080
- 23. Poynter, J.G. and Eglinton, G. (1990) Molecular Composition of Three Sediments from Hole 717C: The Bengal Fan. In: Cochran, J.R., Stow, D.A.V., et al., Eds., Proceedings of the Ocean Drilling Program, Scientific Results, Ocean Drilling Program, College Station, Vol. 116, 155-161.
- 24. Li, L., Li, Q., Tian, J., Wang, H. and Wang, P. (2013) Low Latitude Hydro-Climatic Changes during the Plio-Pleistocene: Evidence from High Resolution Alkane Records in the Southern South China Sea. Quaternary Science Reviews, 78, 209-224. https://doi.org/10.1016/j.quascirev.2013.08.007
- 25. Yang, X. (1996) Pollen Assemblage and Palaeoclimate during Last 2000 Year in Ximen Cuo Region, Qinghai. Acta Micropalaeontologica Sinica, 13, 427-440.
- 26. Hughen, K.A., Eglinton, T.I., Xu, L. and Makou, M. (2004) Abrupt Tropical Vegetation Response to Rapid Climate Changes. Science, 304, 1955-1959. https://doi.org/10.1126/science.1092995
- 27. Crowley, T.J. (2000) Causes of Climate Change over the Past 1000 Years. Science, 289, 270-277. https://doi.org/10.1126/science.289.5477.270
- 28. Galy, V. and Eglinton, T. (2011) Protracted Storage of Biospheric Carbon in the Ganges-Brahmaputra Basin. Nature Geoscience, 4, 843-847. https://doi.org/10.1038/ngeo1293
- 29. Meyers, P.A. and Lal-lier-Verges, E. (1999) Lacustrine Sedimentary Organic Matter Records of Late Quaternary Paleoclimates. Journal of Paleolimnology, 21, 345-372. https://doi.org/10.1023/A:1008073732192
- 30. Agnihotri, R., Dutta, K., Bhushan, R. and Somayajulu, B. (2002) Evidence for Solar Forcing on the Indian Monsoon during the Last Millennium. Earth and Planetary Science Letters, 198, 521-527. https://doi.org/10.1016/S0012-821X(02)00530-7
- 31. Liu, Z., Henderson, A. and Huang, Y. (2006) Alkenone-Based Recon-struction of Late-Holocene Surface Temperature and Salinity Changes in Lake Qinghai, China. GeoRL, 33, L09707.
- 32. Li, C., Sessions, A.L., Valentine, D.L. and Thiagarajan, N. (2011) D/H Variation in Terrestrial Lipids from Santa Barbara Basin over the Past 1400 Years: A Preliminary Assessment of Paleoclimatic Relevance. Organic Geochemistry, 42, 15-24. https://doi.org/10.1016/j.orggeochem.2010.09.011