Bioprocess
Vol.
12
No.
02
(
2022
), Article ID:
51044
,
14
pages
10.12677/BP.2022.122006
不同标签对致病疫霉PiGK5胞外端表达及 稳定性的影响
张乐乐1,刘惠荣1*,韩晓东1,陈阳2
1内蒙古农业大学生命科学学院,内蒙古 呼和浩特
2内蒙古农牧业技术推广中心,内蒙古 呼和浩特
收稿日期:2022年4月2日;录用日期:2022年4月29日;发布日期:2022年5月6日
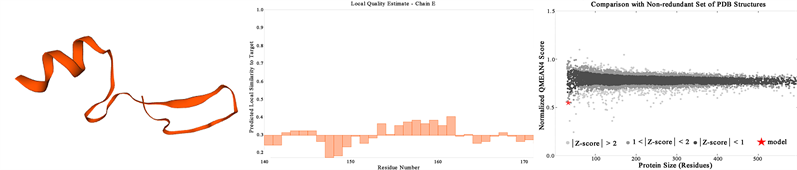
摘要 马铃薯晚疫病病原菌致病疫霉的PiGK5蛋白是一类跨膜蛋白,该蛋白在致病疫霉的致病性、无性生殖及有性生殖方面均有重要作用,但其结构与详细功能目前尚未可知。本研究通过PCR扩增PiGK5胞外端编码区,分别构建融合His标签及MBP和His双标签的原核表达载体,比较了不同标签对PiGK5胞外端表达水平、可溶性及稳定性的影响。结果显示,融合His标签的PiGK5胞外端蛋白主要以包涵体形式存在于沉淀中,仅有少量以可溶性蛋白的形式存在,1 L大肠杆菌表达的蛋白总量为28.11 mg,其中可溶性蛋白总量约为2.73 mg,而纯化后可获得纯蛋白量为0.1175 mg。纯化后的重组蛋白稳定性较好,可在96小时内维持蛋白结构稳定。融合MBP、His双标签的PiGK5胞外端蛋白主要以可溶性蛋白的形式存在,1 L大肠杆菌表达的蛋白总量为49.93 mg,其中可溶性蛋白总量约为44.73 mg,而纯化后可获得纯蛋白量为6.0125 mg。但重组蛋白的稳定性较差,仅在1.5小时就出现了蛋白降解现象。因此,虽然融合His标签的PiGK5胞外端蛋白的可溶性蛋白量较低,但重组蛋白更稳定,融合His标签更适合于PiGK5胞外端的原核表达及后续结构与功能研究。本研究比较了不同标签对PiGK5蛋白胞外端表达的影响,建立了该蛋白的原核表达条件,为其结构与功能的研究奠定了基础。 关键词 致病疫霉,PiGK5蛋白胞外端,标签,表达纯化,稳定性 Effects of Different Tags on the Expression and Stability of Extracellular Terminal of Phytophthora infestans PiGK5 Lele Zhang1, Huirong Liu1*, Xiaodong Han1, Yang Chen2 1College of Life Science, Inner Mongolia Agricultural University, Hohhot Inner Mongolia 2Inner Mongolia Agriculture and Animal Husbandry Technology Popularization Center, Hohhot Inner Mongolia Received: Apr. 2nd, 2022; accepted: Apr. 29th, 2022; published: May 6th, 2022 ABSTRACT PiGK5 protein of potato late blight pathogen Phytophthora infestans is a kind of transmembrane protein, which plays an important role in pathogenicity, asexual reproduction and sexual reproduction of P. infestans. However, the structure and detailed function of this protein have not been known. In this study, the coding region of cDNA of extracellular terminal of PiGK5 was amplified by PCR, and the prokaryotic expression vectors fused with His tag and fused with MBP and His tags were constructed respectively. The effects of different tags on the expression level, solubility and stability of extracellular terminal of PiGK5 were compared. The results showed that the extracellular terminal of PiGK5 fused with His tag mainly existed in the form of inclusion body, and only a small amount of protein was soluble. The total amount of the protein expressed in 1 L culture medium of Escherichia coli was 28.11 mg, in which the total amount of soluble protein was about 2.73 mg, while the amount of the protein obtained after purification was 0.1175 mg. The purified recombinant protein was stable and could maintain structural stability within 96 hours. The extracellular terminal of PiGK5 fused with MBP and His double tags mainly existed in the form of soluble protein. The total amount of the protein expressed in 1 L culture medium of E. coli was 49.93 mg, in which the total amount of soluble protein was about 44.73 mg, while the amount of the protein obtained after purification was 6.0125 mg. However, the stability of the recombinant protein was poor, and the protein degradation appeared only in 1.5 hours. Therefore, although the soluble protein content of PiGK5 extracellular terminal fused with His tag is lower, the recombinant protein is more stable and suitable for prokaryotic expression and follow-up study of structure and function of extracellular terminal of PiGK5. In this study, the effects of different tags on the expression of PiGK5 extracellular terminal were compared, and the prokaryotic expression conditions of the protein were established, which laid a foundation for the study of its structure and function. Keywords:Phytophthora infestans, Extracellular Terminal of PiGK5, Tag, Expression and Purification, Stability Copyright © 2022 by author(s) and Hans Publishers Inc. This work is licensed under the Creative Commons Attribution International License (CC BY 4.0). http://creativecommons.org/licenses/by/4.0/ 1. 引言 疫霉菌(Phytophthora)是属于卵菌纲的一类真菌,能够在大多数经济作物中引起毁灭性疾病,其中以致病疫霉(Phytophthora infesstans)最为严重。致病疫霉(P. infesstans)是马铃薯晚疫病的病原菌,也是疫霉属的模式种。致病疫霉主要会导致马铃薯和番茄晚疫病,曾经在1845年,引发爱尔兰马铃薯饥荒 [1]。今天,它仍然是马铃薯生产的最严重威胁,导致世界范围内的巨大经济损失 [2]。全球每年因致病疫霉造成的经济损失高达170亿美元,其中,我国每年的损失就达40亿美元。 致病疫霉存在一类特殊的G蛋白偶联受体(G protein-coupled receptor, GPCR),这个GPCR的N端是典型的七次跨膜的螺旋结构,C端有一个磷脂酸肌醇激酶(Phosphatidyl inositol phosphokinase, PIPK)结构域(GPCR-PIPK) [3]。对致病疫霉的基因组测序工作已完成,生物信息学分析结果表明,致病疫霉含有12个GPCR-PIPKs (PiGKs)成员 [4] [5] [6] [7]。 研究显示,致病疫霉的PiGK蛋白家族的PiGK5蛋白在致病疫霉的有性生殖、无性生殖及致病性中均起重要作用 [5]。致病疫霉的PiGK5蛋白定位于原生质体膜上及原生质体内部的一些颗粒状结构中。PiGK5在游动孢子中相对表达量最高,在休止孢、营养菌丝、孢子囊等中表达量均相对较少。致病疫霉游动孢子的数量随PiGK5基因表达水平的降低而显著减少,菌丝生长速率也随PiGK5基因表达水平的降低而明显减小。PiGK5沉默转化子的卵孢子数量大量减少,比对照菌株的卵孢子数量减少高达20倍。此外,随PiGK5基因沉默水平增加,致病疫霉游动孢子的侵染能力明显下降,侵染病斑出现时间变晚,且病斑面积的扩大也变慢。PiGK5蛋白包括920个氨基酸,其胞外端由173个氨基酸组成。在该蛋白参与的生物过程中,其胞外端很可能起着与胞外信号分子特异性识别和结合的作用。因此,本研究克隆了PiGK5胞外端编码区并构建了原核表达载体,比较了不同标签对蛋白表达水平、可溶性及稳定性的影响,建立了PiGK5胞外端蛋白表达纯化条件,为其结构及功能的进一步研究奠定了基础,对未来在该蛋白基础上开发有效防治马铃薯晚疫病的靶向药物有重要意义。 2. 实验材料 2.1. 主要试剂及仪器设备 高保真DNA聚合酶Prime STAR GXL,T4 DNA聚合酶,Xba I、Xho I、Ssp I限制性核酸内切酶及琼脂糖凝胶DNA回收试剂盒均购自大连宝生物工程有限公司。Protein Ruler II未染色蛋白Marker购自北京全式金生物技术有限公司,低分子量蛋白质Merker、BCA蛋白浓度测定试剂盒均购自北京索莱宝科技有限公司。质粒小量制备试剂盒购自上海捷瑞生物工程有限公司,超声波信号发生器购自上海比朗仪器制造有限公司,AKTATM prime Plus蛋白纯化系统购自瑞典AKTA公司,10 kD截留规格的超滤离心管购自美国Millipore公司,Trans-Blot转印槽购自美国BIO-RAD公司,Odyssey红外激光扫描成像系统购自美国LI-COR公司。 2.2. 实验菌株及载体 大肠杆菌DH5α、BL21(DE3)菌株购自北京全式金生物技术有限公司。pET30a(+)载体来自本实验室,pTEV载体是在pET30a(+)的His标签前插入麦芽糖结合蛋白(Maltose Binding Protein, MBP)标签的双标签改造载体(图1)。 3. 实验内容 3.1. PiGK5蛋白胞外端的生物信息学分析 通过在线工具Expasy对PiGK5蛋白胞外端进行理化性质分析,在线工具Sopma进行二级结构预测,在线工具SWISS-MODEL进行三维结构预测,得到PiGK5蛋白胞外端的分析数据。 3.2. PiGK5胞外端cDNA编码区的PCR扩增 以实验室保存的PiGK5cDNA编码区克隆载体为模板,PCR扩增PiGK5胞外端cDNA编码区。引物R:TTATCCACTTCCAATGCTActgagacttgcggttctcgaac (大写序列为pET30a(+)载体序列,小写序列为PiGK5胞外端cDNA编码区序列);引物F:TACTTCCAATCCAATGCCatgtcaaggtcatacagtac (同引物R)。PCR反应条件:98℃ 2 min;98℃ 10 s,63.1℃ 15 s,72℃ 3 min,30个循环;72℃ 10 min。PCR扩增产物经电泳后,切取大小正确的条带进行DNA回收。 Figure 1. Expression vector for experiment 图1. 实验用表达载体 3.3. PiGK5胞外端原核表达载体的构建 将pET30a(+)载体/pTEV载体用Ssp I限制性核酸内切酶37℃酶切2 h,琼脂糖凝胶电泳后,酶切的载体片段进行DNA回收。 回收的线性pET30a(+)载体/pTEV载体与T4 DNA聚合酶、dGTP混匀后,于22℃孵育30 min,75℃孵育20 min,利用T4 DNA聚合酶的3'→5'外切酶特性,酶切为具有粘性末端的线性pET30a(+)载体/pTEV载体(图2)。 回收的PiGK5胞外端cDNA编码区片段与T4 DNA聚合酶、dCTP混匀后,于22℃孵育30 min,75℃孵育20 min,同理,酶切为具有粘性末端的PiGK5-N端片段(图2)。 粘性末端互补的PiGK5胞外端cDNA编码区片段与线性pET30a(+)载体/pTEV载体混匀后于22℃孵育5 min,转化入大肠杆菌DH5α感受态细胞,被大肠杆菌中的DNA连接酶连接为环状重组质粒。将含有重组质粒的大肠杆菌DH5α涂布于Kan浓度为50 μg/mL的LB平板,筛选阳性克隆菌株,进行PCR鉴定。菌落PCR鉴定为阳性的菌株扩大培养并提取质粒,进行Xba I、Xho I限制性核酸内切酶的双酶切鉴定。酶切鉴定为阳性的菌液送生工测序。测序正确的表达载体分别命名为pET30a(+)-PiGK5-N和pTEV-PiGK5-N。 3.4. 重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His的表达与纯化 将鉴定正确的载体pET30a(+)-PiGK5-N和pTEV-PiGK5-N分别转化入大肠杆菌BL21(DE3)感受态细胞,涂布于Kan浓度为50 μg/mL的LB平板,挑取单克隆菌株接种于10 mL Kan浓度为50 μg/mL的LB液体培养基活化,以37℃ 180 r/min培养至菌液OD600为0.6。按1:100的比例转接至1 L Kan浓度为50 μg/mL的LB液体培养基,以37℃180 r/min培养至菌液OD600为0.6。然后,将培养温度转为16℃,添加一定浓度IPTG,180 r/min过夜16 h培养诱导目的蛋白表达。室温4500 r/min离心15 min收集菌体沉淀。以同等条件下培养的BL21(DE3)空白菌株及含空载体pET30a(+)/pTEV的BL21(DE3)菌株为对照。 Figure 2. Construction of prokaryotic expression vector pET30a(+)-PiGK5-N 图2. 原核表达载体pET30a(+)-PiGK5-N的构建 用30 mL柱平衡缓冲液(50 mM Tris-HCl, 500 mM NaCl, pH 8.0)重悬菌体沉淀。超声破碎(30%功率,时间20 min,工作2.0 s,间隔4.0 s)菌体,4℃ 18,000 r/min离心40 min得到菌体沉淀与上清液。 使用His Trap TM IMAC (镍柱),分离上清液中的重组蛋白。取上清液缓慢(0.6 mL/min)流经预平衡后的镍柱,用0 mM、10 mM、20 mM、50 mM、100 mM、200 mM、500 mM咪唑的洗脱缓冲液(25 mM Tris-HCl, 500 mM NaCl, pH 8.0)依次洗脱镍柱,收集各洗脱液,并与沉淀、上清液分别取样,进行SDS-PAGE凝胶电泳。 观察SDS-PAGE电泳图,取含有可溶性目的蛋白的洗脱液,用截留规格为10 kD的超滤离心管进行浓缩,浓缩体积至2 mL。0.22 μm滤器过滤后使用AKTATM prime Plus蛋白纯化系统分离纯化蛋白,结合紫外吸收峰收集目的蛋白,SDS-PAGE电泳验证蛋白分子量正确,用超滤离心管对含有目的蛋白的洗脱液进行脱盐处理,得到PiGK5胞外端-His和PiGK5胞外端-MBP-His两种蛋白。 使用软件Image J对两种蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His的SDS-PAGE电泳图进行蛋白条带的灰度分析,得到两种蛋白表达与纯化过程中最合适的IPTG添加浓度。 3.5. 重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His的蛋白浓度与稳定性检测 使用BCA蛋白浓度测定试剂盒测定蛋白浓度。将重组蛋白置于4℃,避光的条件下保存。重组蛋白PiGK5胞外端-His间隔时间24 h,连续5次取样;重组蛋白PiGK5胞外端-MBP-His间隔1.5 h,连续3次取样。SDS-PAGE凝胶电泳,观察两种重组蛋白的稳定性。 3.6. 可溶性重组蛋白PiGK5胞外端-His的Western Blot检测 配制好的12%分离胶4℃过夜,然后配置5%的浓缩胶。每孔蛋白样品的上样体积为20 μL。浓缩胶的电泳电压为90 V,时间30 min;分离胶的电泳电压为120 V,时间90 min。蛋白通过半干转转膜方法转移到PVDF膜上,然后用5%脱脂奶粉封闭3~4 h,100 r/min孵育兔抗His标签的一抗(1:1000,5% BSA稀释) 4℃过夜,1 × TBST清洗一抗,100 r/min避光孵育LICOR IRDye 800 CW山羊抗兔IgG(H + L)二抗(1:10,000,5% BSA稀释) 90 min结束,1 × TBST避光清洗二抗,最后用Odyssey红外激光扫描成像系统照膜,检测目的蛋白PiGK5胞外端-His蛋白。 4. 实验结果 4.1. PiGK5蛋白胞外端的生物信息学分析 使用Expasy预测得到的PiGK5蛋白胞外端的理化性质结果如下,PiGK5蛋白胞外端的氨基酸数目为173个,分子量约为19.3 kD,分子式为C809H1290N262O271S8,理论等电点为10.43,在大肠杆菌中的预计半衰期大于600分钟,不稳定指数为71.98,为不稳定蛋白;亲水平均值为−0.984,为亲水蛋白。 使用Spoma预测得到的PiGK5蛋白胞外端的二级结构分析如图所示(图3),23个α螺旋(蓝)占比13.29%,10个β折叠(绿)占比5.78%,112个无规则卷曲(紫)占比64.74%,28个延伸链(红)占比16.18%。 SWISS-MODEL预测的PiGK5蛋白胞外端的三维结构如图所示(图4),与PiGK5蛋白胞外端相似的模型SMTL ID为4qiw.2,序列一致性为21.88%。 Figure 3. Analysis of the secondary structure of extracellular terminal of PiGK5 protein 图3. PiGK5蛋白胞外端的二级结构分析 4.2. PiGK5胞外端cDNA编码区的PCR扩增 PCR扩增PiGK5胞外端cDNA编码区片段如图5(a),PiGK5胞外端cDNA编码区为519 bp,在琼脂糖凝胶电泳图中的DNA条带大小约为500 bp,二者大小相符,初步说明PiGK5胞外端cDNA编码区的PCR扩增成功。 Figure 4. Analysis of the tertiary structure of extracellular terminal of PiGK5 protein 图4. PiGK5蛋白胞外端的三维结构分析 4.3. 表达载体pET30a(+)-PiGK5胞外端和pTEV-PiGK5胞外端的构建 将具有粘性末端的线性pET30a(+)载体/pTEV载体与PiGK5胞外端cDNA编码区片段孵育后转化入大肠杆菌DH5α感受态细胞。菌落PCR鉴定结果显示(图5(b)、图5(c)),阳性克隆菌株经过扩增得到大小为500 bp的片段,与预计扩增片段519 bp大小相符。对菌落PCR鉴定为阳性的菌株进行Xho I、Xba I限制性核酸内切酶的酶切鉴定,将酶切鉴定为阳性的菌株送公司测序,结果显示,DNA序列无突变,且读码框正确,说明载体构建成功。 Figure 5. PCR amplification of cDNA coding region of extracellular terminal of PiGK5 and colony PCR identification of the recombinants 图5. PiGK5胞外端cDNA编码区的PCR扩增及重组子的菌落PCR鉴定 4.4. 重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His的表达与纯化 将测序正确的表达载体pET30a(+)-PiGK5-N和pTEV-PiGK5-N分别转化入大肠杆菌BL21(DE3)感受态细胞。低温16℃添加终浓度为100 μM IPTG诱导目的蛋白表达,收集菌体沉淀,将菌体超声破碎后取上清液和沉淀进行SDS-PAGE电泳。 电泳结果显示(图6),与其他样品相比在pET30a(+)-PiGK5-N载体转化菌株的菌体沉淀中看到一条染色非常深的分子量约为25 kD的蛋白带,说明该蛋白条带可能为大肠杆菌表达的PiGK5胞外端,且该蛋白主要在大肠杆菌的包涵体中表达。PiGK5胞外端-His蛋白的预测分子量为20.1 kD,在电泳结果中显示的蛋白分子量比预测的蛋白分子量略大,分析其原因可能是,PiGK5胞外端蛋白是碱性蛋白,电泳时碱性蛋白结合的SDS偏少,携带净负电荷低于预期,泳动速率偏慢,迁移距离较小;此外,氨基酸侧链向外延伸,导致蛋白的流体动力学体积大于预期,空间位阻效应较大,迁移距离较小 [8] [9] [10]。 在pTEV-PiGK5-N载体转化菌株的菌体破碎上清液及沉淀中均看到一条约66 kD的蛋白带,而在空白与空载体对照菌株的沉淀和上清中均未看到相应蛋白条带,说明该蛋白条带为大肠杆菌表达的连有MBP标签的PiGK5胞外端蛋白,且该蛋白在大肠杆菌中主要以可溶性的状态表达。PiGK5胞外端-MBP-His蛋白的预测分子量为62.1 kD,电泳结果中的表观分子量稍大于实际分子量,可能原因同PiGK5胞外端-His蛋白一致。 Figure 6. SDS-PAGE results of extracellular terminal-His and extracellular terminal-MBP-His of PiGK5 induced by IPTG 图6. IPTG诱导的PiGK5胞外端-His和PiGK5胞外端-MBP-His的SDS-PAGE结果 Figure 7. SDS-PAGE result of extracellular terminal-His of PiGK5 purified by nickel column affinity chromatography 图7. 重组蛋白PiGK5胞外端-His的镍柱亲和层析SDS-PAGE结果 鉴于重组蛋白PiGK5胞外端-His的表达量较少,接下对诱导表达该蛋白的最佳IPTG浓度进行了优化。分别加入100 μM、150 μM、200 μM和300 μM的IPTG于16℃诱导目的蛋白表达后收集菌体沉淀,将菌体超声破碎后取上清液过His TrapTM IMAC柱(镍柱),用含不同浓度咪唑的洗脱缓冲液进行洗脱。SDS-PAGE电泳结果再次显示(图7),重组蛋白PiGK5胞外端-His主要以包涵体形式存在于沉淀中,只有少量存在于上清中。在咪唑浓度为50 mM和100 mM的洗脱缓冲液中可以看到少量可溶性重组蛋白PiGK5胞外端-His。IPTG浓度为200 μM时得到的可溶性重组蛋白PiGK5胞外端-His最多,因此,确定重组蛋白PiGK5胞外端-His诱导表达的最佳IPTG浓度为200 μM。 重组蛋白PiGK5胞外端-MBP-His表达时,添加常规的100 μM IPTG时即可得到较高产量的可溶性重组蛋白PiGK5胞外端-MBP-His,因此未尝试用其他浓度的IPTG诱导。SDS-PAGE电泳结果表示(图8),重组蛋白PiGK5胞外端-MBP-His主要以可溶性蛋白的形式存在于上清液中,在咪唑浓度为50 mM和100 mM的洗脱缓冲液中含有大量的可溶性重组蛋白,在咪唑浓度为200 mM和500 mM的洗脱缓冲液中也含有少量的可溶性重组蛋白。 Figure 8. SDS-PAGE result of extracellular terminal-MBP-His of PiGK5 purified by nickel column affinity chromatography 图8. 重组蛋白PiGK5胞外端-MBP-His的镍柱亲和层析SDS-PAGE结果 以蛋白电泳采用的ProteinRuler II未染色蛋白Marker的蛋白条带为标准,使用软件Image J对图7和图8的SDS-PAGE电泳图进行蛋白条带的灰度分析,得到目的蛋白电泳条带的蛋白量。灰度分析得到的不同浓度IPTG诱导下重组蛋白PiGK5胞外端-His的表达量及100 μM IPTG诱导下重组蛋白PiGK5胞外端-MBP-His的表达量,其结果与SDS-PAGE电泳结果相似。IPTG添加终浓度为200 μM时,1 L菌体培养得到的重组蛋白PiGK5胞外端-His表达量最高,总蛋白量约为28.11 mg;其中,可溶性蛋白占比9.71%,为2.73 mg。IPTG添加终浓度为100 μM时,1 L菌体培养得到的重组蛋白PiGK5胞外端-MBP-His的总蛋白量约为49.93 mg;其中,可溶性蛋白占比89.59%,为44.73 mg (表1)。 Table 1. Protein expression level of extracellular terminal-His and extracellular terminal-MBP-His of PiGK5 induced by different concentrations of IPTG 表1. 重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His在不同浓度IPTG诱导下的蛋白表达量 表中的“—”表示未进行测量与分析。 将可溶性重组蛋白PiGK5胞外端-His的50 mM咪唑的洗脱缓冲液和100 mM咪唑的洗脱缓冲液浓缩、过滤后经AKTATM prime Plus蛋白纯化系统纯化,收集有明显紫外吸收峰的洗脱液。电泳结果如图所示(图9(a)),第二个吸收峰的蛋白条带位于20 kD与30 kD之间,与重组蛋白PiGK5胞外端-His的预测分子量20.1 kD相符,所以第二个吸收峰的蛋白应该是重组蛋白PiGK5胞外端-His。 Figure 9. SDS-PAGE result of recombinant protein purified by AKTATM prime Plus protein purification system 图9. AKTATM prime Plus蛋白纯化系统纯化后的重组蛋白SDS-PAGE结果 将可溶性重组蛋白PiGK5胞外端-MBP-His的50 mM咪唑的洗脱缓冲液和100 mM咪唑的洗脱缓冲液进行相同处理。经过AKTATM prime Plus蛋白纯化系统纯化后仅出现一组有明显紫外吸收的洗脱液。电泳结果如图所示(图9(b)),在60 kD处有蛋白条带,与重组蛋白PiGK5胞外端-MBP-His的预测分子量62.1 kD相符,应该为重组蛋白PiGK5胞外端-MBP-His条带。但在小于60 kD的下方有几条大小不等的蛋白带,随着洗脱时间的延长,约60 kD的蛋白带呈现越来越弱的趋势,而下方分子量较小的蛋白带却随着洗脱时间有增强的趋势,说明重组蛋白在较长时间的处理纯化过程中可能发生了降解。 4.5. 获得的可溶性重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His纯蛋白量及其稳定性 将含有可溶性重组蛋白PiGK5胞外端-His的1~3号管洗脱液浓缩至500 μL,BCA蛋白浓度测定试剂盒测定蛋白浓度,每1 L菌体纯化获得的可溶性蛋白PiGK5胞外端-His蛋白量为0.1175 mg。可溶性重组蛋白PiGK5胞外端-MBP-His以相同方法测定,每1 L菌体纯化获得的可溶性蛋白PiGK5胞外端-MBP-His蛋白量为6.0125 mg。 500 μL的可溶性重组蛋白PiGK5胞外端-His,于4℃避光保存,每24小时取样一次,连续5次取样,并进行SDS-PAGE电泳验证,结果如图所示(图10(a)),重组蛋白PiGK5胞外端-His在96小时内都保持相对稳定,未出现明显的降解现象。 Figure 10. SDS-PAGE result of stability test of soluble recombinant protein 图10. 可溶性重组蛋白稳定性检测的SDS-PAGE结果 将100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His于4℃避光保存,每1.5小时取样一次,进行SDS-PAGE电泳检测,结果如图所示(图10(b)),重组蛋白PiGK5胞外端-MBP-His结构极不稳定,1.5小时后就出现了降解情况;之后,随着时间的延长,重组蛋白PiGK5胞外端-MBP-His的降解情况进一步加剧。 4.6. 可溶性重组蛋白PiGK5胞外端-His的Western Blot检测 通过Weatern Blot检测表达的PiGK5胞外端-His,结果(图11)表明,PiGK5胞外端-His蛋白在大肠杆菌BL21(DE3)菌株中得到了表达,大小约为25 kD,比蛋白的实际分子量稍大,造成此现象的可能原因同上文。 5. 讨论 马铃薯晚疫病是一种毁灭性的马铃薯病害,但由于马铃薯晚疫病的致病机制、生殖调控机制和相关调控物质等的现有数据有限,还没有有效的方法来预防和控制该病害。目前防治马铃薯晚疫病的常用方式是化学药剂防治。但在马铃薯生产过程中频繁使用杀菌剂 [11],不但会破坏环境,而且会导致致病疫霉耐药性的增加。 Figure 11. Western Blot detection of extracellular terminal-His of PiGK5 purified by nickel column affinity chromatography 图11. 镍柱纯化的PiGK5胞外端-His蛋白的Western Blot检测 致病疫霉是马铃薯晚疫病的病原菌,其无性生殖和有性生殖对病害的发生和传播起着非常重要的作用。致病疫霉无性生殖产生孢子囊,有性生殖产生卵孢子。致病疫霉的孢子囊很轻,能够在风速为1~2 m/s的情况下,3小时内向空中传播20公里。孢子囊的这一特性,使得致病疫霉能够快速扩散,导致马铃薯晚疫病的大范围爆发 [12]。卵孢子作为致病疫霉的一种生殖结构,优势在于卵孢子的细胞壁很厚,可以在恶劣的环境条件下休眠数年 [13] [14] [15]。卵孢子对马铃薯晚疫病的大流行有重要意义,原因为卵孢子是越冬后侵染马铃薯的主要病原菌结构 [16] ;有性生殖可以实现基因重组,进而可能会产生新的基因型菌株,增强菌株的致病性和抗药性 [17]。已有的研究结果显示,PiGK5蛋白与致病疫霉的致病性、无性生殖及有性生殖均密切相关。因此,PiGK5蛋白的结构与功能研究对阐明致病疫霉的致病及生殖机制有重要意义,对未来在该蛋白基础上开发有效防治马铃薯晚疫病靶向生物农药具有重要指导作用。 然而,PiGK5蛋白属于跨膜蛋白,具有疏水性的特点,在大肠杆菌中重组表达时易出现低表达、错误折叠等问题,正确表达时往往会以包涵体的形式表达 [18]。基于PiGK5蛋白的这一特点,分段表达PiGK5蛋白的胞外端和胞内端对研究该蛋白的结构与功能具有重要意义。PiGK5蛋白胞外端由173个氨基酸组成。在该蛋白参与的生物过程中,其胞外端很可能起着与胞外信号分子特异性识别和结合的作用。因此,本研究比较了不同标签对PiGK5蛋白胞外端表达水平、可溶性及稳定性的影响。 在表达重组蛋白时,融合标签的选择极为重要。实验中常用的标签包括谷胱甘肽S-转移酶(GST)、硫氧还蛋白(TRX)、His标签和麦芽糖结合蛋白(MBP)等,其主要作用为增强重组蛋白在大肠杆菌中的可溶性表达及便于重组蛋白的纯化和鉴定。 His标签是由六个组氨酸残基组成的融合标签,可连接在目的蛋白的N端或C端。His标签较其他融合标签分子量小,只有0.84 kD左右,与目的蛋白融合后基本不会改变蛋白的生物结构及溶解性 [19]。并且His残基侧链与固态镍有较强烈的吸引力,可以用固定化金属螯合层析(IMAC)对重组蛋白实现分离纯化。除此之外,His标签可以应用于多种表达系统,且纯化条件较为温和。使用His标签,构建的重组蛋白PiGK5胞外端-His,在软件Image J预测时1 L菌体培养可得到可溶性蛋白量为2.73 mg。但经过蛋白分离与纯化,1 L菌体纯化获得可溶性蛋白PiGK5胞外端-His蛋白量仅为0.1175 mg。 His标签的另一个的优势是因为其分子量很小可以与其他亲和标签构建双亲和标签。MBP标签能够显著增强目的蛋白的溶解度 [20]。MBP由malE基因编码,分子量为42 kD [21]。对MBP、GST和TRX这三种可溶性融合标签进行了比较,发现MBP是一种比其他两种融合标签更有效的增溶标签。对6种不同多肽实验得到的结果,MBP融合蛋白的可溶性蛋白表达量约为其他两种标签融合蛋白可溶性蛋白表达量的三倍左右 [20]。并且MBP在蛋白质的合成期间及合成之后能够起到稳定及保护融合蛋白免受蛋白酶降解的作用 [22],因此可以在大肠杆菌中获得大量的MBP融合蛋白 [23]。在本实验中,也借鉴了His标签和MBP标签双融合标签的优点,既具有MBP标签的增溶作用,又具有His标签的易于分离纯化的作用。使用MBP和His双标签表达重组蛋白PiGK5胞外端-MBP-His,1 L菌体纯化获得可溶性蛋白的PiGK5胞外端-MBP-His蛋白量为6.0125 mg,是重组蛋白PiGK5胞外端-His可溶性表达量的51倍左右,与文献报道MBP标签可促进蛋白的可溶性表达一致。但是,重组蛋白PiGK5胞外端-MBP-His的稳定性明显下降,在1.5 h时就出现了蛋白降解现象。分析蛋白降解的可能原因,大肠杆菌或环境中的蛋白酶导致重组蛋白的降解;蛋白纯化时间过长,且室温环境不利于蛋白结构的稳定;纯的蛋白溶液容易发生降解。对蛋白降解的可能原因做出的相应改进方法,在缓冲液或洗脱液中添加适量的蛋白酶抑制剂,蛋白提取全程置于冰上进行,蛋白质干粉或蛋白溶液加甘油于−20℃保存。 6. 结论 本实验检测了不同标签对PiGK5蛋白胞外端表达的影响,结果表明,1 L大肠杆菌表达的可溶性PiGK5胞外端-MBP-His蛋白量为44.73 mg,获得纯蛋白量为6.0125 mg;可溶性PiGK5胞外端-His蛋白量为2.73 mg,获得纯蛋白量为0.1175 mg。表达的可溶性蛋白量前者是后者的16.38倍,获得纯蛋白量前者是后者的51.17倍。重组蛋白PiGK5胞外端-His可在长达96 h内维持蛋白结构的稳定,但重组蛋白PiGK5胞外端-MBP-His仅在1.5 h时就出现了蛋白降解现象,蛋白结构极不稳定 综合比较重组蛋白PiGK5胞外端-His和PiGK5胞外端-MBP-His的特性,虽然融合His标签的PiGK5胞外端蛋白的可溶性蛋白量较低,但重组蛋白更稳定,融合His标签更适合于PiGK5胞外端的原核表达及后续结构与功能研究。 基金项目 国家自然科学基金“性激素α2调控的疫霉菌有性生殖细胞信号转导通路的初步研究”(编号31760252)。 文章引用 张乐乐,刘惠荣,韩晓东,陈 阳. 不同标签对致病疫霉PiGK5胞外端表达及稳定性的影响 参考文献 NOTES *通讯作者。





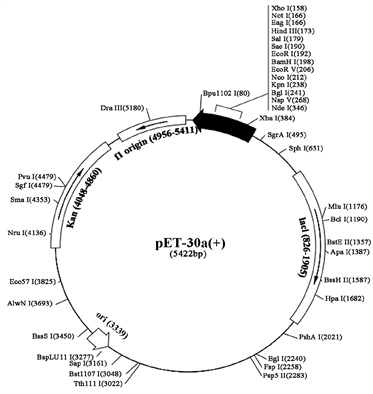 (a) pET30a(+)载体
(a) pET30a(+)载体 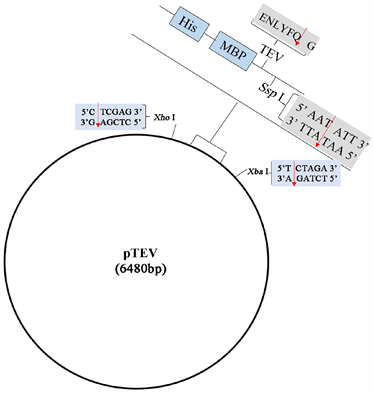 (b) pTEV载体
(b) pTEV载体 (a) pET30a(+)-PiGK5-N载体的构建过程
(a) pET30a(+)-PiGK5-N载体的构建过程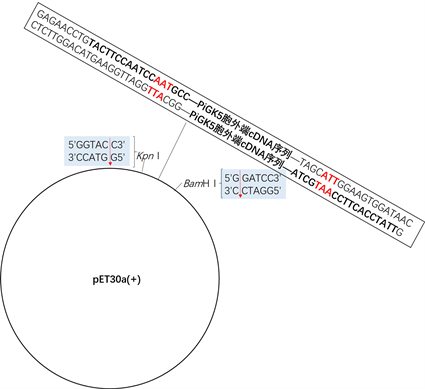 (b) 载体pET30a(+)-PiGK5-N的结构
(b) 载体pET30a(+)-PiGK5-N的结构
 (a) PiGK5胞外端cDNA编码区的PCR扩增。M:DL 1000 DNA Marker;1~3:PiGK5胞外端cDNA编码区的PCR扩增结果。(b) 含pET30a(+)-PiGK5-N载体的菌落PCR扩增。M:DL 500 DNA Marker;1~3:含pET30a(+)-PiGK5-N载体的菌落PCR扩增结果。(c) 含pTEV-PiGK5-N载体的菌落PCR扩增。M:DL 500 DNA Marker;1~3:含pTEV-PiGK5-N载体的菌落PCR扩增结果。
(a) PiGK5胞外端cDNA编码区的PCR扩增。M:DL 1000 DNA Marker;1~3:PiGK5胞外端cDNA编码区的PCR扩增结果。(b) 含pET30a(+)-PiGK5-N载体的菌落PCR扩增。M:DL 500 DNA Marker;1~3:含pET30a(+)-PiGK5-N载体的菌落PCR扩增结果。(c) 含pTEV-PiGK5-N载体的菌落PCR扩增。M:DL 500 DNA Marker;1~3:含pTEV-PiGK5-N载体的菌落PCR扩增结果。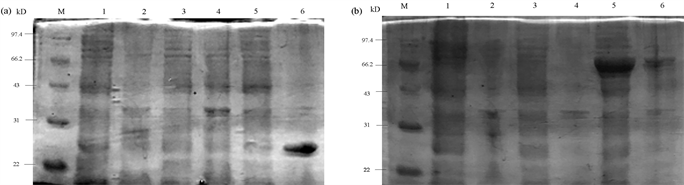 (a):重组蛋白PiGK5胞外端-His的诱导表达。(b):重组蛋白PiGK5胞外端-MBP-His的诱导表达。M:低分子量蛋白质Marker;1:BL21(DE3)菌株超声破碎后的上清液;2:BL21(DE3)菌株超声破碎后的菌体沉淀;A3:含空载体pET30a(+)的BL21(DE3)菌株超声破碎后的上清液;A4:含空载体pET30a(+)的BL21(DE3)菌株超声破碎后的菌体沉淀;A5:含载体pET30a(+)-PiGK5-N的BL21(DE3)菌株超声破碎后的上清液;A6:含载体pET30a(+)-PiGK5-N的BL21(DE3)菌株超声破碎后的菌体沉淀;B3:含空载体pTEV的BL21(DE3)菌株超声破碎后的上清液;B4:含空载体pTEV的BL21(DE3)菌株超声破碎后的菌体沉淀;B5:含载体pTEV-PiGK5-N的BL21(DE3)菌株超声破碎后的上清液;B6:含载体pTEV-PiGK5-N的BL21(DE3)菌株超声破碎后的菌体沉淀。
(a):重组蛋白PiGK5胞外端-His的诱导表达。(b):重组蛋白PiGK5胞外端-MBP-His的诱导表达。M:低分子量蛋白质Marker;1:BL21(DE3)菌株超声破碎后的上清液;2:BL21(DE3)菌株超声破碎后的菌体沉淀;A3:含空载体pET30a(+)的BL21(DE3)菌株超声破碎后的上清液;A4:含空载体pET30a(+)的BL21(DE3)菌株超声破碎后的菌体沉淀;A5:含载体pET30a(+)-PiGK5-N的BL21(DE3)菌株超声破碎后的上清液;A6:含载体pET30a(+)-PiGK5-N的BL21(DE3)菌株超声破碎后的菌体沉淀;B3:含空载体pTEV的BL21(DE3)菌株超声破碎后的上清液;B4:含空载体pTEV的BL21(DE3)菌株超声破碎后的菌体沉淀;B5:含载体pTEV-PiGK5-N的BL21(DE3)菌株超声破碎后的上清液;B6:含载体pTEV-PiGK5-N的BL21(DE3)菌株超声破碎后的菌体沉淀。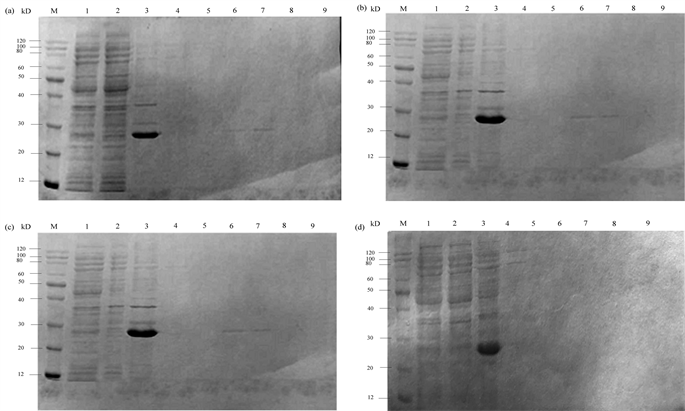 (a):100 μM IPTG诱导表达的PiGK5胞外端-His。(b):150 μM IPTG诱导表达的PiGK5胞外端-His。(c):200 μM IPTG诱导表达的PiGK5胞外端-His。(d):300 μM IPTG诱导表达的PiGK5胞外端-His。M:Protein Ruler II未染色蛋白Marker;1:菌体超声破碎离心后的上清;2:镍柱亲和层析后的穿透液;3:菌体超声破碎离心后的菌体沉淀;4:10 mM咪唑洗脱缓冲液的洗脱产物;5:20 mM咪唑洗脱缓冲液的洗脱产物;6:50 mM咪唑洗脱缓冲液的洗脱产物;7:100 mM咪唑洗脱缓冲液的洗脱产物;8:200 mM咪唑洗脱缓冲液的洗脱产物;9:500 mM咪唑洗脱缓冲液的洗脱产物。
(a):100 μM IPTG诱导表达的PiGK5胞外端-His。(b):150 μM IPTG诱导表达的PiGK5胞外端-His。(c):200 μM IPTG诱导表达的PiGK5胞外端-His。(d):300 μM IPTG诱导表达的PiGK5胞外端-His。M:Protein Ruler II未染色蛋白Marker;1:菌体超声破碎离心后的上清;2:镍柱亲和层析后的穿透液;3:菌体超声破碎离心后的菌体沉淀;4:10 mM咪唑洗脱缓冲液的洗脱产物;5:20 mM咪唑洗脱缓冲液的洗脱产物;6:50 mM咪唑洗脱缓冲液的洗脱产物;7:100 mM咪唑洗脱缓冲液的洗脱产物;8:200 mM咪唑洗脱缓冲液的洗脱产物;9:500 mM咪唑洗脱缓冲液的洗脱产物。 M:Protein Ruler II未染色蛋白Marker;1:菌体超声破碎离心后的上清;2:镍柱亲和层析后的穿透液;3:菌体超声破碎离心后的菌体沉淀;4:10 mM咪唑洗脱缓冲液的洗脱产物;5:20 mM咪唑洗脱缓冲液的洗脱产物;6:50 mM咪唑洗脱缓冲液的洗脱产物;7:100 mM咪唑洗脱缓冲液的洗脱产物;8:200 mM咪唑洗脱缓冲液的洗脱产物;9:500 mM咪唑洗脱缓冲液的洗脱产物。
M:Protein Ruler II未染色蛋白Marker;1:菌体超声破碎离心后的上清;2:镍柱亲和层析后的穿透液;3:菌体超声破碎离心后的菌体沉淀;4:10 mM咪唑洗脱缓冲液的洗脱产物;5:20 mM咪唑洗脱缓冲液的洗脱产物;6:50 mM咪唑洗脱缓冲液的洗脱产物;7:100 mM咪唑洗脱缓冲液的洗脱产物;8:200 mM咪唑洗脱缓冲液的洗脱产物;9:500 mM咪唑洗脱缓冲液的洗脱产物。
 (a):重组蛋白PiGK5胞外端-His的SDS-PAGE电泳。M:Protein Ruler II未染色蛋白Marker;1~8:第一个紫外吸收峰的第1~8号管蛋白洗脱液;9~11:第二个紫外吸收峰的1~3号管蛋白洗脱液。(b):重组蛋白PiGK5胞外端-MBP-His的SDS-PAGE电泳。M:Protein Ruler II未染色蛋白Marker;1~13:紫外吸收峰的第17~29号管蛋白洗脱液。
(a):重组蛋白PiGK5胞外端-His的SDS-PAGE电泳。M:Protein Ruler II未染色蛋白Marker;1~8:第一个紫外吸收峰的第1~8号管蛋白洗脱液;9~11:第二个紫外吸收峰的1~3号管蛋白洗脱液。(b):重组蛋白PiGK5胞外端-MBP-His的SDS-PAGE电泳。M:Protein Ruler II未染色蛋白Marker;1~13:紫外吸收峰的第17~29号管蛋白洗脱液。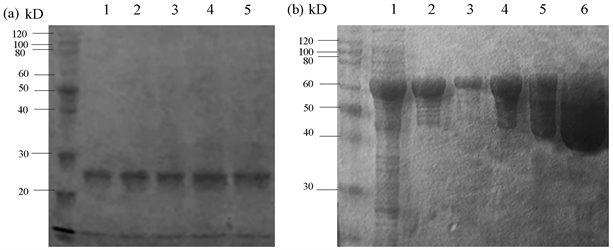 (a):重组蛋白PiGK5胞外端-His每24小时取样的SDS-PAGE结果。M:Protein Ruler II未染色蛋白Marker;1~5:重组蛋白PiGK5胞外端-His保存0 h、24 h、48 h、72 h、96 h时的电泳结果。(b):重组蛋白PiGK5胞外端-N-NBP-His每1.5小时取样的SDS-PAGE结果。M:Protein Ruler II;1:含PiGK5胞外端-MBP-His的菌体超声破碎离心后的上清液;2:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在0 h时取样;3:50 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在0 h时取样;4:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在1.5 h时取样;5:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在3 h时取样;6:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在4.5 h时取样。
(a):重组蛋白PiGK5胞外端-His每24小时取样的SDS-PAGE结果。M:Protein Ruler II未染色蛋白Marker;1~5:重组蛋白PiGK5胞外端-His保存0 h、24 h、48 h、72 h、96 h时的电泳结果。(b):重组蛋白PiGK5胞外端-N-NBP-His每1.5小时取样的SDS-PAGE结果。M:Protein Ruler II;1:含PiGK5胞外端-MBP-His的菌体超声破碎离心后的上清液;2:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在0 h时取样;3:50 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在0 h时取样;4:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在1.5 h时取样;5:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在3 h时取样;6:100 mM咪唑洗脱缓冲液洗脱的重组蛋白PiGK5胞外端-MBP-His在4.5 h时取样。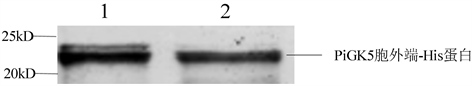 1、2:经镍柱纯化后的PiGK5胞外端-His蛋白。
1、2:经镍柱纯化后的PiGK5胞外端-His蛋白。
Effects of Different Tags on the Expression and Stability of Extracellular Terminal of Phytophthora infestans PiGK5[J]. 生物过程, 2022, 12(02): 40-53. https://doi.org/10.12677/BP.2022.122006