Dynamical Systems and Control
Vol.07 No.03(2018), Article ID:25491,8
pages
10.12677/DSC.2018.73017
Mixed Bursting of Coupled Neuron in the Pre-Bötzinger Complex
Zijian Wang*, Fen Ma, Tongtong Liang
School of Science, North China University of Technology, Beijing

Received: May 2nd, 2018; accepted: Jun. 13th, 2018; published: Jun. 20th, 2018

ABSTRACT
The pre-Bötzinger complex is a kernel part in brainstem region that generates respiratory rhythm, and it is the center in control of respiratory pattern in mammals. In this paper, based on the Butera model, two-coupled neuron networks including the dynamics of calcium ion in the Pre-Bötzinger complex are studied. The generation mechanism and synchronous discharge of mixed bursting are studied. The results show that periodic fluctuation of calcium ions has a great influence on the generation of the mixed bursting, so as the relative position of bifurcation curves.
Keywords:Respiratory System, Pre-Bötzinger Complex, Mixed Bursting, Bifurcation
耦合Pre-Bötzinger复合体神经元中混合簇放电模式的研究
王子剑*,马芬,梁桐桐
北方工业大学理学院,北京
收稿日期:2018年5月2日;录用日期:2018年6月13日;发布日期:2018年6月20日

摘 要
Pre-Bötzinger复合体是新生哺乳动物呼吸节律起源的关键部位,是呼吸节律产生的中枢。本文以pre-Bötzinger复合体中两个耦合的神经元为研究对象,并考虑钙离子动力学的耦合神经元模型。利用快慢动力学和分岔分析,研究混合簇同步放电模式及其产生机制。结果表明钙离子的周期性波动对混合簇放电模式的产生有极大的影响,钙离子的波动及分岔曲线相对位置的改变是混合簇放电产生的主要原因。
关键词 :呼吸系统,Pre-Bötzinger复合体,混合簇放电,分岔

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
众所周知,呼吸运动是一种重要的生理活动,包括人在内的所有哺乳动物都要依靠呼吸运动来维持生命。而呼吸运动又是一种节律性运动,这种节律性活动及其产生机制已逐渐被人们所关注。随着科技的进步和神经生理学及相关领域研究方法的改进,人们对于呼吸中枢的认识逐步深入 [1] 。呼吸行为依赖于大型的神经元网络,该网络分布于大脑区域的神经系统中,如大脑皮层、小脑、杏仁核、脑桥、髓质和脊髓 [2] [3] 。实验表明,在这个大型呼吸网络中,其中的一个区域是产生和调节呼吸节律必不可少的部分,这个区域称为pre-Bötzinger复合体。该区域被认为是新生哺乳动物呼吸节律起源的关键部位,是呼吸节律产生的中枢 [4] 。目前,研究者大多认为呼吸运动之所以能自主地有节律地进行,是因为pre-Bötzinger复合体中的一些细胞群,其中包括一些吸气神经元,它们可自主地按一定的频率发出冲动,即神经元放电活动。这些神经元集群能够与其它呼吸组织互相联系,共同调节和控制呼吸运动。
Pre-Bötzinger复合体中神经元的放电模式可以分为峰放电和簇放电两种基本模式。其中,普遍认为簇放电比峰放电携带更多的神经信息。因此神经元的簇同步放电是目前神经动力学研究的热点之一。Ivanchenko等给出簇放电映射神经元的混沌相位同步并发现相位同步并非只与耦合强度相关 [5] [6] 。Ryland的研究结果表明钠离子及钙离子浓度对簇放电有着重要的影响 [7] 。Rubin等人研究了单个神经元在钙离子影响下的混合簇放电模式及其鲁棒性 [8] 。虽然混合簇放电已经有了一些结果,但是耦合神经元的混合簇放电模式产生机制及其受到的钙的影响仍值得我们进一步研究。
本文主要研究加钙耦合Pre-Bötzinger复合体神经元中钠离子浓度变化条件下混合簇放电模式的产生机制。给出模型后我们首先研究钙离子、钠离子浓度对复杂模式簇放电产生的影响,随后通过快慢分析、分岔分析等方法,发现钙离子的波动导致平衡点位置改变从而产生复杂模式簇放电。最后,给出本文的结论。
2. 模型
Pre-Bötzinger复合体中簇放电的产生是依靠持续钠电流、非特异性阳离子电流以及钙离子电流的,其中钙离子电流是由来自树突的细胞内钙离子激活的,它可能受突触输入和细胞内钙储存的影响,由IP3受体调控 [9] 。基于实验结果,Toporikova和Butera于2011 [10] 年建立了一种基于两个隔间电导的pre-Bötzinger complex神经元模型,该模型的动力学模型如下:
(1a)
(1b)
(1c)
(1d)
(1e)
(1f)
其中,钙离子的特性由方程(1e)和(1f)描述。 , 表示电压,n、m是电压依赖性的门控变量, 是细胞内的钙离子浓度,l表示未被灭活的IP3通道的部分,该通道会影响由 和 表示的胞浆和内质网(ER)之间钙的通量。 、 、 、 、 分别代表泄露电流,钾离子,钠离子,持续钠离子和钙离子的电导。模型(1)中的部分变量及参数具体描述或取值范围见附录。
模型(1)中,相对于其他变量,变量 和l变化过程都极为缓慢,h的变化率要次之,所以上述模型可以看作具有不同快慢时间尺度的动力学模型。方程(1e)、(1f)为钙离子相关的树突细胞子系统,可以近似地看作常数。(1c)是关于变量hi的等式,为慢子系统,其余方程(1a)、(1b)和(1d)为快子系统。由于h1与h2近乎完全相同,所以下文研究中,可以将h1和h2看作一个慢变量,即可令 作为一个变量。
3. 耦合神经元簇同步放电模式
耦合Pre-Bötzinger复合体神经元,在相同的参数下,对不同的初始条件具有一定的敏感性,从而表现出两种不同模式的簇放电。在初始条件不同时为反向簇放电模式,而在初始条件相同时表现为同向簇放电模式,膜电位V1与V2随时间t产生的变化完全一致。当多种类型的簇交替进行时,我们称其为混合
簇放电。本文仅研究同相混合簇放电模式,即 且 ,所以在下文中只提及V1和 。
3.1. 混合簇放电模式
图1中,红色曲线为钙离子浓度 随t的变化趋势,黑色曲线是膜电位V1随t的变化趋势,钙离
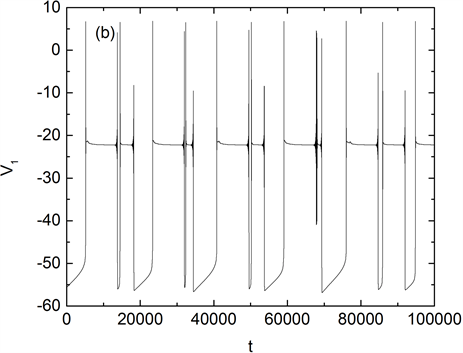
子与电压都呈现周期性的变化。而膜电位V1随时间的变化是由两种不同类型的簇交替产生的即形成混合簇放电模式。此时,其它离子电导的参数值分别为 , , , 。随着钠离子电导gNa的升高,当 时,混合簇的形态逐渐发生变化。两种类型簇中间的静息态逐渐消失,成为单一类型簇,如图2(a)所示。当gNa持续升高至 时,混合簇重新出现,如图2(b)所示。
3.2. 混合簇放电模式的分岔分析
钙离子呈周期性波动变化,如图3(a)所示。将其分为两部分,一部分称作平稳状态,如图3(a)所示的部分I,此时钙离子浓度约为0.034 nS;另一部分称作波动状态,如图3(a)所示的部分II,此时钙离子浓度呈振荡状态。混合簇的出现与钙离子周期性波动关系密切,当钙离子浓度呈平稳状态时混合簇表现为第一类簇,如图3(b)中的簇I;而当钙离子浓度呈波动状态时,混合簇表现为第二类簇。如图3(b)中的簇II。
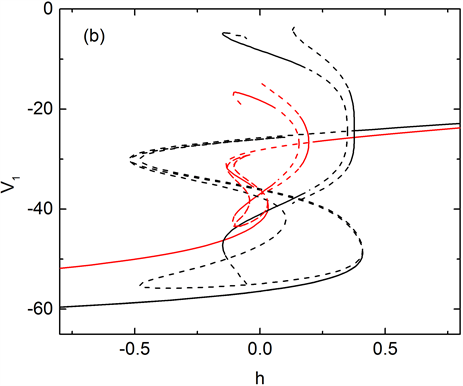
快子系统(1a)、(1b)和(1d)对慢变参数h的分岔如图4(a)所示。平衡点的分岔曲线为S形曲线,S形曲线的下支和中支分别由稳定结点和鞍点组成,稳定结点和不稳定鞍点在鞍结分岔点F1处相遇后消失。S形曲线的上支由焦点组成,随着慢变参数h的增加,不稳定焦点在Hopf分岔点(supH1和supH2)处变为稳定焦点,同时,supH1和supH2点处极限环产生,两组极限环分别在同宿轨分岔(HC)和极限环的鞍结分岔点(LPC)处消失,如图4(a)所示。在钙离子浓度处于平稳阶段,簇I出现,快子系统轨线由静息态经鞍结分岔(F)跃迁至放电态并围绕稳定焦点振荡,且振荡幅度越来越小,直到轨线遇到超临界Hopf分岔点supH2,
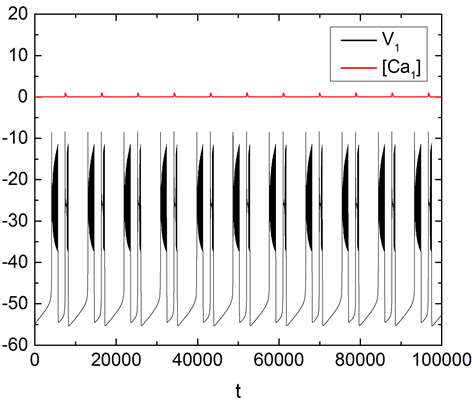
Figure 1. Changes of mixed-mode bursting and concentration of calcium ions with time t when the parameter gNa = 5 nS. The black and red curves represent the change of membrane potential and t of concentration of calcium ion with time t respectively. See Appendix for other parameters
图1. 当参数gNa = 5 nS时耦合神经元混合簇放电模式及钙离子的周期性变化。其中,黑色曲线表示膜电位随时间的变化曲线,红色曲线表示钙离子浓度随时间的变化曲线。其他参数见附录
 (a)
(a)  (b)
(b)
Figure 2. Curve of membrane potential with time t when (a) gNa = 10 nS, which is a single mode bursting. (b) gNa = 28 nS, which is a mixed-mode bursting
图2. (a) gNa = 10 nS时膜电位V1随时间的变化曲线,为单一模式簇放电。(b) gNa = 28 nS膜电位V1随时间的变化曲线,为混合簇放电模式
 (a)
(a)
 (b)
(b)
Figure 3. gNa = 5 ns, (a) Changes of concentration of calcium ions with time t. Here we mark the nearly steady state as partial I, and the bump as partial II. (b) Curve of membrane potential V1 with time t, and the curve of concentration of calcium ion with time t is also superimposed on the graph. Correspondingly, the first and the second bursting in the mix mode bursting are marked as bursting I and bursting II respectively
图3. gNa = 5 ns时(a) 钙离子[Ca1]随时间的变化曲线,钙离子浓度随时间t呈周期性变化,其中标记平部分为部分I,波峰为部分II。(b) 膜电位V1随时间的变化曲线,其中钙离子浓度[Ca1]随时间的变化曲线也叠加在图中。此时,为混合簇放电模式,记这两种簇分别为簇I和簇II
 (a)
(a) 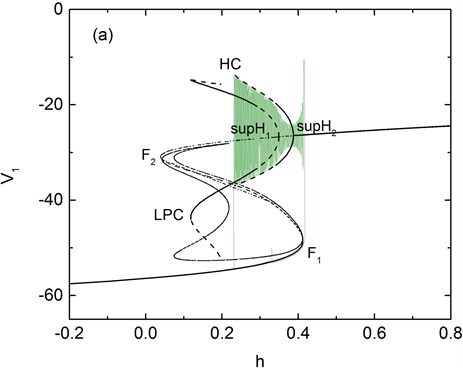 (b)
(b)
Figure 4. (a) Bifurcation diagram of the fast subsystem, the trajectory of the system (green) is also superposed on it. The bursting type is “fold/homoclinic” bursting with gNa = 5 nS. Where supH1 and supH2 are Hopf bifurcation points, F1, F2, HC and LPC denote saddle-node bifurcation, homoclinic bifurcation and limit saddle-node bifurcation of limit cycle respectively. (b) Bifurcation diagram of fast subsystem with different concentration of calcium ion, in which the black curve indicates the concentration of calcium ion at [Ca1] = 0.034 nS, and the red curve indicates [Ca1] = 0.8 nS
图4. (a) 快子系统的分岔曲线,系统轨线(绿色)也叠加在上面。簇放电的类型为“fold/homoclinic”型簇放电,其中gNa=5nS,supH1与supH2是Hopf分岔,F1、F2、HC和LPC分别表示鞍结分岔、同宿轨分岔和极限环的鞍结分岔。(b) 不同钙离子浓度值对应的快子系统对慢变量的分岔曲线,其中黑色曲线表示钙离子浓度[Ca1] = 0.034 nS,红色曲线表示钙离子浓度[Ca1] = 0.8 nS
之后轨线的振幅逐渐增大并受极限环的吸引重复振荡,最后经极限环的同宿轨分岔(HC)跃迁至静息态,所以簇I为“fold/homoclinic”型簇放电,如图4(a)。而在钙离子浓度波动阶段,快子系统的平衡点曲线不断产生变化,随着钙离子浓度升高,平衡点及极限环左移,且静息态对应的平衡点上移,放电态对应的极限环振幅下移,见图4(b)所示。簇II的产生对应钙离子浓度波动时期,此时,当轨线的静息态经由平衡点的Hopf分岔跃迁到放电态,钙离子浓度升高,极限环左移,系统轨线的放电态稳定在极限环右侧的平衡点上。之后钙离子浓度下降,极限环右移,系统轨线在稳定的极限环间振荡,并由极限环的同宿轨分岔跃迁至静息态。
4. 混合簇的产生机制
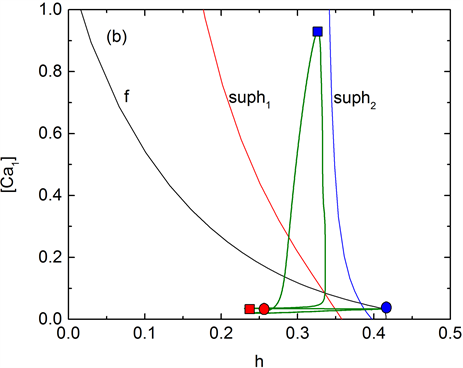
快子系统(1a)、(1b)和(1d)在 -平面上的双参数分岔如图5(b)所示。f是平衡点的鞍结分岔曲线(黑色),suph1和suph2分别表示平衡点的Hopf分岔点supH1和supH2对应的分岔曲线(红色和蓝色)。全系统的钙振荡曲线(绿色)也叠加在 -平面上。混合簇的形成受到平衡点的鞍结分岔(F1)与Hopf分岔(supH1, supH2)相对位置的影响。Hopf分岔曲线suph1水平位置位于曲线suph2左侧。当f水平位置位于suph2右侧时,簇Ⅰ产生,此时钙离子浓度 介于0~0.05 nS之间。而当f水平位置位于suph1与suph2之间, 处于0.05~0.08 nS之间,系统轨线簇II的静息态将通过鞍结分岔(F1)跃迁至放电态。混合簇与f,suph1及suph2位置关系见图5(b)所示。钙离子周期性的波动导致F与supH1、supH2相对位置发生了改变,使得三条曲线f,suph1和suph2产生了交叉,从而使得簇放电呈现出混合模式。
5. 结论
耦合Pre-Bötzinger复合体神经元中,细胞内钙离子是影响其放电模式的关键因素之一。系统呈现出两种类型簇交替出现的情形,即混合簇放电,其中簇I对应于钙离子浓度变化极小的时间段,而簇II (图3b)是由于钙离子浓度出现剧烈波动时产生的,此时平衡点、鞍结分岔、Hopf分岔位置发生了极大改变,
导致簇II与簇I有所不同。随着钠离子电导 升高,簇放电模式发生了一定改变,但都会受到 周
期性的波动影响。综上所述,混合簇放电模式的产生是由钙离子受到未失活的IP3调节,产生周期性的波动,从而使得快子系统的平衡点位置变化以及鞍结分岔、Hopf分岔相对位置发生改变,导致簇放电模式
 (a)
(a) 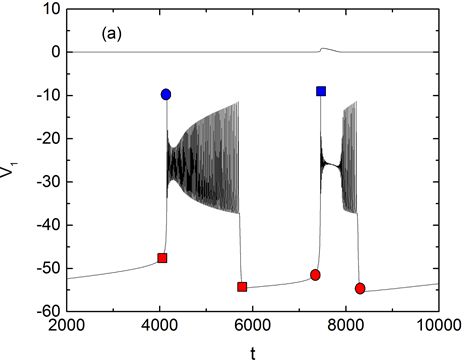 (b)
(b)
Figure 5. gNa = 5 ns, (a) Mixed-mode bursting. (b) Two parameter bifurcation diagram in -plane. Curves f, suph1 and suph2 are saddle-node bifurcation and Hopf bifurcation respectively. Changes of concentration of calcium ion [Ca1] with time t (green) is also superimposed on the diagram. The red square and the blue dot indicate the position of the membrane potential and concentration at different time t.
图5. gNa = 5 nS时,(a) 混合簇放电模式,(b) 双参数平面中的分岔图,f,suph1与suph2分别表示鞍结分岔曲线与Hopf分岔曲线,钙离子浓度[Ca1]随时间的变化曲线(绿色)也叠加在图中。红色方块和蓝色圆点标志分别表示不同时刻下膜电位V1和[Ca1]浓度所处的位置
的改变,呈现出混合簇同步放电模式。在哺乳动物的呼吸节律中常常产生混合模式簇放电,因此我们对混合簇及其产生机制的研究对认识Pre-Bötzinger复合体中大规模网络的动力学有着重要的意义,为进一步探索呼吸节律的产生机制提供有益的思考。
基金项目
本文得到国家自然科学基金(项目编号:11472009)北方工业大学“科研创新团队建设计划”项目(项目编号:XN018010)及北方工业大学大学生科技活动的资助。
文章引用
王子剑,马 芬,梁桐桐. 耦合Pre-BO¨ tzinger复合体神经元中混合簇放电模式的研究
Mixed Bursting of Coupled Neuron in the Pre-BO¨ tzinger Complex[J]. 动力系统与控制, 2018, 07(03): 154-161. https://doi.org/10.12677/DSC.2018.73017
参考文献
- 1. Best, J., Borisyuk, A., Rubin, J., et al. (2005) The Dynamic Range of Bursting in a Model Respiratory Pacemaker Network. SIAM Journal on Applied Dynamical Systems, 4, 1107-1139.
https://doi.org/10.1137/050625540 - 2. Ramirez, J.M., Quellmalz, U.J. and Richter, D.W. (1996) Postnatal Changes in the Mammalian Respiratory Network as Revealed by the Transverse Brain Stem Slice of Mouse. J. Physiol., 491, 799-812.
https://doi.org/10.1113/jphysiol.1996.sp021258 - 3. Ramirez, J.M., Koch, H., Garcia III, A.J., Doi, A. and Zanella, S. (2011) The Role of Spiking and Bursting Pacemakers in the Neuronal Control of Breathing. The Journal of Physiology, 37, 241-261.
https://doi.org/10.1007/s10867-011-9214-z - 4. Smith, J.C., Ellenberger, H.H., Ballanyi, K., Richter, D.W. and Feldman, J.L. (1991) Pre-Bötzinger Complex: A Brain Stem Region That May Generate Respiratory Rhythm in Mammal. Science, 254, 726-729.
https://doi.org/10.1126/science.1683005 - 5. Ivanchenko, M.V., Osipov, G.V., Shalfeev, V.D., et al. (2004) Phase Synchronization in Ensembles of Bursting Oscillators. Physical Review Letters, 93, 101-134.
https://doi.org/10.1103/PhysRevLett.93.134101 - 6. Ivanchenko, M.V., Osipov, G.V., Shalfeev, V.D., et al. (2007) Network Mechanism for Burst Generation. Physical Review Letters, 98, 101-108.
https://doi.org/10.1103/PhysRevLett.98.108101 - 7. Pace, R.W., Mackay, D.D., Feldman, J.L. and Negro, C.A.D. (2007) Role of Persistent Sodium Current in Mouse Pre-Bötzinger Complex Neurons and Respiratory Rhythm Generation. The Journal of Physiology, 580, 485-496.
https://doi.org/10.1113/jphysiol.2006.124602 - 8. Wang, Y.Y., Jonathan, E. and Rubin, J. (2016) Multiple Timescale Mixed Bursting Dynamics in a Respiratory Neuron Model. Journal of Computational Neuroscience, 41, 245-268.
https://doi.org/10.1007/s10827-016-0616-6 - 9. Mironov, S.L. (2008) Metabotropic Glutamate Re-ceptors Activate Dendritic Calcium Waves and TRPM Channels Which Drive Rhythmicre Spiratory Patterns in Mice. Journal of Physiology, 586, 2277-2291.
https://doi.org/10.1113/jphysiol.2007.149021 - 10. Toporikova, N. and Butera, R.J. (2011) Two Types of Inde-pendent Bursting Mechanisms in Inspiratory Neurons: An Integrative Model. Journal of Computational Neuroscience, 30, 515-528.
https://doi.org/10.1007/s10827-010-0274-z
附录
模型(1a)~(1f)中,变量和参数的表达式或数值(附表1):
(2a)
(2b)
(2c)
(2d)
(2e)
(2f)

Table 1. Parameter values in model 1(a)-1(f)
附表1. 模型1(a)~1(f)中参数值