Botanical Research
Vol.
08
No.
01
(
2019
), Article ID:
28244
,
8
pages
10.12677/BR.2019.81002
Research Progress on Resources and Nutritional Components of Brasenia schreberi
Hong Liu1, Wen Huang1, Lingli Huang1, Kong Yang2, Jing Wang3, Hairong Xiong1, Zhihua Wu1, Rui Qin1*
1Key Laboratory for Protection and Application of Special Plant Germplasm in Wuling Area of Hubei Province, South-Central University for Nationalities, Wuhan Hubei
2Institute of Qinghai-Tibetan Plateau, Southwest Minzu University, Chengdu Sichuan
3Engineering Research Center of CAAS for Dual Protein, Institute of Food and Nutrition Development, Ministry of Agriculture, Beijing

Received: Dec. 8th, 2018; accepted: Dec. 20th, 2018; published: Dec. 27th, 2018

ABSTRACT
Brasenia schreberi J. F. Gmel., is an aquatic plant, classified as “First-Class National Protected Plants” in China. Due to enrichment of polysaccharides and proteins, as well as various amino acids, microelement and flavonoids, it is considered to be an important material for research and utilization. During the past decades, natural community area, population number and genetic diversity of B. schreberi decreased sharply in that human’s intervention caused the loss of natural habitat and deterioration of water quality. Therefore, to provide the evidence of conservation and sustainable development, we review the progress in germplasm resources and nutritional components of B. schreberi.
Keywords:B. schreberi, Resource Distribution, Genetic Diversity, Nutrients Content
莼菜资源及其营养成份研究进展
刘虹1,黄文1,黄凌丽1,杨孔2,王靖3,熊海容1,吴智华1,覃瑞1*
1中南民族大学武陵山区特色资源植物种质保护与利用湖北省重点实验室,湖北 武汉
2西南民族大学青藏高原研究院,四川 成都
3农业部食物与营养发展研究所,中国农业科学院双蛋白工程技术研究中心,北京

收稿日期:2018年12月8日;录用日期:2018年12月20日;发布日期:2018年12月27日

摘 要
莼菜是我国的一类保护的野生植物,因含有特殊生物活性的多糖、丰富的蛋白质及氨基酸、微量元素和黄酮类化合物等,具有较高的研究和利用价值。近二十年来,由于人类破坏导致自然栖息地丧失和水质恶化等因素,莼菜的自然群落面积和居群数量急剧减少,遗传多样性降低。本文对国内外莼菜种质资源和营养成份的研究进展进行了总结,为莼菜种质资源的保护和综合开发利用提供科学依据。
关键词 :莼菜,资源分布,遗传多样性,营养成份

Copyright © 2019 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
莼菜(Brasenia schreberi J. F. Gmel.),又名水葵、马蹄菜、湖菜等,为睡莲科的一种多年生宿根水生植物,生长于池塘、湖泊和沼泽中。莼菜在亚洲、澳大利亚、非洲、印度、北美洲和南美洲的温带和热带地区具有广泛但零星的地理分布 [1] [2] [3] [4] [5]。在中国主要分布于浙江、江苏和湖北等长江中下游地区 [6] [7] [8] ,近年来调查发现云南、四川、湖南和福建等地也均有分布,由于人为破坏、品种退化及病虫害等因素影响,莼菜原生地区日益减少 [9]。
莼菜以嫩茎和嫩叶供食用,有较高的营养和药用价值。莼菜含有丰富的胶质蛋白、多糖、脂肪、多种维生素、微量元素和黄酮类化合物。吴永尧等人 [12] [13] 对采自恩施的莼菜品种进行营养成份分析发现,莼菜中总蛋白含量占干重质量的34.15%,是烘干莼菜中含量最高的物质,其蛋白质含量远远高于其他水生蔬菜 [13]。此外,莼菜性味甘凉、清热、利尿、消肿、解毒,可治疗热痢、黄疸、痈肿、疗疮等症,也具有调节免疫功能,抗肿瘤,降血糖血脂等药用价值 [10] [11]。本文对国内外对莼菜种质资源和营养价值成份研究进行了归纳总结。
2. 种质资源分析
2.1. 资源分布
莼菜在浅水湖泊、池塘内呈间断分布,其生境隔离现象十分突出。近年来,由于人类活动导致的野生生境破坏及过度的经济开发,莼菜的群落面积与种群数量急剧减少。我国湖北利川、重庆石柱、四川雷波和杭州西湖四大主产区也未见到野生种群分布 [14] [15] [16]。
为了进行莼菜原产地野生种质资源的保护,中南民族大学近年对我国野生莼菜资源进行了大范围的调查,发现除四大人工种植区外,在云南、湖南、江西和浙江等地还存在广泛零星的野生种群分布(资源分布点见图1)。在全国第四次中药资源普查中,郑丽香等人 [17] 在福建柘荣县东源乡发现莼菜野生种群分布,文献报道福建周宁县和屏南县也有野生种群分布 [18] [19]。Li等人也在广东南雄和台湾采集到莼菜样品 [20]。

Figure 1. Location of B. schreberi in China
图1. 莼菜在国内的分布区域
2.2. 遗传多样性分析
莼菜目前被列为中国极度濒危的物种,属于国家一类重点保护野生植物 [21]。因此,有效的保护管理来保护野生莼菜种群势在必行。珍稀植物物种的遗传多样性和种群结构通常对于保护和管理策略的制定至关重要,它为人口学、繁殖和生态学等方面提供了重要的见解 [22]。保护遗传变异也是生物学家保护濒危物种的主要目标,因为保持其在不断变化的环境中生存的进化潜力也至关重要 [28] [29]。
已有研究采用各种分子标记来评估莼菜的遗传多样性,包括简单序列重复标记 [23] [30] ,随机扩增多态性DNA、扩增片段长度多态性 [24] ,核核糖体间隔区和叶绿体DNA序列 [25]。尽管莼菜种群的数量日益减少,但这些研究揭示了显着的遗传多样性,突出了影响遗传多样性的负面因素,并提出了潜在的保护措施。张光富等人 [23] 调查了浙江和苏州两省半天然种群的种群多样性,Kim等人 [25] 则侧重韩国的自然种群。董翔等人 [26] 研究结果表明ISSR标记用于鉴别华中利川和华东苏杭莼菜品种种质资源是有效的,同时揭示了地理位置的隔离及气候地带的差异化与它们在分子水平上的分化是一致的,采用多种分子标记联合构建不同莼菜品种的分子身份证数据库,并与传统性状特征相结合,将有利于更准确地鉴定莼菜品种资源。马梦雪等人 [27] 使用α-多样性指数分析莼菜种群发现人工定植管理和频繁采摘可能是导致莼菜群落的物种多样性降低的重要原因。Li等人 [20] 使用简单序列重复(SSR)标记对21个莼菜种群的多样性和遗传结构进行了分析,发现与其他地方性水生植物相比,莼菜表现出较低的遗传多样性和显著的遗传分化,NJ树状图(图2)表明了研究群体之间的遗传关系,分为两个主要组,每组有两个亚组。此外,这些莼菜分布位置与人口分布处于同一地理区域,这些发现揭示了中国西南部、中部和东南部的人群之间的遗传分化。
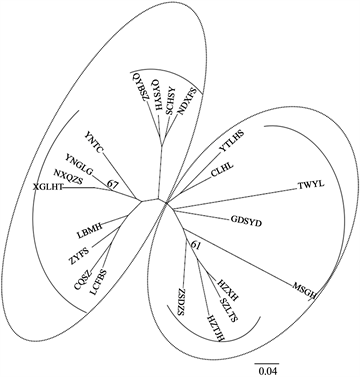
Figure 2. NJ tree based on genetic distances (Nei, 1987) of 21 studied populations of B. schreberi [20]
图2. 基于21个莼菜种群的遗传距离(Nei, 1987)的NJ树 [20]
3. 营养成份
3.1. 蛋白质及氨基酸
莼菜含有丰富的植物蛋白和多种氨基酸。余昌均 [33] 研究发现福宝山莼菜蛋白质鲜重更是高达0.76%。吴永尧等人 [12] 利用凯氏定氮法测得利川莼菜中总蛋白干重含量高达34.15%。周毅峰 [34] 将莼菜嫩叶水溶性蛋白分离发现是由两个富锌蛋白质组分组成,且均有较高的甘氨酸和硒代半胧氨酸,两个富锌蛋白质组分均含有能与锌元素结合的组氨酸和硒代半肤氨酸。食物蛋白质是机体能量的来源之一,而机体内必需的营养素是蛋白质中的氨基酸。吴永尧 [12] 利用氨基酸自动分析仪测得莼菜中18种氨基酸的干重含量,其中必需氨基酸8种,其中的组氨酸和精氨酸是儿童身体需要的特有氨基酸种类。缬氨酸能与异亮氨酸和亮氨酸一起作用促进身体正常生长,修复组织,调节血糖,并提供需要的能量。周德荣等人 [44] 也测得太湖莼菜中胶质、初生卷叶和嫩梢的氨基酸含量。张朝阳等人 [45] 研究发现与露地栽培模式相比,设施栽培能显著提高太湖莼菜的可溶性糖含量、抗氧化性及除精氨酸外的其余15种氨基酸含量。
3.2. 多糖
莼菜鲜样含水量高达97.5%,总糖含量为0.86%,总糖是莼菜多糖,存在于莼菜茎叶上的透明粘液质中。莼菜多糖主要存在于莼菜表面含有的嫩滑胶质中,这也是莼菜的主要特征成分,该多糖主要是碱溶性的酸性多糖和热水溶的水性多糖 [35] [44]。Kakuta等人 [32] 采用Smith降解、PC、电泳、GC、GC-MS、HPLC和IR等方法推断出莼菜多糖的中心链和外围侧链结构,并断定构成莼菜多糖的单糖(质量分数)包括:D-半乳糖(32%~40%),D-甘露糖(10%~14%),L-岩藻糖(13%~16%),L-鼠李糖(6%~9%),D-木糖(2%~7%),L-阿拉伯糖(2%~3%),D-葡萄糖醛酸(12%~29%)和葡萄糖(痕迹量) 8种糖基。该研究工作不仅证实莼菜多糖含有8种糖基,而且对其多糖链的总体结构,糖基间连接方式有了较为清晰的了解,为生物活性研究提供了一定的理论基础。周毅峰 [34] 研究发现莼菜体外胶质并不完全是碱溶性的酸性多糖,其组成成分是复杂的,就多糖而言,还含有热水溶的中性多糖和酸溶性的碱性多糖,其所占比例也不是很小。有研究发现莼菜多糖的校正含有量为88.38%,完全酸水解后,含甘露糖、鼠李糖、葡萄糖醛酸、葡萄糖、半乳糖、木糖和岩藻糖(0.318:0.358:0.317:0.112:1.823:0.725:1.329);莼菜多糖部分酸水解后,含鼠李糖、葡萄糖醛酸、葡萄糖、半乳糖、木糖和岩藻糖(0.154:0.235:0.074:1.662:0.479:1.095)。表明是一种酸性多糖,其主链以甘露糖为主要单糖,支链以半乳糖为主要单糖 [35] [43]。
3.3. 微量元素
中国农科院原子能利用研究所(1992年)对鄂产莼菜样品测出了27种微量元素(Mn, Mg, Cu, Cl, Al, Ca, V, Ge, Cr, La, Ba, Br, As, Sb, Cs, Sc, Rb, Fe, Zn, Co, Na, K, Th, Lu, Se, Hf, Sr),其中硒为0.23 μg/g,铜为56.5 μg/g,锌为153 μg/g~288 μg/g,铁为393 μg/g,钙为0.745% [33]。吴永尧等人 [12] 在利川莼菜的研究中发现,莼菜对环境中的锌有较强的富集作用,其主根、须根和茎都含较高锌,均在0.50 mg/g以上,嫩叶含锌高达1.29 mg/g,有机态锌占总锌的69.77%,蛋白质中锌高达2.92 mg/g,说明纯菜具有很强的将无机锌转化为有机锌的能力,可见莼菜对锌的富集能力远远超过其他植物,是一种强集锌植物 [12] [36]。为进一步证实莼菜对微量元素锌的富集作用,吴永尧等人 [37] 在其它粮食作物以及土壤和水的锌含量进行了分析,发现莼菜对锌的含量明显高于水和土壤中的锌含量,而其它粮食作物则均低于水和土壤中的锌含量,进一步证明了莼菜是一种强集锌的野菜,是一种纯天然的补锌食品。
3.4. 黄酮类化合物
Legault等人 [31] 2011年从莼菜提取物中分离出来两种主要的酚类化合物槲皮素-7-O-β-D-葡萄糖苷和没食子酸,Shimoda等人 [39] 2014年从莼菜中分离出多种黄酮类化合物(图3),其中有一种新型的黄酮醇糖苷Junsainoside A,研究表明,莼菜中的黄酮类化合物可以抑制皮下脂肪细胞的脂质积累。没食子酸乙酯和槲皮素的抑制影响TNF-α诱导的纤维母细胞的损害。此外,还发现阿魏酸、槲皮素和山奈酚可以通过脂联素增强HAS2和HAPB2的表达。

Figure 3. Flavonoids isolated from B. schreberi [38]
图3. 莼菜中分离的黄酮类化合物 [38]
4. 结语
本文对莼菜的资源分布、遗传多样性和营养成份进行了综述,为莼菜种质资源的保护和综合开发利用提供了科学依据。莼菜的栖息地受到人类活动的破坏,包括农业中除草剂的广泛使用和资源过度开发都是造成莼菜濒危的因素 [40] [41] [42]。龚汉雨等人 [15] 的研究表明水土中重金属离子对莼菜生长和产量并未产生实质影响,但重金属离子元素种类和环境条件都会对莼菜生长表现出加性、协同和拮抗效应,上述观点还需要进行深入研究。Li等 [20] 研究发现,与一些濒危物种相比,莼菜遗传多样性较低,种群内(58.68%)的多样性高于种群间(41.32%),群体间地理距离与遗传距离无显著相关性,可能是生境破碎化产生的基因流动障碍所致。
莼菜中的多糖及黄酮类化合物的种类之多在植物中少见,其中构成多糖的糖基非常丰富,多糖结构也尚未完全弄清 [33] ,有待学者进一步研究其复杂的多糖结构与药理之间的关系;Shimoda等人 [38] 在莼菜中发现一种新型的黄酮类化合物Junsainoside A,对其结构和功能需要进一步挖掘利用。莼菜吸收利用环境中锌的能力远远超过其它植物,是一种强集锌植物,是天然富锌食物和食品生产中良好的全天然锌强化剂 [36]。周毅峰 [34] 研究发现莼菜富锌蛋白质中含有硒代半胧氨酸,但是含有硒代半肤氨酸的原因尚待研究;将富锌蛋白所结合的其他大分子分离开,研究富锌蛋白中氨基酸的确定组成,阐明锌结合机制,对未来开发利用莼菜中的富锌蛋白具有重要意义。
基金项目
国家科学技术部科技基础性工作专项重点项目资助(2014FY110100),湖北省科技条件平台建设专项(2017BEC014),湖北省重点实验室后补助专项(2018BFC360)。
文章引用
刘 虹,黄 文,黄凌丽,杨 孔,王 靖,熊海容,吴智华,覃 瑞. 莼菜资源及其营养成份研究进展
Research Progress on Resources and Nutritional Components of Brasenia schreberi[J]. 植物学研究, 2019, 08(01): 7-14. https://doi.org/10.12677/BR.2019.81002
参考文献
- 1. Lloyd, P.J. and Kershaw, A.P. (1997) Late Quaternary Vegetation and Early Holocene Quantitative Climate Estimates from Morwell Swamp, Latrobe Valley, Southeastern Australia. Australian Journal of Botany, 45, 549-563. https://doi.org/10.1071/BT96034
- 2. Miura, T., Takeuchi, K. and Yamano, N. (1979) Fast Neutron Transport through Laminated Iron-Water Shield. Journal of Nuclear Science & Technology, 16, 563-573. https://doi.org/10.1080/18811248.1979.9730947
- 3. Taylor, M.L. and Osborn, J.M. (2006) Pollen Ontogeny in Brasenia (Cabombaceae, Nymphaeales). American Journal of Botany, 93, 344-356. https://doi.org/10.3732/ajb.93.3.344
- 4. Lot, A., Ramos, F. and García, P.R. (2002) Brasenia schreberi JF Gmel. (Cabombaceae), un nuevo registro para Chihuahua, México. Boletín de la Sociedad Botánica de México, 70, 87-88.
- 5. Dkhar, J., Kumaria, S. and Tandon, P. (2011) Molecular Adaptation of the Chloroplast matK Gene in Nymphaea tetragona, a Critically Rare and Endangered Plant of India. Plant Genetic Resources, 9, 193-196. https://doi.org/10.1017/S1479262111000396
- 6. 董元火, 高威, 雷刚, 等. 江苏省苏州市太湖地区莼菜群落的α-多样性特征[J]. 江苏农业科学, 2013, 41(4): 342-343.
- 7. 刘朝贵, 李小孟, 须建, 等. 三大产区莼菜遗传多样性及亲缘关系的RAPD分析[J]. 西南大学学报(自然科学版), 2012, 34(10): 54-59.
- 8. 吴洪梅, 吕泽芳, 张昭, 等. 四大莼菜产区商品成分种类和质量分数测定及比较研究[J]. 西南大学学报(自然科学版), 2017, 39(5): 76-82.
- 9. 郑阳霞, 肖龙仲, 屈正波, 等. 雷波县莼菜产业发展现状与前景展望[J]. 长江蔬菜, 2018(6): 82-84.
- 10. 巩江, 倪士峰, 司衍乐, 等. 莼菜药学研究概述[J]. 安徽农业科学, 2009, 37(10): 4489-4490.
- 11. 李燕, 柯剑鸿, 焦大春, 等. 莼菜的营养价值及其应用研究进展[J]. 长江蔬菜, 2018(18): 36-39.
- 12. 吴永尧, 陈建英. 莼菜营养成份的初步研究[J]. 资源开发与市场, 2000, 16(6): 342-342.
- 13. 彭静, 柯卫东, 叶元英, 等. 317份水生蔬菜种质资源品质性状分析[C]//武汉市蔬菜科学研究所. 全国蔬菜遗传育种学术讨论会论文集, 成都: 中国园艺学会, 2002: 21-28.
- 14. 杨红. 凉山雷波马湖莼菜资源的保护与开发利用[J]. 农学学报, 2011, 1(7): 17-20.
- 15. 龚汉雨, 宋雪扬, 邓楚洪, 等. 重金属离子含量对利川莼菜生长的影响[J]. 生物资源, 2018, 40(1): 83-91.
- 16. 李佳楠, 董元火. 杭州西湖莼菜种质资源保护区的莼菜群落特征[J]. 安徽农业科学, 2010, 38(27): 14905-14906.
- 17. 郑丽香, 缪夏添, 安昌, 等. 福建省种子植物新资料[J]. 福建林业科技, 2017, 44(2): 109-112.
- 18. 何国生. 福建2新记录属及6新记录种[J]. 热带亚热带植物学报, 2009, 17(6): 600-601.
- 19. 陈勇, 吴发旺. 福建省被子植物分布新记录[J]. 亚热带植物科学, 2011, 40(1): 65-66.
- 20. Li, Z.Z., Gichira, A.W., Wang, Q.F., et al. (2018) Genetic Diversity and Population Structure of the Endangered Basal Angiosperm Brasenia schreberi (Cabombaceae) in China. PeerJ, 6, e5296. https://doi.org/10.7717/peerj.5296
- 21. 于永福. 中国野生植物保护工作的里程碑——《国家重点保护野生植物名录(第一批)》出台[J]. 植物杂志, 1999(5): 4.
- 22. Zaya, D.N., Molano-Flores, B., Feist, M.A., et al. (2017) Assessing Genetic Diversity for the USA Endemic Carnivorous Plant Pinguicula ionantha RK Godfrey (Lentibulariaceae). Conservation Genetics, 18, 171-180. https://doi.org/10.1007/s10592-016-0891-9
- 23. 张光富, 高邦权. 江浙莼菜遗传多样性和遗传结构的ISSR分析[J]. 湖泊科学, 2008, 20(5): 662-668.
- 24. Kim, C., Na, H.R. and Choi, H.K. (2008) Conservation Genetics of Endangered Brasenia schreberi Based on RAPD and AFLP Markers. Journal of Plant Biology, 51, 260-268. https://doi.org/10.1007/BF03036125
- 25. Kim, C., Jung, J., Na, H.R., et al. (2012) Population Genetic Structure of the Endangered Brasenia schreberi in South Korea Based on Nuclear Ribosomal Spacer and Chloroplast DNA Sequences. Journal of Plant Biology, 55, 81-91. https://doi.org/10.1007/s12374-011-9193-4
- 26. 董翔, 马梦雪, 刘虹, 等. 基于ISSR图谱构建华中利川和华东苏杭莼菜品种的分子身份证[J]. 华中农业大学学报, 2018, 37(4): 50-57.
- 27. 马梦雪, 董翔, 黄文, 等. 基于α-多样性指数的西湖莼菜群落物种多样性分析[J]. 湖北农业科学, 2018, 57(17): 66-69.
- 28. Shao, J.W., Wang, J., Xu, Y.N., et al. (2015) Genetic Diversity and Gene Flow within and between Two Different Habitats of Primula merrilliana (Primulaceae), an Endangered Distylous Forest Herb in Eastern China. Botanical Journal of the Linnean Society, 179, 172-189. https://doi.org/10.1111/boj.12305
- 29. Hu, Y., Wang, L., Xie, X., et al. (2010) Genetic Diversity of Wild Populations of Rheum tanguticum Endemic to China as Revealed by ISSR Analysis. Biochemical Systematics and Ecology, 38, 264-274. https://doi.org/10.1016/j.bse.2010.01.006
- 30. Liu, H., Long, Z.C., Li, L.N., et al. (2016) Development and Characterization of Microsatellite Loci in Brasenia schreberi (Cabombaceae) Based on the Next-Generation Sequencing. Genetics and Molecular Research: GMR, 15.
- 31. Legault, J., Perron, T., Mshvildadze, V., et al. (2011) Antioxidant and Anti-Inflammatory Activities of Quercetin 7-O-β-D-Glucopyranoside from the Leaves of Brasenia schreberi. Journal of Medicinal Food, 14, 1127-1134. https://doi.org/10.1089/jmf.2010.0198
- 32. Kakuta, M. and Misaki, A. (1979) Polysaccharide of “Junsai (Brasenia schreberi J. F. Gmel)” Mucilage: Constitution and Linkage Analysis. Journal of the Agricultural Chemical Society of Japan, 43, 13.
- 33. 刘永琼, 余世鑫, 桂昭明, 等. 莼菜的研究进展[J]. 武汉化工学院学报, 1997, 19(2): 15-18.
- 34. 周毅峰. 莼菜胶质及含锌生物大分子的初步研究[D]: [硕士学位论文]. 长沙: 湖南农业大学, 2005.
- 35. 崔杰, 屠银芳, 刘婧, 等. 莼菜多糖的分离、纯化及结构初步研究[J]. 中成药, 2018, 40(4): 990-993.
- 36. 吴永尧, 周毅峰, 周大寨. 莼菜中锌分布研究[J]. 湖北民族学院学报(自科版), 2001, 19(4):1-3.
- 37. 吴永尧, 彭振坤, 周大寨, 等. 莼菜富集锌的能力及其锌的分布探讨[J]. 食品科学, 2003, 24(2): 125-127.
- 38. Shimoda, H., Nakamura, S., Hitoe, S., et al. (2014) Anti-Adipogenic Polyphenols of Water Shield Suppress TNF-à ±-Induced Cell Damage and Enhance Expression of HAS2 and HAPB2 in Adiponectin-Treated Dermal Fibroblasts. Natural Products Chemistry & Research, 2, 146.
- 39. 余昌均. 福宝山莼菜的生物学特性和营养成份[J]. 资源开发与市场, 1995, 11(6): 262-263.
- 40. 董元火, 雷刚, 刘红艳. 利川绝迹莼菜种群原因分析[J]. 安徽农业科学, 2010, 38(11): 6047-6048.
- 41. 董翔, 刘虹, 马梦雪, 等. 利川莼菜种质资源原生境保护区的莼菜群落特征[J]. 生物资源, 2018, 40(3): 252-256.
- 42. 刘美玉, 习向银, 罗丽娟, 等. 莼菜资源利用研究综述及展望[J]. 长江蔬菜, 2011 (10): 7-10.
- 43. 王倩. 莼菜多糖的提取及其功能特性的研究[D]: [硕士学位论文]. 无锡: 江南大学, 2009.
- 44. 周德荣, 袁卫明. 太湖莼菜营养成份, 药用价值及烹调技术[J]. 上海农业科技, 1995(5): 47-48.
- 45. 张朝阳, 严吴炜, 李浩宇, 等. 不同栽培方式对太湖莼菜产量及品质的影响[J]. 现代农业科技, 2017(11): 73-74.
NOTES
*通讯作者。