Botanical Research
Vol.07 No.04(2018), Article ID:25723,11
pages
10.12677/BR.2018.74046
Responses of Cleistogenes songorica Blade Ecological Anatomy and N/P Stoichiometry to Stocking Rates
Haijun Chen1,2, E Er-Dun-Hua2, Yumei Shan3,4, Lijun He5, Yong Yang6, Chunli Bai3, Mingjiu Wang2
1Inner Mongolia Institute of Biotechnology, Hohhot Inner Mongolia
2College of Grassland, Resources and Environment, Inner Mongolia Agricultural University, Hohhot Inner Mongolia
3Inner Mongolia Agriculture & Animal Husbandry Academy of Sciences, Hohhot Inner Mongolia
4Inner Mongolia Prataculture Research Center, Chinese Academy of Sciences, Hohhot Inner Mongolia
5Agricultural College, Inner Mongolia Agricultural University, Hohhot Inner Mongolia
6Inner Mongolia Institute of Grassland Survey and Planning, Hohhot Inner Mongolia
Received: Jun. 11th, 2018; accepted: Jun. 25th, 2018; published: Jul. 2nd, 2018

ABSTRACT
Cleistogenes songorica, a perennial herb, is the constructive and dominant species of desert steppe in northern China. With excellent palatability as well as heat/drought/cold tolerance and trampling resistance, this species has significant effects on the restoration and conservation of desert steppe. In order to understand the response and adaption strategy of Cleistogenes songorica to grazing, we carried out field experiment to examine the influence of different stocking rates on its blade anatomy structure and N/P stoichiometry. Leaf samples of Cleistogenes songorica (3rd totally matured and sun-sloped disease-free blade from the top) were collected from a controlled-experimental platform with four stocking densities, which has been lasting for nine years on the desert steppe of Inner Mongolia. Using paraffin method, upper epidermal cuticle thickness, lower epidermis thickness, upper epidermal cell thickness, lower epidermis cell thickness, upper epidermal cell area, lower epidermis cell area, main vein thickness, leaf thickness, measured using high-power microscope. Total N and P contents were measured by Semi-micro kjeldahl determination and the H2SO4-HClO4 fusion method, respectively. All leaf anatomic characters of Cleistogenes songorica, except for upper and lower epidermal cell thickness, showed significant variation among different grazing rates (P < 0.05). However, there was no consistent pattern for the changes of these anatomic characters with the increase of stocking rates. Leaf N and P contents showed limited changes under LG and MG levels. Grazing significantly increased leaf N contents (P < 0.05), with the highest value in HG. Total P contents in HG were significantly lower than the 3 others (P < 0.05). Leaf N:P was highest in HG and lowest in MG. In the control plots, leaf N contents were negatively correlated with upper epidermal cell area (P < 0.05). In the MG plots, total P contents were positively correlated with upper epidermal cell area (P < 0.05) and negatively correlated with main vein thickness (P < 0.05).
Keywords:Desert Steppe, Cleistogenes songorica, Ecological Anatomy Structure, N & P, Stoichiometry, Stocking Rate
无芒隐子草(Cleistogenes songorica)叶片生态解剖及氮磷化学计量特征对载畜率的响应
陈海军1,2,额尔敦花2,单玉梅3,4,何丽君5,杨勇6,白春利3,王明玖2
1内蒙古自治区生物技术研究院,内蒙古 呼和浩特
2内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特
3内蒙古自治区农牧业科学院,内蒙古 呼和浩特
4中国科学院内蒙古草业研究中心,内蒙古 呼和浩特
5内蒙古农业大学农学院,内蒙古 呼和浩特
6内蒙古自治区草原勘察规划院,内蒙古 呼和浩特
收稿日期:2018年6月11日;录用日期:2018年6月25日;发布日期:2018年7月2日

摘 要
无芒隐子草为多年生强旱生密丛禾草,是荒漠草原的建群种和优势种,不仅适口性好,而且具有抗热、抗旱、耐寒和耐践踏等特性,对荒漠草原生态系统的恢复和保育具有重要作用。该文以内蒙古荒漠草原无芒隐子草为研究对象,分析了不同载畜率(不放牧、轻度放牧、中度放牧、重度放牧)对无芒隐子草叶片解剖结构和N、P化学计量特征的影响,以探究无芒隐子草叶片上述性状对环境改变的响应与适应对策。结果表明:不同载畜率水平下,无芒隐子草叶片解剖结构除了上、下表皮细胞厚度之外,其余的指标均产生了显著变化(P < 0.05),但随着载畜率的增加没有明显的一致性变化规律。所有的放牧处理均显著增加了无芒隐子草叶片中的全N含量(P < 0.05)。轻度和中度放牧没有显著影响叶片全P含量,而重度放牧显著降低了叶片的全P含量(P < 0.05)。任何放牧处理对叶片N:P比均没有显著影响。在CK条件下,无芒隐子草叶片的全N含量与上表皮细胞面积之间存在显著的负相关(P < 0.05)。在MG条件下,全P含量与上表皮细胞面积之间存在显著的正相关(P < 0.05),同时与主脉厚度之间存在着极显著负相关(P < 0.01)。因此,在不同载畜率下,无芒隐子草叶片解剖结构及N、P化学计量学特征的变化和二者之间的关系均存在一定的差别。
关键词 :荒漠草原,无芒隐子草,解剖结构,氮和磷,化学计量学,载畜率

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
生态化学计量学(ecological stoichiometry)既是研究生物系统能量平衡和多重化学元素(主要是碳、氮和磷)平衡的科学,也是分析化学元素平衡与生态学过程间相互作用的一种理论,还是研究植物元素分配的重要方法 [1] [2] [3] [4] ,更为研究碳(C)、氮(N)与磷(P)等元素在生态系统过程中的耦合关系提供了一种综合方法 [2] [4] 。由于元素化学功能的耦合性以及元素的不可替代性,植物对N与P的需求和利用存在严格的比例。植物N、P化学计量特征,包括:N、P浓度、N:P及N、P间异速指数等,在不同功能群、生长地区、生长季、器官之间以及环境梯度下存在明显的变化规律 [4] [5] 。
放牧对植被的影响一直是放牧生态学研究的重要领域之一,其研究内容主要集中在放牧对植物个体、种群、群落的影响以及放牧对土壤的影响 [6] 。草原植物对放牧的忍耐和适应,或抵抗和躲避是其生存和发育的策略和方式 [7] 。植物在长期放牧过程中,已经进化出多种适应机制,以保护其与非生物环境和放牧家畜的协调共存,并在群落中与其他物种保持良好的竞争 [8] [9] 。放牧对植物元素含量及化学计量学特征的作用机制已被提出 [4] [10] 。首先,放牧通过植物–土壤层面的反馈作用改变植物元素含量和化学计量特征。放牧增加植物根系分泌物中的化感物质,以刺激土壤微生物活性和周转率,提高土壤养分有效性,促进植物对养分的吸收和利用,从而影响植物体内元素的化学计量关系 [4] [11] 。然而,这些刺激性反馈作用在高放牧率或资源环境相对贫瘠的环境中是难以维持的;其次,放牧通过改变群落的结构与组成,进而影响植物的生长过程中元素含量及其化学计量特征 [4] [10] 。
此外,养分环境是植物赖以生存与发展的基础条件,在各种不同的生境中,聚生着特定的植物种类。植物长期生活在各种特定环境中,获得(或进化)了一些适应环境相对稳定的特性,其中包括形态结构方面的适应特征。同时,随着环境因子的改变,植物在形态结构上也出现某些变化,其中受影响比较大的主要是植物的营养器官 [12] [13] [14] 。有研究发现,长期放牧条件下,放牧率通过改变角质层厚度或表皮细胞的面积大小等影响了植物叶片解剖结构,进而影响植物对养分及化学元素的吸收。不同物种、功能群植物性状对放牧干扰反应程度不同,其结构和生态化学特征均受到放牧的显著影响 [15] 。因此,对不同载畜率水平下的植物叶片生态解剖与N、P化学计量特征的研究将能更好地阐明叶片生理生态对环境变化的响应机理。
无芒隐子草(Cleistogenes songorica)既是禾本科多年生强旱生草本植物,也是短花针茅(Stipa breviflora)荒漠草原的优势种之一,还是我国西部比较重要的牧草,具有很好的饲草价值。由于其具有抗旱、抗寒和耐风沙的特性,已成为一种具有重要生态和遗传价值的植物材料 [16] 。荒漠草原气候干旱,养分匮乏,物种之间的竞争主要是对水分资源和养分资源的竞争。叶片是植物进行光合作用和蒸腾作用的主要器官,其结构特征不仅是生理作用的基础,也是环境变化的基础。叶片解剖结构的改变将影响其生理功效和对环境适应性的发挥,即植物解剖结构特征不仅能够反映所处环境的变化,也能够揭示植物对环境的适应过程和对策。
在此背景下,本文以内蒙古荒漠草原无芒隐子草为研究对象,拟解决两个科学问题:1) 无芒隐子草解剖结构是否对载畜率存在响应变化?2) 无芒隐子草叶片N、P化学计量特征是否随载畜率变化而变化,及其与解剖结构的相关性,由此探究无芒隐子草叶片解剖结构和N、P 化学计量特征等功能性状对载畜率的响应过程及适应对策,旨为荒漠草原退化生态系统的恢复与放牧管理提供基础依据。
2. 材料与方法
2.1. 试验区自然概况
试验地设在内蒙古农牧业科学院四子王旗试验基地内,地理位置为:111˚53'41.7''E,41˚46'43.6''N,平均海拔为1456 m。研究区处于中温带大陆性季风气候区,春季气温变化剧烈,大风日数较多;夏季炎热,降雨集中,日照充裕;秋季秋高气爽,降雨逐渐减少;冬季漫长而寒冷,气候干燥,多寒潮寒流天气,土地封冻。根据四子王旗气象站1961~2010年监测气象数据显示,多年平均气温为3.6℃,年均降水量为311.4 mm,主要集中在6~9月,占全年降水量的70%以上。无霜期约90~105 d。
试验区植被是以短花针茅为建群种的荒漠草原,植物群落类型为短花针茅(Stipa breviflora) + 冷蒿(Artemisia frigida) + 无芒隐子草,植被低矮,平均高度一般为8 cm,且植被较稀疏,盖度为12%~18%。种类较贫乏,植物群落主要由20余种植物组成。主要多年生草本植物为短花针茅,无芒隐子草、阿尔泰狗娃花(Heteropappus altaicus)、银灰旋花(Convolvulus ammannii)、冰草(Agropyron cristatum)、羊草(Leymus chinensis)等;一、二年生草本植物有蓖齿蒿(Artemisia pectinata)、猪毛菜(Salsola collina)等。还有小半灌木冷蒿(Artemisia frigida)和木地肤(Kochia prostrata),矮灌木狭叶锦鸡儿(Caragana stenophylla),灌木小叶锦鸡儿(Caragana microphylla)等,其中无芒隐子草在群落中生物量所占比重为20%以上。土壤为淡栗钙土。
2.2. 研究内容与方法
2.2.1. 试验设计
整个放牧试验开始于2003年,至本试验取样时为止,已具有9年的连续放牧历史。采用完全随机区组设计,每个区组内设3个载畜率水平和1个无牧区作为对照,分别表示为轻度放牧(LG,0.93只/hm2∙半年)、中度放牧(MG,1.82只/hm2∙半年)、重度放牧(HG,2.71只/hm2∙半年)和对照区(CK,不放牧),各处理重复3次,共计12个试验区。各试验区面积基本相等,约为50 hm2。供试羊只为2岁蒙古细毛羯羊。试验期每年持续6个月(5月初至11月末),期间各试验区供试羊只自由采食,夜间赶回畜圈休息,统一管理。
2.2.2. 样品采集
在2011年8月中旬从上述12个放牧试验区取材料。在各试验小区随机选择无芒隐子草株丛,并采集其新鲜叶片。叶片选择标准为:向阳面、完全成熟(从顶部数第3个成熟叶片)、无病虫害。将叶片切成0.5~1.0 cm的小段快速放入FAA固定液(70%酒精:冰乙酸:福尔马林溶液 = 90:5:5)中保存,封存后带回实验室进行相关性状观测。
同时,采集另一份相同标准的无芒隐子草完整叶片置于纸袋中,标记后带回实验室。
2.2.3. 切片制作和叶片解剖结构参数的测定
将固定的叶片段,先用蒸馏水清洗干净,然后用不同浓度(70%,85%,95%和100%)酒精依次脱水、经过二甲苯透明、渗蜡、包埋和修块等实验过程,采用石蜡切片法切成10~12 μm厚的切片,再用番红固绿对染,然后用加拿大树胶封片,充分干燥后,在Olympus光学显微镜下进行拍照,所得照片在Photoshop8.0软件下观测叶的上表皮角质层厚度(Ctue)、下表皮角质层厚度(Ctle)、上表皮细胞厚度(Tuec)、下表皮细胞厚度(Tlec)、上表皮细胞面积(Auec)、下表皮细胞面积(Alec)、主脉厚度(Mvt)和叶片厚度(Lt)等8项指标,每项指标重复5次,取其平均值。
2.2.4. 叶片N和P元素测定
将纸袋中的叶片清洗干净后,置于65℃恒温条件下烘干至恒重后,用球磨仪(Retsch MM400;Retsch, Haan, Germany)进行粉碎。粉碎样品的全N含量采用凯氏定氮法进行测定(Kjeltec 2300 Auto System II, Foss Tecator AB, Hoganas, Sweden),全P含量用钼锑抗比色法进行测定 [17] 。
2.2.5. 数据处理
首先用Excel 2010对所有的原始数据进行初步分析、处理。然后用SAS 9.0软件(SAS Institute,Cary,North Carolina,USA)对数据进行方差分析以及各解剖结构指标之间、化学成分含量与各指标间做相关性分析。
所有作图均在Sigmaplot 11.0 (Systat Software Inc.)软件环境下完成。
3. 结果与分析
3.1. 无芒隐子草叶片的基本解剖结构特征
无芒隐子草叶的横切面为长条形,由表皮、叶肉和维管束构成(图1 CK)。其中,表皮由单层细胞构成,表皮细胞近圆形,排列紧密,大小不一,有些细胞壁向外形成突起。在光学显微镜下,上、下表皮具明显的角质层(Cu)被染成红色(图1 LG)。叶肉包括下表皮(Le)、栅栏组织和海绵组织。维管束鞘(Vbs)有2层细胞,其中外层细胞体积较大,从横切面角度上看,其直径明显大于叶肉厚度,细胞质较浓。泡状细胞(Bc)较发达,其厚度相当于叶肉厚度。
3.2. 载畜率对无芒隐子草叶片解剖结构的影响
不同载畜率水平对无芒隐子草叶片的Ctue、Ctle、Mvt和Lt均产生显著影响(P < 0.05) (图2(a)),但随载畜率的增加没有一致性的变化规律。Ctue表现为CK > LG > HG > MG,在MG显著小于其他3个载畜率水平(P < 0.05),其中CK、LG和HG间无显著差异(P > 0.05)。Ctle表现为MG > LG > CK > HG,

Figure 1. The anatomy structure for foliar transverse section of Cleistogenes songorica under different stocking rates (×400)
图1. 不同载畜率下无芒隐子草叶片横切面解剖结构(×400)

Figure 2. The character index variation of Cleistogenes songorica foliar anatomy structure under the different stocking rates (Thickness including Ctue, Ctle, Tuec, Tlec, Mvt and Lt) (The different lowercase letters identified in the histogram represent significant differences at the 0.05 level. The same as follows)
图2. 不同载畜率下无芒隐子草叶片解剖结构性状的变化(厚度包括:上、下表皮角质层厚度,上、下表皮细胞厚度,主脉厚度和叶片厚度等) (柱状图中标识的不同小写字母表示在0.05水平下差异性显著,以下同)
在HG显著小于其他3个载畜率水平(P < 0.05),CK、LG和MG间均无显著差异(P > 0.05),但这3个水平有着随载畜率的增加而加厚的趋势。可见,MG和HG对无芒隐子草叶片的Ctue和Ctle影响显著,而LG对Ctue和Ctle的影响不显著。Mvt表现为CK > LG > HG > MG,CK和LG显著高于MG和HG (P < 0.05)。Lt表现为CK > LG > HG > MG,这与Ctue在各载畜率水平下变化趋势相一致。另外,Tuec和Tlec二者在CK、LG、MG和HG间均无显著差异(P > 0.05),这表明到目前为止,放牧干扰对Tuec和Tlec二者均无显著的影响。
Auec表现为MG > HG > LG > CK (图2(b)),其中CK和LG之间存在显著差异(P < 0.05),且CK显著小于MG (P < 0.05)。Alec则为MG > LG > CK > HG,LG和MG显著高于HG (P < 0.05),这3个载畜率水平与CK间均无显著差异(P > 0.05)。总之,MG对Auec和Alec的影响最大,而HG条件下的Auec和Alec与CK条件下的Auec和Alec之间没有显著差异(P > 0.05)。综上可知,LG对无芒隐子草叶片解剖结构各指标的影响不明显。
3.3. 载畜率对无芒隐子草叶片N、P含量及其计量比的影响
无芒隐子草叶片的全N、全P含量在不同载畜率水平下的变化模式不相同(图3)。叶片全N含量在CK显著低于LG、MG和HG (P < 0.05),说明动物啃食对叶片全N有显著影响,但是全N含量对不同载畜率水平的反映不明显。全P含量在HG显著低于其他3个载畜率水平(P < 0.05),说明全P含量在重度载畜率下有显著响应。
无芒隐子草叶片氮磷比(N/P)在HG最高,在MG最低。由此可知,MG和HG对全N:全P的影响较大,不利于无芒隐子草叶片全N含量与全P含量的正常运转,从而影响整个植物的充分生长。总之,LG在一定程度上增加了无芒隐子草叶片的全N含量,有利于植物的正常生长。
3.4. 载畜率水平下的无芒隐子草解剖结构各指标与N、P之间的相关关系
不同载畜率水平下的无芒隐子草叶片解剖结构个别指标与全N含量和全P含量显示了较显著的相关关系(表1)。其中,在CK条件下,无芒隐子草叶片全N含量与Auec之间存在显著的负相关关系(P < 0.05),

Table 1. The correlation between index of Cleistogenes songorica foliar anatomy structure and N&P under the different stocking rates
表1. 不同载畜率下无芒隐子草叶片解剖结构性状指标与N、P间的相关性分析
r, correlation coefficient; p, significance level. * and ** indicate that the regression coefficients are significant at the 0.05 and 0.01 levels. r,相关系数;p,显著性水平;*和**分别表示显著性达到0.05和0.01水平。
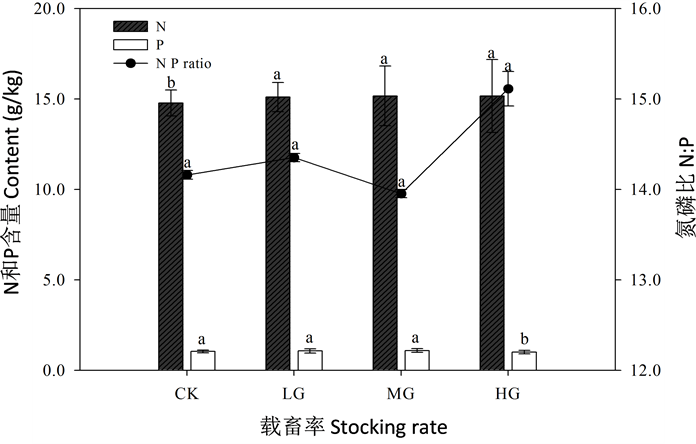
Figure 3. The N & P content and N P ratio of Cleistogenes songorica foliar under the different stocking rates
图3. 不同载畜率下无芒隐子草叶片N、P含量及N:P比变化
全P含量与解剖结构各指标之间不存在显著的相关关系。在MG条件下,无芒隐子草叶片全P含量与Auec呈显著的正相关关系(P < 0.05),同时与Mvt呈负相关关系(P < 0.01)。可见,在MG条件下的无芒隐子草叶片全P含量与解剖结构有相对明显的相关关系。
在LG和HG条件下,无芒隐子草的全N含量及全P含量都与解剖结构各指标之间的相关关系不明显(P > 0.05)。
4. 讨论
4.1. 无芒隐子草叶片解剖结构对载畜率的响应
载畜率的不同影响了无芒隐子草叶片解剖结构。植物的叶片角质层为脂肪类物质,是反映植物抗旱能力的一个重要指标,即能够防止植物的体内水分过分蒸腾。较厚的角质层会降低牧草的适口性,所以普遍认为植物角质层变厚是草原植物普遍的对家畜啃食的一种防御措施 [15] [18] [19] 。从分析结果看出,无芒隐子草叶片Ctue和Ctle在MG显著小于CK、LG和HG (P < 0.05) (图2(a)),CK、LG和HG彼此间均无显著差异(P > 0.05),但3个水平有着随载畜率的增加而加厚的趋势。在放牧条件下,土壤紧实且渗透率低 [15] [20] ,植物为了争取更好的生存环境而提高水分利用率,就需要降低因蒸腾而丧失的水分。无芒隐子草叶片的Tuec和Tlec对载畜率水平的响应不显著(P > 0.05),可能是植物对荒漠草原严酷的生态环境长期适应的结构基础。无芒隐子草叶片的表皮由一层细胞构成,以上结果说明无芒隐子草的叶片表皮细胞层数和厚度可能是较为保守的性状,受环境因子变化的影响比较小,这与李芳兰等相关研究结果基本一致 [21] 。
Auec和Alec均在MG达到最大值。其中Auec在CK和LG之间存在显著差异(P < 0.05),CK显著小于MG (P < 0.05)。Alec在CK、LG和MG条件下,呈现随着载畜率的增加而变大。Ctue和Ctle的表现正好相反,在MG达到最小值,而且显著小于其他3个载畜率水平(P < 0.05)。无芒隐子草是旱生植物,加强了旱生结构的特性 [22] 。过度放牧退化后,草地植物株丛矮小,群落中的枯落层消失,群落中地表裸露度较大,地表蒸发较强 [18] [23] ;土壤近地表层因家畜践踏而紧实,降雨后水分下渗速率降低,且因涵养水源能力丧失而易于形成地表径流,从而使群落可获得的水量大幅度降低,改变了植物生长的小环境,植物表现出旱生或加强旱生结构的特性,如角质层变厚、叶肉细胞密度增大、叶片增厚、叶表皮细胞变大。
Lt是反映植物抗旱性的另一个指标,抗旱性强的植物往往叶片较厚 [15] [18] 。本研究发现(图2(a)),无芒隐子草的Lt和Mvt均在CK条件下达到最大值。Lt在MG显著低于其他3个载畜率水平(P < 0.05),说明中度载畜率使旱生植物无芒隐子草叶片厚度变薄,其影响正常生长。这跟Ctue的表现相同。主脉厚度在CK和LG显著高于MG和HG (P < 0.05)。有研究表明,扁蓿豆、糙隐子草、冷蒿、羊草的叶片厚度及主脉厚度均随着放牧率增大而降低 [15] 。本研究表明,无芒隐子草叶片结构对不同环境因素的适应性比较强。叶作为植物暴露在环境中面积最大的器官,受水分、温度光照、牲畜的采食等环境因子的影响,主要表现为叶的外部形态的变化、叶厚度的变化及内部解剖结构的差异 [18] ,这也在本研究中得到了验证。
4.2. 无芒隐子草叶片N、P含量与N:P对载畜率的响应
植物叶片中N、P分配因物种、生理生长策略以及土壤环境等的不同而异,但大量研究发现植物叶片N、P的分配服从某一化学计量规律 [5] 。围封和放牧中的植物N、P化学计量存在明显差异 [5] [24] 。本研究的结果进一步显示:不同的放牧强度和频率对草地生态系统植物化学计量同样存在显著影响。无芒隐子草叶片的全N含量在CK显著低于在LG、MG和HG (P < 0.05),在HG最高。全P含量在HG显著低于其他3个载畜率水平(P < 0.05),在MG略高于其他3个载畜率水平。说明无芒隐子草叶片全N、全P含量都在LG、MG变化幅度较小。N:P是衡量生物体营养状况和判断植物群落受养分限制情况的重要指标 [25] [26] 。本研究中,无芒隐子草叶片的N:P在HG最高,在MG最低,表明MG和HG对无芒隐子草叶片的全N:全P的影响较大。可知,HG使全N含量变高,从而全P含量变低。相反,MG使全N含量变低,从而全P含量变高。受牲畜类别、食性偏好等影响,放牧对生态系统的干扰程度也明显不同。放牧可影响植物功能和生产、养分的积累,改变土壤养分状况和显著影响植物化学计量特征 [5] [27] 。在自然界中,植物N:P化学计量特征虽然能较好地反映出N、P养分的限制作用,只反映了N、P元素限制作用的相对大小以及相互转化趋势,其价值主要在于指示作用,故而对N:P的诊断意义应该客观对待 [3] [28] 。白永飞等利用样带研究法对欧亚草原进行生态化学计量学的研究,结果表明放牧改变了生态系统功能和C:N:P生态化学计量学特征,其中放牧对植物生态化学计量学的影响,草甸草原植物最为明显,典型草原次之,荒漠草原差异不明显,这与本研究结果相一致 [4] 。陈海军对荒漠草原不同载畜率水平下优势植物生态化学计量学研究表明,长期持续放牧影响了无芒隐子草不同构件的生态化学计量学特征变化,不同构件的特征对对载畜率响应过程不同,无芒隐子草茎和主根均受N限制;叶受N和P共同限制;侧根在CK受P限制,在LG、MG和HG下受N和P共同限制。无芒隐子草种群整体、地上部分和地下根系均受N限制 [10] 。
4.3. 不同载畜率水平下的无芒隐子草叶片解剖结构与N、P化学计量的相关性
植物化学元素含量反映了植物在一定生境条件下从土壤中吸收和蓄积矿质养分的能力。植物的N、P化学计量特征与植物特性之间的关系可以解释植物群落的功能差异及其对环境变化的适应性 [5] [29] [30] ,同时对评定N、P对陆地生态系统初级生产力的限制作用具有重要意义 [5] [31] [32] 。本研究对不同载畜率水平下的无芒隐子草叶片的解剖结构与N、P化学计量特征之间的关系。在CK条件下,无芒隐子草叶片全N含量与Auec之间存在显著的负相关(P < 0.05,r = −0.997) (表1),说明在无放牧条件下,无芒隐子草叶片全N含量的增加导致上表皮细胞面积变小。在MG条件下,全P含量与Auec之间存在显著的正相关(P < 0.05,r = 0.999),同时与主脉厚度之间存在着极显著负相关(P < 0.01,r = −1.000),表明中度放牧条件下的无芒隐子草叶片的全P含量与解剖结构有相对明显的相关性,可以认为随着全P含量的增加Auec有变大的趋势,而全P含量和主脉厚度的关系却相反。在轻度放牧和重度放牧条件下的无芒隐子草叶片解剖结构各指标与全N含量和全P含量之间的相关性不太明显,因为荒漠草原生态系统中影响植物叶片N和P含量的因素是复杂和综合的,不同种群(或群落)的养分限制性大小受众多因素的控制,亟需今后进一步深入研究。
5. 结论
不同载畜率水平下,无芒隐子草叶片解剖结构除了Tuec和Tlec之外其余各指标均产生了显著变化(P < 0.05),但随着载畜率的增加没有一致性的变化规律。
无芒隐子草叶片全N、全P含量均在LG、MG变化幅度较小。放牧显著增加了全N含量(P < 0.05),在HG略高于其他3个载畜率水平。全P含量在HG显著低于其他3个水平,在MG略高于其他3个载畜率水平,载畜率(放牧干扰)对叶片的N:P未产生了显著影响(P > 0.05)。
不同载畜率水平下的无芒隐子草叶片解剖结构与全N含量及全P含量之间的相关关系不明显。在CK条件下的无芒隐子草叶片的全N含量与上表皮细胞面积之间存在显著的负相关关系(P < 0.05)。而在MG条件下的无芒隐子草叶片全P含量与上表皮细胞面积之间存在显著的正相关(P < 0.05),同时与主脉厚度之间存在着极显著负相关(P < 0.01)。
致谢
内蒙古自然科学基金项目(2009MS0410 和 2015MS0384)、内蒙古草甸草原生态保育与可持续利用模式研究课题(20091402)、内蒙古自治区科技计划项目(201602083)、内蒙古农业大学科研专项资金(YZFC2017023)和内蒙古农牧业创新基金项目(2017CXJJM07)资助。
文章引用
陈海军,额尔敦花,单玉梅,何丽君,杨勇,白春利,王明玖. 无芒隐子草(Cleistogenes songorica)叶片生态解剖及氮磷化学计量特征对载畜率的响应
Responses of Cleistogenes songorica Blade Ecological Anatomy and N/P Stoichiometry to Stocking Rates[J]. 植物学研究, 2018, 07(04): 375-385. https://doi.org/10.12677/BR.2018.74046
参考文献
- 1. Elser, J.J., Dobberfuhl, D., MacKay, N.A. and Schampel, J.H. (1996) Organism Size, Life History, and N:P Cycle. Biogeochemistry, 37, 237-252.
- 2. 贺金生, 韩兴国. 生态化学计量学:探索从个体到生态系统的统一化理论[J]. 植物生态学报, 2010, 34(1): 2-6.
- 3. 银晓瑞, 梁存柱, 王立新, 王炜, 刘钟龄, 刘小平. 内蒙古典型草原不同恢复演替阶段植物养分化学计量学[J]. 植物生态学报, 2010, 34(1): 39-47.
- 4. Bai, Y.F., Wu, J.G., Chris, M.C., Pan, Q.M., Zhang, L.X., Chen, S.P., Wang, Q.B. and Han, X.G. (2012) Grazing Alters Ecosystem Functioning and C:N:P Stoichiometry of Grasslands along a Regional Precipitation Gradient. Journal of Applied Ecology, 49, 1204-1215. https://doi.org/10.1111/j.1365-2664.2012.02205.x
- 5. 刘超, 王洋, 王楠, 王根轩. 陆地生态系统植被氮磷化学计量研究进展[J]. 植物生态学报, 2012, 3(11): 1205-1216.
- 6. 闫瑞瑞, 卫智军, 杨静, 周忠义. 短花针茅草原优势种群特征对不同放牧制度的响应[J]. 干旱区资源与环境, 2008, 7(22): 188-191.
- 7. 侯牡丹, 王明玖, 李志强, 杨宁, 陈海军. 放牧强度对短花针茅草原3种主要植物营养繁殖性能的影响[J]. 内蒙古农业大学学报, 2009, 1(30): 71-75.
- 8. Novoplansky, A. (1996) Hierarchy Establishment among Potentially Similar Buds. Plant, Cell and Environment, 19, 781-786. https://doi.org/10.1111/j.1365-3040.1996.tb00415.x
- 9. Gordon, I.J. (2003) Browsing and Grazing Ruminants: Are They Different Beasts? Grassland Ecology and Management, 181, 13-21. https://doi.org/10.1016/S0378-1127(03)00124-5
- 10. 陈海军. 荒漠草原主要植物种群繁殖性状及化学计量特征对载畜率的响应[D]: [博士学位论文]. 呼和浩特: 内蒙古农业大学, 2011.
- 11. Anderson, T.M., Ritchie, M.E., Mayemba, E., Eby, S., Grace, J.B. and McNaughton, S.J. (2007) Forage Nutritive Quality in the Serengeti Ecosystem: The Roles of Fire and Herbivory. The American Naturalist, 170, 343-357. https://doi.org/10.1086/520120
- 12. 郑学平, 张承烈, 陈国仓. 河西走廊芦苇的光合碳同化途径对生境条件的适应[J]. 植物生态学报, 1993, 17(1): 1-8.
- 13. 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较[J]. 生态学报, 2005, 2(25): 304-311.
- 14. 高强. 17种荒漠植物的形态与环境适应性[D]: [硕士学位论文]. 呼和浩特: 内蒙古农业大学, 2011: 1-21.
- 15. 赵雪艳, 汪诗平. 不同放牧率对内蒙古典型草原植物叶片解剖结构的影响[J]. 生态学报, 2009, 6(29): 2913-2916.
- 16. 曲涛. 无芒隐子草(Cleistogenes songorica)的抗旱性研究[D]: [硕士学位论文]. 兰州: 兰州大学, 2008: 1-21.
- 17. Sparks, D.L., Page, A.L., Loeppert, P.A., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T. and Sumner, M.E. (1996) Methods of Soil Analysis Part 3: Chemical Methods. Soil Science Society of America and American Society of Agronomy, Madi-son.
- 18. 于向芝, 贺晓, 张韬, 王炜. 内蒙古退化草原8种植物叶结构对禁牧的响应[J]. 生态学报, 2007, 4(27): 1640-1641.
- 19. Bussotti, F., Bottaci, A. and Bartolesi, A. (1995) Morphoanatomy Alterations in Leaves Collected from Beech Trees in Condition of Natural Water Stress. Environmental and Experimental Botany, 35, 201-213. https://doi.org/10.1016/0098-8472(94)00040-C
- 20. 范春梅, 廖超英, 李培玉, 孙长忠, 许喜明. 放牧强度对林草地土壤物理性状的影响——以黄土高原丘陵沟壑区为例[J]. 中国农业科学, 2006, 39(7): 1501-1506.
- 21. 李芳兰, 包维楷, 吴宁. 岷江上游干旱河谷海拔梯度上四川黄栌(Cotinus szechuanensis)叶片的生态解剖[J]. 应用与环境生物学报, 2007, 13(4): 486-491.
- 22. 李正理, 张新英. 植物解剖学[M]. 北京: 高等教育出版社, 1983: 246-261.
- 23. 王炜, 梁存柱, 刘钟龄, 郝敦元. 羊草+大针茅草原群落退化演替机理的研究[J]. 植物生态学报, 2000, 24(3): 268-274.
- 24. 董晓玉, 傅华, 李旭东, 牛得草, 郭丁. 放牧与围封对黄土高原典型草原植物生物量及其碳氮磷贮量的影响[J]. 草业学报, 2010, 19(2): 175-182.
- 25. 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索[J]. 植物生态学报, 2005, 29(6): 1007-1019.
- 26. 徐冰, 程雨曦, 甘慧洁, 周文嘉, 贺金生. 内蒙古锡林河流域典型草原植物叶片与细根性状在种间及种内水平上的关联[J]. 植物生态学报, 2010, 34(1): 29-38.
- 27. 杨慧敏, 王冬梅. 草-环境系统植物碳氮磷生态化学计量学及其对环境因子的响应研究进展[J]. 草业学报, 2011, 20(2): 244-252.
- 28. Güsewell, S. (2004) N:P Ratios in Terrestrial Plants: Variation and Functional Significance. New Phytologist, 164, 243-266. https://doi.org/10.1111/j.1469-8137.2004.01192.x
- 29. Bedford, B.L., Walbridge, M.R. and Aldous, A. (1999) Patterns in Nutrient Availability and Plant Diversity of Temperate North American Wetlands. Ecology, 80, 2151-2169. https://doi.org/10.1890/0012-9658(1999)080[2151:PINAAP]2.0.CO;2
- 30. Matson, P.A., McDowell, W.H., Townsend, A.R. and Vitousek, P.M. (1999) The Globalization of N Deposition: Ecosystem Consequences in Tropical Environments. Biogeochemistry, 46, 67-83. https://doi.org/10.1007/BF01007574
- 31. Townsend, A.R., Cleveland, C.C., Asner, G.P. and Bustamante, M.M.C. (2007) Controls over Foliar N:P Ratios in Tropical Rain Forests. Ecology, 88, 107-118. https://doi.org/10.1890/0012-9658(2007)88[107:COFNRI]2.0.CO;2
- 32. Venterink, H.O., Wassen, M.J., Verkroost, A.W.M. and De Ruiter, P.C. (2003) Species Richness-Productivity Patterns Differ between N-, P-, and K-Limited Wetlands. Ecology, 84, 2191-2199. https://doi.org/10.1890/01-0639