Botanical Research
Vol.3 No.02(2014), Article ID:13263,15 pages
DOI:10.12677/BR.2014.32009
茜草科7种木质藤本茎部异常构造
Sheng-Zehn Yang*, Po-Hau Chen, Hsuan Fan, Kuan-Wei Li, Yueh-Feng ChenDepartment of Forestry, National Pingtung University of Science and Technology, Pingtung County, Taiwan
Email: *yangsz@mail.npust.edu.tw
Copyright © 2014 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received: Jan. 1st, 2014; revised: Feb. 3rd, 2014; accepted: Feb. 14th, 2014
ABSTRACT
Rubiaceae are one of the dominant families in the broad leaved forest of Taiwan in low altitude, including some lianas. There was a paucity of the cambium variations of lianas in this family that affects the lianas identification correctly. The aims of this study are to identify the cambium variation of Rubiaceae seven liana stems, line drawings, photo the cross-section of stem structures, and make an indented key in order to aid the field survey. Four xylem types are found, including deeply lobe types such as Hedyotis hedyotidea, Morinda parvifolia and Morinda umbellate; twolobed type as Paederia foetida; circular type as Mussaenda pubescens, Psychotria serpens, and Uncaria lanosa var. appendiculata belonging to angular type. Except Paederia foetida, six species develop including phloem. The specific character of Uncaria lanosa var. appendiculata about included phloem is square shape. Psychotria serpens develop adventitious root, and Morinda parvifolia and Morinda umbellata were together with thick phellem. Four species with flame secondary phloem are Hedyotis hedyotidea, Paederia foetida, Mussaenda pubescens, and Psychotria serpens. Secondary xylem and secondary phloem developing mosaic pattern are Hedyotis hedyotidea, Morinda parvifolia and Morinda umbellata. Only one species with flame secondary phloem and mosaic pattern is Hedyotis hedyotidea in which its pith is eccentric.
Keywords:Anomalous Structure; Lianas; Rubiaceae; Secondary Phloem

茜草科7种木质藤本茎部异常构造
杨胜任*,陈柏豪,范 轩,李冠纬,陈月凤
国立屏东科技大学森林系,台湾屏东县
Email: *yangsz@mail.npust.edu.tw
收稿日期:2014年1月1日;修回日期:2014年2月3日;录用日期:2014年2月14日

摘 要
茜草科是台湾低海拔次生林及阔叶林之优势科之一,缺乏木质藤本形成层变异资料。本研究目的探讨茜草科7种木质藤本茎形成层变异,绘制拍照横切面构造图、制作检索表以及描述茎部各部位构造,供野外采集调查及鉴定上之参考。本文归类出4种木质部形态,即深裂状有南投凉喉茶、红珠藤、羊角藤;两浅裂状为鸡屎藤;毛玉叶金花、拎壁龙为圆形;恒春钩藤为角状。除鸡屎藤外,其余6种均具内含韧皮部,恒春钩藤内含韧皮部四方形最为特殊。拎壁龙具不定根。红珠藤和羊角藤具木栓层构造。南投凉喉茶、鸡屎藤、毛玉叶金花、拎壁龙具火焰状次生韧皮部;次生韧皮部与次生木质部镶嵌在一起为南投凉喉茶、红珠藤和羊角藤。南投凉喉茶是唯一具有火焰状次生韧皮部、次生韧皮部与木质部镶嵌等2种特征且髓心偏一侧。
关键词
茜草科;木质藤本;异常构造;次生韧皮部

1. 引言
台湾维管束植物依不同生长形态,区分为树木、灌木、木质藤本(lianas)、草质藤本、草本、着生植物、寄生植物和腐生植物,其中木质藤本约6.9%。所有科中木质藤本最多者为豆科(Leguminosae)24种、葡萄科(Vitaceae)21种和蔷薇科(Rosaceae)19种[1] 。木质藤本通常具较粗且木质的茎,幼苗生长于成熟的森林[2] 。木本植物与藤本植物除外观上差异,在内部构造方面也是有很大的不同。许多藤本植物内部构造起源,只有束状形成层产生典型的次生木质部和次生韧皮部,而束间形成层产生次生薄壁组织[3] 。木质部对于植物本身是具有支持及运输水分功能之组织,不论是对于一般木本植物或是木质藤本都相当重要。木本植物之木质部形态轮廓通常为圆形,但在藤本植物茎横切面上木质部多变,此为木质部各区域不均等生长所导致。维管束形成层向内产生木质部,向外产生韧皮部,各区域分化出木质部具比例的不同[4] 。横切面中某些区域次生木质部会大量产生,并将维管束形成层向外推挤;而邻近区域韧皮部组织比木质部产生较多,使形成层向外推挤变缓慢,因此造成木质部不规则状轮廓出现。
茎横切面的描述主要是根据木质部的特征,特别是连续性或不连续性及轮廓,其代表生长的连续过程中不同状况或不同阶段,与藤本的次级生长有关[3] 。维管束形成层活动的变异,无论是特定部位组织产生性质或数量变化,或在形成层不同区域细胞分裂的频率变化,均会产生木质部和韧皮部不正常的分布模式,或不同形状的茎,其中茎的外部很大部分是次生木质部的形状。许多热带藤本的特征在于异常的茎结构,但这种结构并不限制于热带藤本[3] 。
Caballé[5] 将茎横切面与木质部形态分类为12种,分别为圆形(circular)、弯曲或平面(sinuate or plane)、波浪状(crenate)、角状(angular)、分裂状(lobed)、放射状(rays)、年轮状(rings)、年轮状和放射状(rings and rays)、块状(blocks)及四分裂(quarter)、复合木质部圆柱状(duplicated xylem cylinders)、股状内含韧皮部(included phloem strands)。
Hawthorne等[6] 依大型木质藤本茎与较小茎特征分别描述木质藤本正常与异常构造,并辅以照片说明不同科别的形成层变异。例如木质藤本具正常木质,或具不规则多孔导管(lacunose anomalies),散播在圆柱状茎,如茜草科玉心花属(Tarenna),钩藤属(Uncaria);木质茎具明显辐射状变化(radial pattern),可清楚看到射线(rays)或辐射状内含韧皮部(radial lines of included phloem),或木质部星状深裂或沟渠状 (deeply lobed star-shaped, sulcate xylem);茎具同心圆或似波纹状年轮曲线(concentric or ripple-like annular curvy pattern);茎圆形(rounded),但常为扁平状(flat)或不规则裂(irregularly lobed),或具非星状深裂(deep lobes and not star shaped)。
Isnard等[7] 述及木质藤本7种异常构造,所谓异常构造是源自于不规则形成层以及增生(supernumerary)形成层变异。例如圆形构造或分裂状形成层(lobed cambium);不规则形成层(irregular cambium),是因为形成层有部分区域往侧面发展,使得横切面的次生木质部少于次生韧皮部;多数形成层产生为多元分裂状形成层(multiple and lobed cambia)变异;多数形成层产生黑点状的内含韧皮部(interxylary phloem);同心圆形成层(concentric cambia);连续性有方向形成层(successive and directional cambia)。
Angyalossy等[8] 则分成内含韧皮部、弧形/楔形韧皮部(phloem arcs/wedges)、连续形成层(successive cambia)、复维管束圆柱体(compound vascular cylinder)、茎不规则结构(stem with irregular conformation)、维管束成分纵向分段(axial vascular elements in segments)、薄壁组织分裂分散木质部(xylem dispersed by parenchyma divisions)、外部次生圆柱体(external secondary cylinders)等8种形态。
台湾中低海拔广泛分布茜草科(Rubiaceae)藤本植物,多喜爱阳光,依其不同攀爬机制,攀爬至树冠顶端,叶和花部常生长在相当高之树冠层,并与其它植物混合生长而不易辨识,导致研究人员在野外采集时调查及鉴定上之困难,若能从横切面鉴定这些物种,将有助于研究。杨胜任等[9] 说明夹竹桃科及萝摩科12种木质藤本茎部异常构造。有关茜草科木质藤本横切面异常构造在国内外研究较少,Metcalfe等[10] 茜草科茎部横切面与木质部等5种异常构造,包括1) 茎具内含韧皮部,如Basanacantha asperifolia,Chiococca,Myrmecodia等3属植物(台湾未见分布)。2) 木质部和韧皮部同心圆交替排列(concentrically alternating xylem and phloem),如Chiococca属。3) 茎偏心至卵形或椭圆形(stems eccentric to oval or elliptical),如Manettia,Sabicea (台湾无此2属植物)。4) 茎沟状或分裂状(stems furrowed or lobed),如Canthium (鱼骨木属) (台湾未见分布),Mussaenda chippii,M. tristigamatica (玉叶金花属)。5) 木质部沟状(xylem furrowed),如Chiococca,Manettia,Sabicea,鸡屎树属(Paederia)。Caballé[5] 提及Canthium venosum (鱼骨木属)为4分裂(quarter),非洲钩藤(Uncaria africana)为圆形,髓心明显居中,并统计非洲17种茜草科植物木质部圆柱体横切面有6种形态(分裂状、分裂状–复合木质部、波浪状–角状、波浪状–角状–复合木质部、弯曲或平面、4分裂–分裂状–射线);美洲3种茜草科植物有3种形态(弯曲或平面、波浪状–角状、4分裂–分裂状–射线)。
本研究目的为藉由判释木质藤本横切面异常构造及相关术语作为鉴定藤本植物之依据,利用横切面构造来分类茜草科的木质藤本,并将其横切面构造进行描述、比较、制作不齐头检索表(indented key)、绘制横切面图及拍照,并指出每一物种横切面上各部位之名称,供日后之研究者野外采集调查及鉴定上之参考。
2. 研究材料与方法
木质藤本样本大多采集于成年植株,同一物种重复采集观察不同粗细的横切面构造是否异同,供比对分析之用。将采集回来的新鲜材料切段约5 cm长,使用标签纸及透明胶带进行标示。以徒手将横切面削切平整,并以放大20倍率清晰看出外观构造,供解剖显微镜(型号Stemi 2000-c,Carl Zeiss)观察并绘制构造图,再将材料置于翻拍架使用Nikon D80单眼相机(镜头AF MICRO NIKKOR 60 mm 1:2.8D)拍摄藤本横切面,及拍摄照片后,将标本送入60℃烘箱烘干及低温杀虫后制成干燥标本,存放在国立屏东科技大学植物标本馆(PPI),供后续鉴定之用。取木质藤本约15~20 cm烘干供黏贴制作标本。制作完整的干燥标本亦归档于标本馆中。依照Flora of Taiwan第二版[11] [12] 记录学名以及PPI标本馆标本做确认。
本研究于2011年08月到2012年09月至台湾中、南部采集7种木质藤本,简述如下:1) 南投凉喉茶(Hedyotis hedyotidea (DC.) Merr.)采集地点:南投,海拔500~700 m,标本直径大小约1.9 cm。2) 红珠藤(Morinda parvifolia Bartl.)采集地点:台中县龙井乡,海拔200~300 m,标本直径大小约1.3 cm。3) 羊角藤(Morinda umbellata L.)采集地点:台东县达仁乡达仁林场,邻接屏东县,海拔约180~900 m;牡丹乡的恒春23林班,海拔210~490 m,标本直径大小约2.6 cm。4) 毛玉叶金花(Mussaenda pubescens W. T. Aiton) 采集地点:台东县达仁乡达仁林场;牡丹乡的恒春23林班;南投中寮乡,海拔500~700 m;屏东真笠山,海拔1166 m,标本直径大小约2.5 cm。5) 鸡屎藤(Paederia foetida L.)采集地点:南投中寮乡,标本直径大小约1.5 cm。6) 拎壁龙(Psychotria serpens L.)采集地点:台东县达仁乡达仁林场,标本直径大小约1.8 cm。7) 恒春钩藤(Uncaria lanosa Wall. var. appendiculata (Benth.) Ridsdale) 采集地点:牡丹乡恒春23林班,标本直径大小约3.3 cm。
本文参考 Metcalfe等[10] ,Caballé[5] ,Carlquist[13] [14] ,Isnard等[7] ,Angyalossy等[8] 等所绘制的图示做比对,将茎横切面形成层变异归类成16项(表1),以提供本文描述之依据。木质藤本的重要术语依据蔡淑华[15] 、刘业经等[16] 、Mauseth[4] 、Metcalfe等[10] 等学者描述藤本所用的定义(如附录)。
3. 结果
3.1. 茜草科7种木质藤本植物茎部构造特征描述
1) 南投凉喉茶(图1):茎非正圆,不规则,无附属物,质地软易砍断;次生木质部深裂状,次生木质部内具导管不规则分布,与次生韧皮部镶嵌一起,次生木质部外侧亦具火焰状次生韧皮部,次生木质部大小分布不均;髓心黑色,偏一侧,椭圆形,外侧具内含韧皮部。
2) 红珠藤(图2):茎圆形或波浪形,具木栓层构造;次生木质部深裂状,几深达髓心,无射线,次生木质部导管不规则分布,无年轮状,与次生韧皮部镶嵌一起;髓心深黑色,位于中心,圆形,近髓心具内含韧皮部。
3) 羊角藤(图3):茎圆形或波浪状,具木栓层构造;次生木质部深裂状,几深达髓心,深裂状次生木质部先端又细分成2裂,如Y形,次生木质部导管不规则分布,与次生韧皮部镶嵌一起;髓心圆形,深黑色。小茎近髓心具内含韧皮部,大茎则不明显。
4) 毛玉叶金花(图4):茎明显圆柱状,表皮无附属物;横剖面具射线,多而密,导管略呈年轮状排列;次生木质部外侧具火焰状构造的次生韧皮部,干燥后呈黑色;髓心白色,圆形,近髓心具内含韧皮部。
5) 鸡屎藤(图5):茎通常两裂,大小不一,表皮略白,无附属物,皮层明显呈绿色,老茎有疣状突起,茎质地软易砍断;横剖面具明显射线,两裂凹陷处射线密集非常明显,次生木质部两裂状,导管不规则分布且密,近髓心处导管较小;次生木质部外侧具次生韧皮部,呈火焰状构造;髓心明显,长椭圆形,随茎的增大而逐渐变小,近髓心无内含韧皮部。
6) 拎壁龙(图6):茎具不定根,圆形,质地硬;横剖面具具明显射线,多而密,次生木质部内导管不规则分布;次生木质部外侧具浅黑色次生韧皮部,呈火焰状构造,干燥后呈黑色;髓心色黑,圆形,近髓心具内含韧皮部。
7) 恒春钩藤(图7):幼茎四方形,老茎略圆形,干燥后波浪状或角状,表皮无附属物;横剖面不具明显射线,次生木质部角状,导管不规则分布,次生韧皮部新鲜时浅黄棕色,干燥后呈黑色,周围韧皮部(peripheral phloem) (包括次生韧皮部与表皮、皮层)厚度非常明显,深黄褐色;髓心白色,圆形,内含

Table 1. Integrated table of morphological classification of stem cross-section
表1. 茎横切面形态分类整合表
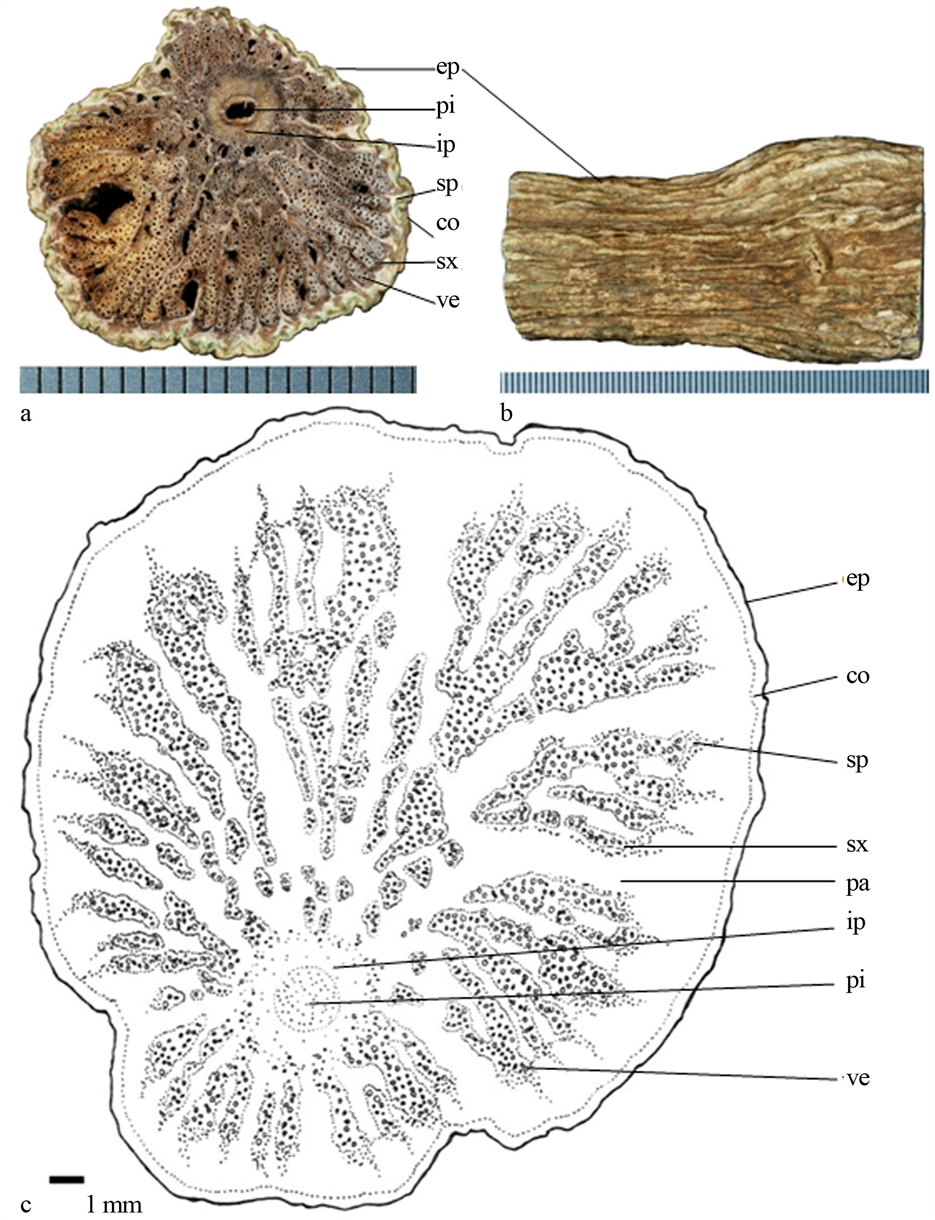
Figure 1. (a) Transverse section photo of Hedyotis hedyotidea, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ep (epidermis表皮);co (cortex皮层);sp (secondary phloem次生韧皮部);pa (parenchyma 薄壁细胞);sx (secondary xylem次生木质部);ip (included phloem内含韧皮部);pi (pith髓心);ve (vessels导管)
图1. (a) 南投凉喉茶(Hedyotis hedyotidea)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图
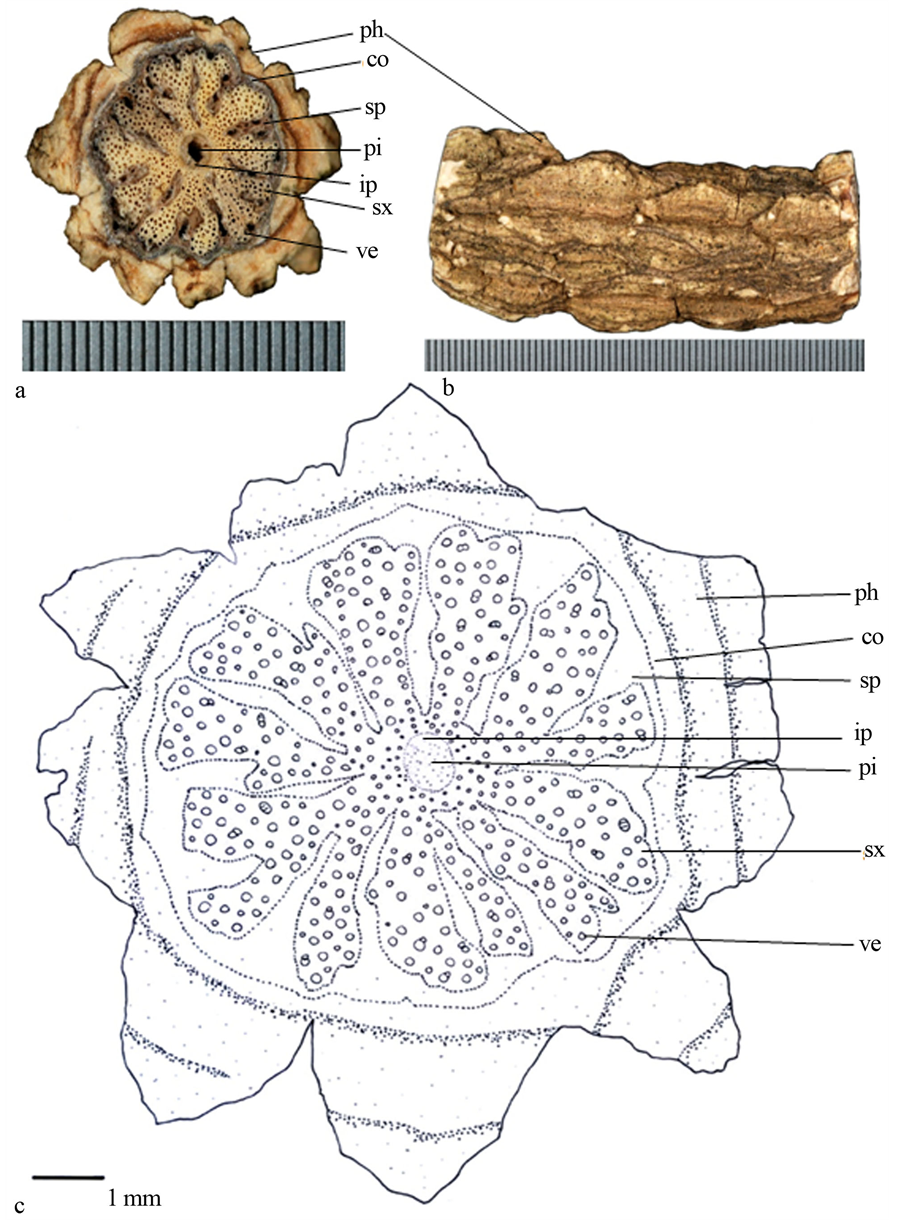
Figure 2. (a) Transverse section photo of Morinda parvifolia, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ph (phellem木栓层);co (cortex皮层);sp (secondary phloem次生韧皮部);ip (included phloem内含韧皮部);pi (pith髓心);sx (secondary xylem次生木质部);ve (vessels导管)
图2. (a) 红珠藤(Morinda parvifolia )横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图
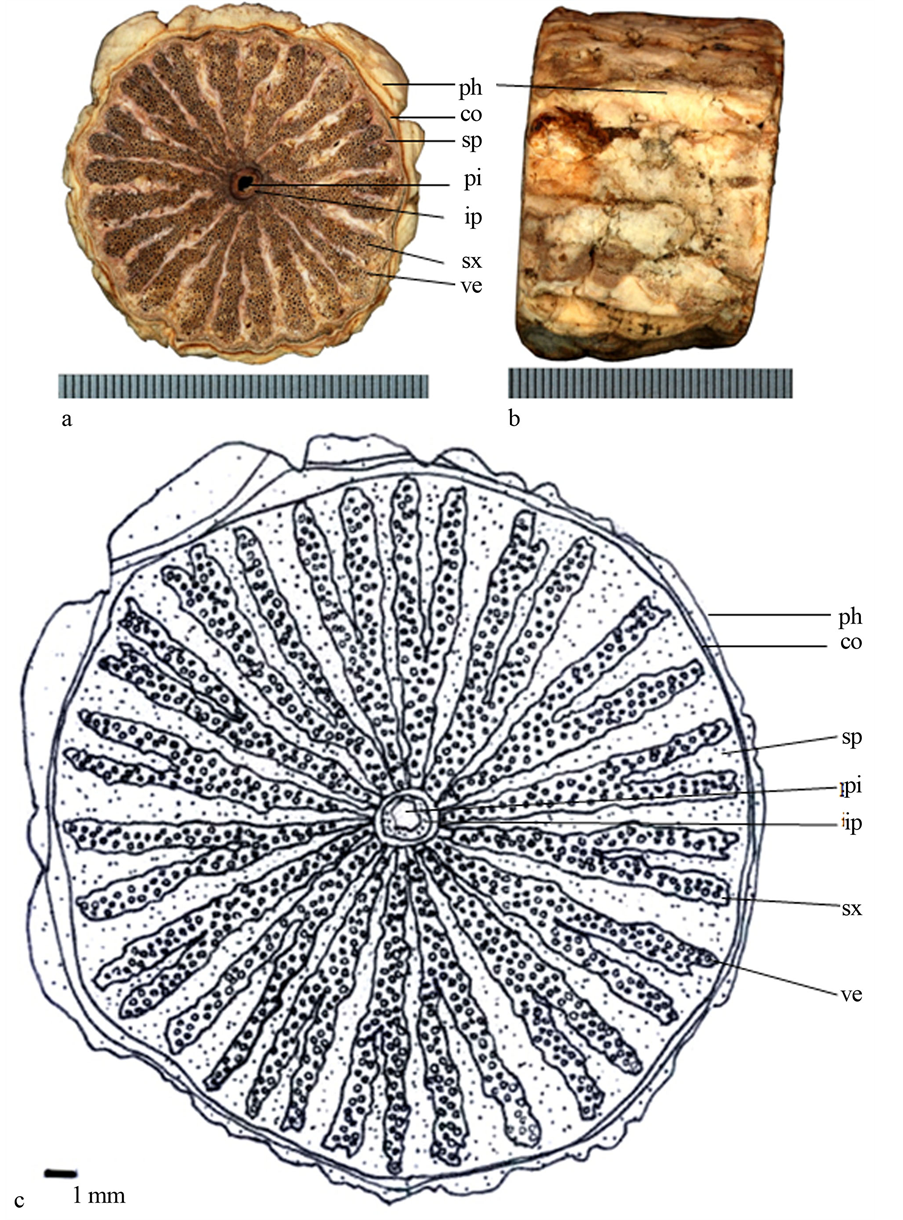
Figure 3. (a) Transverse section photo of Morinda umbellata, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ph (phellem木栓层);co (cortex皮层);sp (secondary phloem次生韧皮部);pi (pith髓心);ip (included phloem内含韧皮部);sx (secondary xylem次生木质部);ve (vessels导管)
图3. (a) 羊角藤(Morinda umbellata)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图
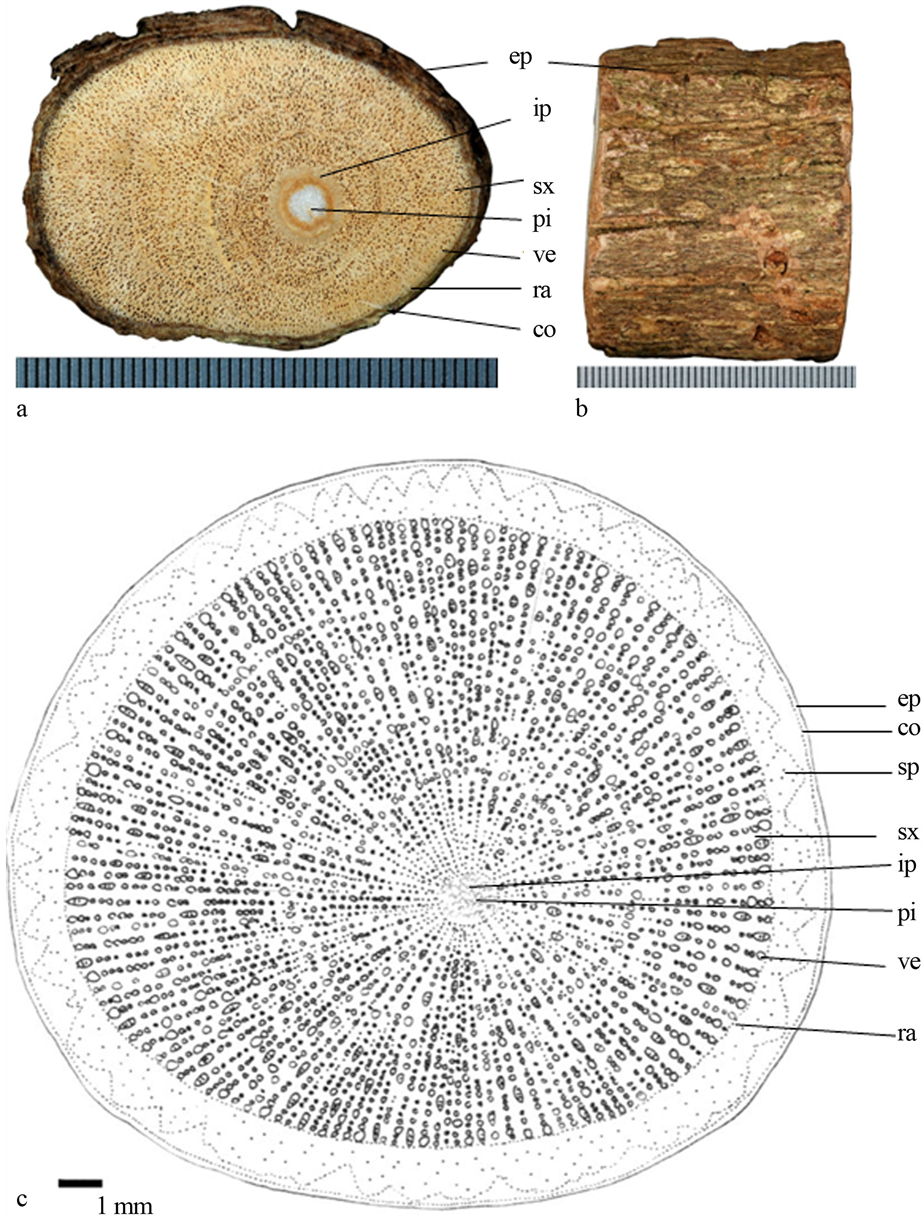
Figure 4. (a) Transverse section photo of Mussaenda pubescens, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ep (epidermis表皮);co (cortex皮层);sp (secondary phloem次生韧皮部);sx (secondary xylem次生木质部);ip (included phloem内含韧皮部);pi (pith髓心);ve (vessels导管);ra (ray射线)
图4. (a) 毛玉叶金花(Mussaenda pubescens)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图
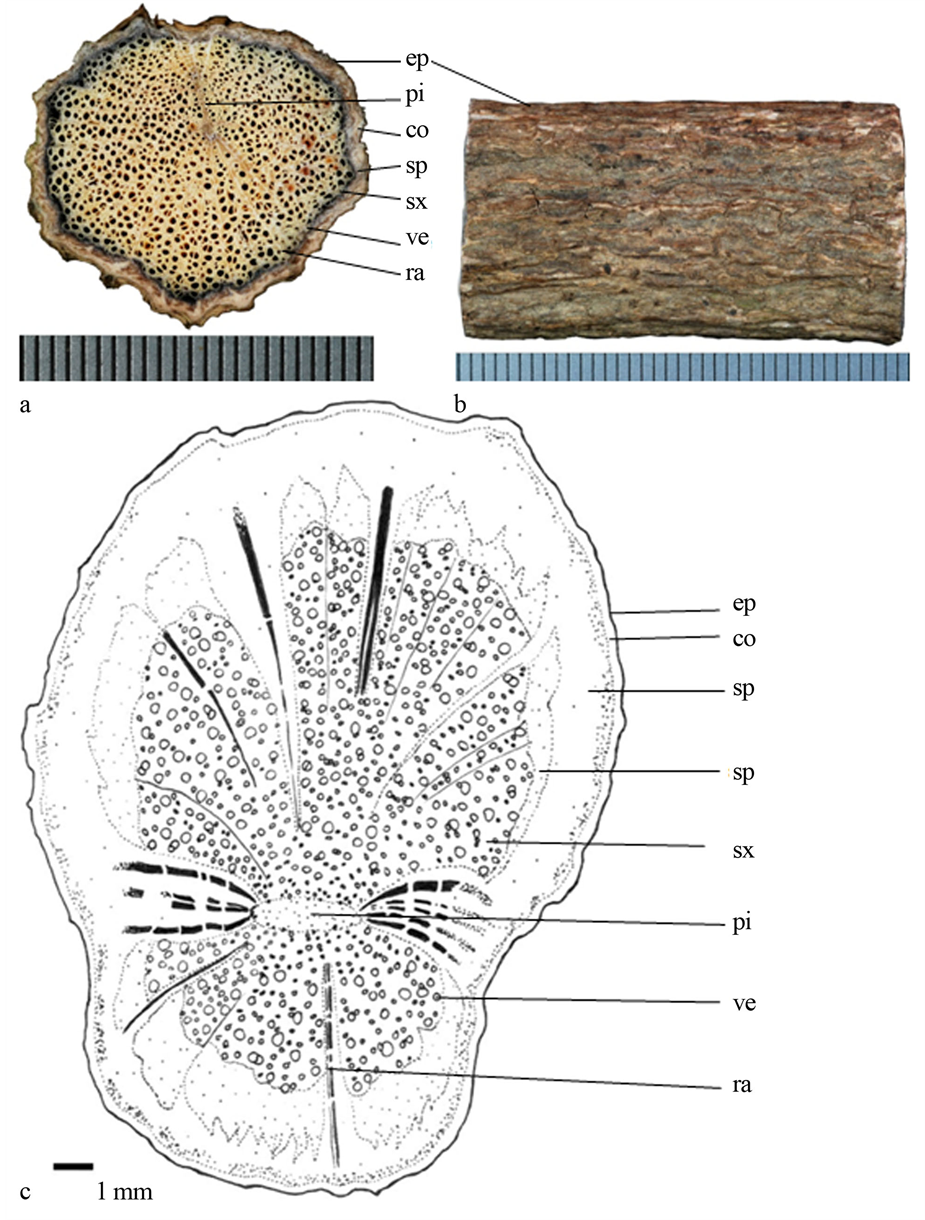
Figure 5. (a) Transverse section photo of Paederia foetida, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ep (epidermis表皮);co (cortex皮层);sp (secondary phloem次生韧皮部);sx (secondary xylem次生木质部);pi (pith髓心);ve (vessels导管);ra (ray射线)
图5. (a) 鸡屎藤(Paederia foetida)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图

Figure 6. (a) Transverse section photo of Psychotria serpens, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ep (epidermis表皮);co (cortex皮层);sp (secondary phloem次生韧皮部);ip (included phloem内含韧皮部);pi (pith髓心);sx (secondary xylem次生木质部);ve (vessels导管);ra (ray射线);ad (adventitious root不定根)
图6. (a) 拎壁龙(Psychotria serpens)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图

Figure 7. (a) Transverse section photo of Uncaria lanosa var. appendiculata, scale = 0.5 mm; (b) Epidermis of stem; (c) Line-drawing
ep (epidermis表皮);pi (pith髓心);ip (included phloem内含韧皮部);co (cortex皮层);sp (secondary phloem次生韧皮部);sx (secondary xylem次生木质部);ve (vessels导管)
图7. (a) 恒春钩藤(Uncaria lanosa var. appendiculata)横切面照片,刻度 = 0.5 mm;(b) 表皮照片;(c) 手绘图
韧皮部呈四方形。
3.2. 茜草科7种木质藤本检索表
依据7种木质藤本横切面各项结构特征归纳出如表2,重要特征包括不定根、木栓层、茎部、次生木质部、射线、髓心、内含韧皮部、次生韧皮部、导管分布。导管分布分为两类,一为不规则分布,二为年轮状分布。次生韧皮部分为火焰状、与木质部镶嵌状。拎壁龙具不定根,表皮无附属物;红珠藤和羊角藤具木栓层构造;次生韧皮部火焰状有南投凉喉茶、鸡屎藤与毛玉叶金花、拎壁龙;次生韧皮部与木质部镶嵌在一起为南投凉喉茶、红珠藤和羊角藤;恒春钩藤无明显次生韧皮部构造。由上述各项横切面结构特征(表2),制成7种木质藤本不齐头检索表。
1.木质部分裂状,髓心黑色。
2.不具木栓层,髓心偏心-------------------------------------------------南投凉喉茶
2.具木栓层,髓心居中心。
3.木栓层浅色----------------------------------------------------------------羊角藤
3.木栓层棕色----------------------------------------------------------------红珠藤
1.木质部圆形状。
4.茎具不定根----------------------------------------------------------------拎壁龙
4.茎不具不定根。
5.近髓心不具内含韧皮部-------------------------------------------------鸡屎藤
5.近髓心具内含韧皮部。
6.内含韧皮部圆形,具射线----------------------------------------------毛玉叶金花
6.内含韧皮部四方形,不具射线----------------------------------------恒春钩藤
4. 讨论
Metcalfe等[10] 描述茜草科有5种异常现象,包括1) 茎具内含韧皮部。2) 木质部和韧皮部同心圆交替排列。3) 茎偏心至卵形或椭圆形。4) 茎沟状或分裂状。5) 木质部沟状等。本研究结果有3种异常构造,包括茎具内含韧皮部、茎沟状或分裂状、木质部分裂状(表1)。本文所称木质部分裂状即与木质部沟状相同,均指次生木质部的构造。除鸡屎藤外,其余6种均具内含韧皮部(included phloem),此些特征与Metcalfe等[10] 描述相同。南投凉喉茶茎髓心偏一侧,鸡屎藤茎横切面呈2浅裂状,如上述第第3至5点相同,茎偏心且木质部沟状。
Mauseth[4] 指出形成层各区域分化出不同比例的木质部,羊角藤和红珠藤的木质部有不均等生长之木质部,深裂状木质部于末端常细分成二裂状,二者无明显区别之特征,唯一较明显差异特征为木栓层色泽之不同,本研究则以木栓层的色泽做为区别红珠藤和羊角藤之重要特征。具有不定根之木质藤本为拎壁龙,与鸡屎藤、毛玉叶金花皆有明显射线。导管年轮状排列较明显者有玉叶金花和恒春钩藤,两者差异之处为毛玉叶金花具有射线,恒春钩藤则无射线。研究材料中次生韧皮部形状不一,具火焰状次生韧皮部者为南投凉喉茶、鸡屎藤、毛玉叶金花、拎壁龙;次生韧皮部与木质部镶嵌一起,如红珠藤和羊角藤。南投凉喉茶是唯一具有火焰状次生韧皮部、次生韧皮部与木质部镶嵌等2种特征。与次生木质部镶嵌一起的次生韧皮部,Metcalfe等[10] 亦称之为由薄壁细胞扩展(dilation of parenchyma)而来。
Hawthorne等[6] 表示茜草科针刺性状亦可用来区别藤本植物,其特殊表征为枝条硬有固定长度,与茎呈垂直,如玉心花属;枝条硬且规则着生,具短螺旋状勾状物(short spiral hooks)或卷曲勾状物(coiled hooks, hook-like thorn),如玉叶金花属(Mussaenda)、钩藤属;针刺非着生在表皮而是源自于枝条,与茎的
Table 2. The characters of stem cross-section about seven lianas of Rubiaceae
表2. 茜草科7种木质藤本横切面特征
木质连接,似针状,如九节木属(Psychotria);茎、叶柄与卷须(twining stems, petioles, tendrils)具缠绕性,如具正常的主茎缠绕;茎产生不定根(adventitious roots from the stems)。陈建帆等[17] 以植物攀爬方式分出8种类型,其中恒春钩藤具卷曲勾状物,为钩刺依附(scrambling by hooks),拎壁龙具不定根,为不定根吸附(adhesive adventitious root),等。毛玉叶金花、拎壁龙、南投凉喉茶、红珠藤、羊角藤等5种为主茎缠绕(twining stem)。本研究物种中针刺构造与攀缘方式确实可辅助物种的鉴定。
Lindorf[18] 研究钩藤花草(Uncaria guianensis)与绒毛钩藤(U. tomentosa) (俗称猫爪Cats claw)的树皮与木材解剖,是南美民族重要的民间医药药材,因为经常卖掺假与其他物种,才有本文之研究。Gattuso等[19] 研究钩藤花草与绒毛钩藤的树皮与叶成分。钩藤属广泛分布在热带地区的物种。两个原生种的叶子“猫爪”是民间中医使用治疗药物。本文进行比较形态解剖显微特征,韧皮纤维(phloem fibers),草酸钙晶体(calcium oxalate crystals),淀粉颗粒(starch granules),皮毛(trichomes)。本属于台湾有3种植物,从药学观点有3篇研究论文,如徐列庆[20] 研究台湾产钩藤对抗忧郁作用比较研究;周俊孝[21] 从神经科学研究钩藤碱功能性;张婷翔[22] 研究台湾产钓藤生药学研究,然从形成层变异特征仍未有相关报告,实值得进行比较研究。
本研究采集之木质藤本,属于生活史中某一阶段横切面构造,木质藤本次级生长会随生长阶段不同而导致内部构造变化,所以采集不同阶段之木质藤本,进行内部构造比较及探讨,实有需做更进一步研究。
5. 结论
本研究将7种木质藤本归类出4种木质部形态,即深裂状有南投凉喉茶、红珠藤、羊角藤;二浅裂状为鸡屎藤;毛玉叶金花、拎壁龙为圆形;恒春钩藤为角状。除鸡屎藤外,其余6种均具内含韧皮部。7种木质藤本仅拎壁龙具不定根,表皮无附属物。红珠藤和羊角藤具木栓层构造,以木栓层色泽做为区别。南投凉喉茶、鸡屎藤、毛玉叶金花、拎壁龙具火焰状次生韧皮部;次生韧皮部与次生木质部镶嵌在一起为南投凉喉茶、红珠藤和羊角藤;恒春钩藤无明显次生韧皮部构造。南投凉喉茶是唯一具有火焰状次生韧皮部、次生韧皮部与木质部镶嵌等2种特征。南投凉喉茶茎髓心偏一侧。恒春钩藤的内含韧皮部四方形最为特殊。
参考文献 (References)
- Hsieh, C.F. (2003) Composition, endemism and phytogeographical affinities of Taiwan flora. In: Huang, T.C., Ed., Flora of Taiwan, 2nd Edition, Vol. 6, 1-14.
- Gentry, A.H. (1985) Anecotaxonomic survey of Panamanian lianas. In: Darcy, W.G. and Correa, M.D., Eds., Historia Natural de Panama, Monographs of Systematic Botany, Missouri Botanical Garden, Saint Louis.
- Beck, C.B. (2011) An Introduction to plant structure and development: Plant anatomy for the twenty-first century. 2nd Edition, Cambridge University Press, Cambridge.
- Mauseth, J.D. (1988) Plant anatomy.
- Caballé, G. (1993) Liana structure, function and selection: A comparative study of xylem cylinders of tropical rainforest species in Africa and America. Botanical Journal of the Linnean Society, 113, 41-60.
- Hawthorne, W.D. and Jongkind, C.C.H. (2006) The woody plants of western African forests: A guide to the forest tree, shrubs and lianas from Senegal to Ghana. Royal Botanic Gardens, Kew.
- Isnard, S. and Silk, W.K. (2009) Moving with climbing plants from Charles Darwin’s time into the 21st century. American Journal of Botany, 96, 1205-1221.
- Angyalossy, V., Angeles, G., Marcelo, R.P., Lima, A.C., Dias-Leme, C.L., Lohmann, L.G. and Madero-Vega, C. (2012) An overview on the anatomy, development and evolution of the vascular system of lianas. Plant Ecology & Diversity, 5, 167-182.
- 杨胜任, 陈建帆, 赖冠伶 (2013) 夹竹桃科及萝摩科12种木质藤本茎部异常构造. 国立台湾博物馆学刊, 66, 57-86.
- Metcalfe, C.R. and Chalk, L. (1985) Anatomy of the dicotyledons: Volume II: Wood structure and conclusion of the general introduction. 2nd Edition, Oxford University Press, New York.
- Bufford, D.E., Chang, C.E., Chaw, S.M., Chen, C.H., Deng, M.J., Hartley, T.C., Hoch, P.C., Hsieh, C.F., Hsieh, T.H., Huang, S.F., Huang, T.C., Kao, M.T., Li, H.L., Lievens, A.W., Lin, S.C., Liu, H.Y., Liu, T.S., Liu, Y.C., Lo, H.C., Lu, F.Y., Lu, S.Y., Ohashi, H., Ou, C.H., Peng, C.I., Raven, P.H., Tang, W.S., Wagner, W.L., Wang, C.M., Wang, J.C., Yang, A.T.Y., Yang, K.C. and Yang, Y.P. (1993) Flora of Taiwan. 2nd Edition, Vol. 3, Editorial Committee, Department of Botany, Nationl Taiwan University, Taipei.
- Boufford, D.E., Hsien, C.F., Huang, T.C., Lowry, P.P.H., Ohashi, H. and Peng, C.I. (1998) Flora of Taiwan. 2nd Edition, Vol. 4, Editorial Committee, Department of Botany, National Taiwan University, Taipei.
- Carlquist, S. (1991) 2. Anatomy of vine and liana stems: A review and synthesis. In: Putz, F.E. and Mooney, H.A., Eds., The Biology of Vines, Cambridge University Press, Cambridge, 53-71.
- Carlquist, S. (2001) 8. Cambial variants (anomalous secondary growth). In: Carlquist, S., Ed., Comparative Wood Anatomy, Systematic, Ecological and Evolutionary Aspects of Dicotyledon Wood, Springer-Verlag, Berlin, 271-295.
- 蔡淑华 (1973) 植物解剖学(全1册). 国立编译馆, 台北.
- 刘业经, 吕福原, 欧辰雄 (1994) 台湾树木志. 国立中兴大学农学院出版, 台中.
- 陈建帆, 陈柏豪, 杨胜任 (2013) 台湾中南部地区藤本植物的攀爬类型. 中华林学季刊, 46, 441-460.
- Lindorf, H. (2005) Bark and wood anatomy of (Uncaria guianensis) and Uncaria tomentosa (cat’s claw). IAWA Journal, 26, 239-251.
- Gattuso, M., Sapio1, Di, O., Gattuso, S. and Pereyra, E.L. (2004) Morphoanatomical studies of Uncaria tomentosa and Uncaria guianensis bark and leaves. Phytomedicine, 11, 213-223.
- 徐列庆 (2011) 台湾产及市售钩藤对小鼠忧郁模式之抗忧郁作用比较研究. 硕士论文, 中国药学暨中药资源学系, 台中.
- 周俊孝 (2009) 钩藤碱功能性地转换延迟整流型钾离子通道成为A型钾离子通道. 硕士论文, 中国医药大学神经科学与认知科学研究所, 台中.
- 张婷翔 (2009) 台湾产钓藤(钩藤)之生药学研究. 硕士论文, 中国医药大学中国药学研究所, 台中.

附录:木质藤本的重要术语
1) 不定根(adventitious root):自植物的叶或茎上长出来,而不是由胚根发育成的根,此由不正常的部位长出来的根称为不定根。
2) 皮层(cortex):位于表层至维管束之间的组织,最简单的皮层只含有薄壁细胞。
3) 表皮(epidermis):即指覆盖于除了顶端分生组织以外的所有初生植物体最外层的一至数层细胞,又称表层组织。
4) 周皮(periderm):位于皮层外围,为木栓形成层(phellogen)、木栓层(phellem)和栓皮层(phelloderm)所构成,并与初生组织和/或次生韧皮部合称为树皮(bark),亦称为周围韧皮部(peripheral phloem)。
5) 木栓层(phellem):位于皮层外围,并包含于周皮中。茎中木栓层在奠定下来后会取代表皮组织,栓化细胞壁防止养分到细胞外。木栓层在茎中为次生保护组织,由死细胞所构成栓化的墙,几乎由软木形成层或木栓形成层所产生的。
6) 韧皮部(phloem):由筛细胞、伴细胞、射线筛管和纤维所构成之复合组织,负责运送养分供给植物体。韧皮部又分成初生韧皮部与次生韧皮部。次生韧皮部横行方向之射线薄壁细胞和木质部之射线相连,且大小相似,但位于外围(较老韧皮部)之一些韧皮射线可能会加宽,这是因为树干圆周加大时射线即开始垂周分裂(anticlinal division)以增加射线宽度。韧皮射线的加宽,使次生韧皮部彼此间的距离加大,并向外形成火焰状。
7) 髓心(pith):通常位于茎部中央,大部分由薄壁细胞所组成,为分化程度最低的组织。
8) 射线(ray):髓心部和皮层相连的薄壁组织。
9) 次生韧皮部(secondary phloem):位于木质部与皮层间,初生韧皮部形成后,由维管束形成层分裂,分化而成之韧皮部,包含筛细胞、韧皮纤维和韧皮薄壁细胞。
10) 次生木质部(secondary xylem):位于髓心与韧皮部间,初生木质部形成后,由维管束形成层分裂、分化而成的木质部。
11) 导管(vessels):位于被子植物木质部内,由导管细胞相连而成之整个细胞列称之为导管,具有穿孔。
NOTES
*通讯作者。