Advances in Applied Mathematics
Vol.
07
No.
11
(
2018
), Article ID:
27553
,
12
pages
10.12677/AAM.2018.711161
The Dynamical Behavior of Dengue Virus Transmission Model with Maturation Delay and Incubation Delay
Yu Zhou, Tingting Zheng*
College of Mathematics and System Sciences, Xinjiang University, Urumqi Xinjiang
Received: Oct. 27th, 2018; accepted: Nov. 7th, 2018; published: Nov. 14th, 2018

ABSTRACT
The dynamical behavior of dengue virus transmission model with incubation delay and relatively long maturation delay compared to the average life span of adult mosquito is investigated. The positivity of solutions is showed by using contradiction. The local stability of the disease free equilibrium and endemic equilibrium are obtained by using the linearization method. Further, the global asymptotic stability of the positive equilibrium is proved by using Lyapunov functional method.
Keywords:Dengue, Incubation Delay, Maturation Delay, Disease-Free Equilibrium and Endemic Equilibrium, Local and Global Stability
具有孵化期时滞和潜伏期时滞的登革热病毒传播动力学模型研究
周瑜,郑庭庭*
新疆大学数学与系统科学学院,新疆 乌鲁木齐
收稿日期:2018年10月27日;录用日期:2018年11月7日;发布日期:2018年11月14日

摘 要
考虑到登革热病毒传播过程中在蚊子和人体内都具有一定的潜伏期,以及相对于蚊子的生命周期来说蚊子有一个较长的孵化期,本文考虑了一类具有潜伏期时滞和孵化期时滞的登革热病毒传播动力学模型。利用反证法证明了模型解的正性,通过线性化方法得到模型无病平衡点和地方病平衡点的局部渐近稳定性,给出了刻画疾病消除或流行的阈值条件。进一步,通过构造合适的李雅普诺夫泛函,得到模型无病平衡点和地方病平衡点的全局渐近稳定性的判别准则。
关键词 :登革热,潜伏期时滞,孵化期时滞,无病平衡点和地方病平衡点,局部稳定性和全局稳定性

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
最近几十年来,由于登革热的高发病率和高死亡率,它已成为当前威胁人类并跨越国界的最重要的虫媒疾病之一。该媒介传染病主要发生在热带,亚热带和温带国家,每年受感染者至少5000~10,000万,人主要通过埃及伊蚊和白纹伊蚊感染登革热病毒,主要症状为高烧、头疼、眼睛疼痛、关节疼痛、呕吐等症状 [1] [2] [3]。早在1981年,Jousset就找到了埃及伊蚊菌株和登革热病毒的起源地 [4]。为了更好地描述登革热病毒的影响,许多学者已经提出数学模型来研究登革热病毒的传播(看 [5] [6] [7] 及其引用的文献)。特别地,Esteva等人提出的登革热病毒传播模型分析了平衡点的全局稳定性,并就阈值条件讨论了媒介的控制策略 [8]。进一步,Wang等人提出了一个登革热病毒传播的非局部的时滞扩散模型,并建立了关于基本再生数的动力学性态 [9]。此外,Garba等人提出了登革热菌株传播动力学的定性模型,他们证明了当基本再生数小于1时无病平衡点的存在性和局部渐近稳定性,作者也发现模型会出现后向分支的现象 [10]。
如何控制和消灭登革热已经是一个热点话题。尽管有很多疫苗在开发,其中包括四价登革热疫苗已经被推荐使用于疾病高传染性的国家和地区,登革热仍然没有具体的治疗方法 [1]。在疫苗的功效没有被确定之前,登革热的控制策略主要采取适当的防护措施。主要防护措施为减少蚊子机制和个人的自我保护。减少蚊子的数量可以通过消除蚊子的繁殖地(如清洁暗沟、路边的水道,处理固体废物,消除人类制造的废弃场所等)和杀成虫药(通过播撒杀虫剂杀死成年蚊子) [11] [12]。另一方面,个人的自我保护依赖于防止蚊子的叮咬(如使用纱窗,穿长袖衣服,利用杀虫剂处理过的线圈和汽化器等物质。尽管有社区参与和媒介控制的协作与积极的疾病监测和杀虫剂的使用,在全国范围内对登革热预防和控制的成功案例还是很少的 [1]。此外,随着媒介抵抗力的增加,再感染的几率也随之加大。另外,由于杀虫剂产品开发与研究的高成本和低回报,在市场上仅供有极少的杀虫剂产品。
临床研究表明,蚊子的生命周期约为3周,登革热病毒在蚊子体内会有一个潜伏期,时间一般在5~8天,登革热病毒在人体内也会有一个潜伏期,时间一般在3~14天 [1]。然而,在现有的大部分研究工作中,考虑到理论分析的便利性,往往忽略了登革热病毒在人和蚊子体内的潜伏期时滞,这将高估疾病传播的风险。近年来已有部分学者研究具有时滞的媒介传染病模型,例如Zhao等人讨论了具有时滞的登革热模型,给出疾病消除的阈值条件及地方病平衡点全局吸引的充分条件 [13]。阮士贵等人研究了疟疾在媒介和宿主均带有潜伏期时滞的疟疾传播动力学模型,讨论了潜伏期时滞对阈值和疾病传播动态的影响 [14]。
另一方面,蚊子的生命周期有三个连续的水生幼年阶段(卵,幼虫,蛹)和成虫蛹阶段 [1]。蚊子从卵到成虫的发育时间(1~2周)经常用来和成年蚊子的平均寿命比较。蚊子种群的大小受到温度的强烈影响,并且由于季节变化母蚊子的数量也会相应地变化 [15] [16]。因此,考虑蚊子的孵化期(从卵发育到成年蚊子的幼年阶段的长度)及其对登革热病毒传播的影响是十分重要的。
基于上述讨论,本文构建了一类具有潜伏期时滞和孵化期时滞的登革热病毒传播动力学模型,讨论了模型无病平衡点和地方病平衡点的全局渐近稳定性,给出了确定疾病消除或流行的阈值条件。
2. 模型构建及其基本性质
本文,我们提出一个研究登革热病毒传播的模型。该模型将人分为易感类、感染类和康复类,分别用
,
,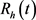 表示,将蚊子分为易感类、感染类,分别用
,
表示。人和蚊子的总数分别用
,
表示,将蚊子分为易感类、感染类,分别用
,
表示。人和蚊子的总数分别用
,
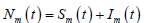 。取一个Ricker型函数,在文献 [17] 和文献 [18] 中就选取过Ricker型函数来建模。蚊子的孵化期,即从卵发育成成年蚊子这个过程所需要的平均时间用常数
表示,幼蚊的死亡率用
表示。用
表示蚊子的出生率函数,其中每只蚊子的最大蚊卵日产率用
表示;忽略孵化期时滞产卵量最大时蚊子的数量用
表示。
表示
单位之前蚊子产卵的个数,
反映了潜伏期内幼蚊的存活率。即使蚊卵在潜伏期内存活下来,现时的环境也决定着它们能否转变为成年蚊子,因此在当前时间Ricker型出生率函数
。取一个Ricker型函数,在文献 [17] 和文献 [18] 中就选取过Ricker型函数来建模。蚊子的孵化期,即从卵发育成成年蚊子这个过程所需要的平均时间用常数
表示,幼蚊的死亡率用
表示。用
表示蚊子的出生率函数,其中每只蚊子的最大蚊卵日产率用
表示;忽略孵化期时滞产卵量最大时蚊子的数量用
表示。
表示
单位之前蚊子产卵的个数,
反映了潜伏期内幼蚊的存活率。即使蚊卵在潜伏期内存活下来,现时的环境也决定着它们能否转变为成年蚊子,因此在当前时间Ricker型出生率函数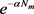 含有
。
,
分别表示感染类人在
和感染类蚊子在
的存活率,其中
,
分别表示登革热病毒在蚊子和人体内的潜伏期,
,
分别表示人和蚊子的死亡率。用
表示人的补充率,用a表示蚊子的平均叮咬率,用
表示人的康复率,感染类人将病毒传播给易感类蚊子的传播率用
表示,感染类蚊子将病毒传播给易感类人的传播率用
表示。由上述假设我们得到下列模型
含有
。
,
分别表示感染类人在
和感染类蚊子在
的存活率,其中
,
分别表示登革热病毒在蚊子和人体内的潜伏期,
,
分别表示人和蚊子的死亡率。用
表示人的补充率,用a表示蚊子的平均叮咬率,用
表示人的康复率,感染类人将病毒传播给易感类蚊子的传播率用
表示,感染类蚊子将病毒传播给易感类人的传播率用
表示。由上述假设我们得到下列模型
(1)
初始条件为
,
,
,
,
, (2)
其中,
,
i = 1, 2,
,
是正的连续函数。
模型(1)蚊子的总数满足下列方程:
, (3)
初始条件为
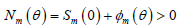 ,
。 (4)
,
。 (4)
令
, (5)
由此可知
是方程(3)的唯一正解,该解存在当前仅当
。定义
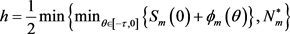 和
,则描述(3)的全局动力学行为的定理如下。
和
,则描述(3)的全局动力学行为的定理如下。
定理1:以(4)为初始条件的模型(3),对任意的
,解
是正的且
1) 若
,则对任意的
,有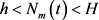 ,此外,存在一个全局渐近稳定的正解
。
,此外,存在一个全局渐近稳定的正解
。
2) 若
,则解
是有界的,平衡点
关于初始条件是全局渐近稳定的。
证:我们利用反证法证明对任意的
,
。注意到对任意的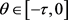 ,
,假设存在一个
,使得
,对
,
,则有
。由(3)可得
,
,假设存在一个
,使得
,对
,
,则有
。由(3)可得
,矛盾。因此对任意的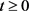 ,
。
,
。
下面我们证明情况(i)和(ii)。
情况(i) 设
。令
,
,
,则有
,
,我们断言对任意
,
。否则存在
,使得
,对任意的
,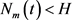 。又因为
,故由(3)得
。又因为
,故由(3)得
,
由
的定义知
,矛盾。因此,对任意的
,
。类似地,我们假设存在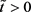 使得
,对任意的
,
,且有
。又
,故再由(3)得
使得
,对任意的
,
,且有
。又
,故再由(3)得
,
矛盾。因此,对任意的
,
。
情况(ii) 设
。L定义如情况(i),则
。假设存在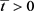 ,使得
,对任意的
,使得
,对任意的
,
Nm(t) < L。且
。由(3)得
,
矛盾。因此对任意的
,
。
为了证明
的全局渐近稳定性,我们记(3)的右边为
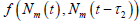 。因为
,所以
。因为
,所以
模型(3)产生由定义在
上的连续函数形成的空间C上的逐点排序的强单调半流 [19]。若
,存在一个平衡点
。由定理2.3.1 [19] 知
是全局渐近稳定的。若
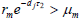 ,则有两个平衡点
和
。由定理2.3.2 [19] 知,(3)的解收敛于这两个平衡点中的一个。为了排除
是吸引子的可能,我们在
处线性化模型并利用定理A2 [20] ,可得到当
时,它是不稳定的。因此,当
时,
。
,则有两个平衡点
和
。由定理2.3.2 [19] 知,(3)的解收敛于这两个平衡点中的一个。为了排除
是吸引子的可能,我们在
处线性化模型并利用定理A2 [20] ,可得到当
时,它是不稳定的。因此,当
时,
。
模型(1)人的总数满足下列方程:
, (6)
可得
。
由于模型(1)中的其余四个方程都不含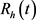 ,故模型(1)等价于如下极限系统
,故模型(1)等价于如下极限系统
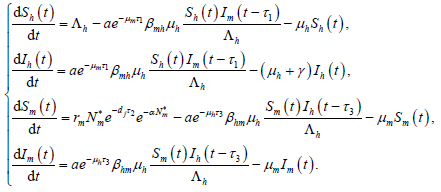 (7)
(7)
令
,易证
Ω 是模型(7)的正不变集。
3. 平衡点的存在性和无病平衡点的局部渐近稳定性
由文献 [21] 中的基本再生数的计算方法可得
。
易得模型(7)当
时,存在唯一的无病平衡点:
;当
时,除无病平衡点
外,模型(7)存在唯一的地方病平衡点: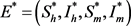 ,
,
其中
,
,
,
。
下面得到无病平衡点的局部渐近稳定性。
定理2:若
,则模型(7)的无病平衡点
是局部渐近稳定的;若
,则该无病平衡点是不稳定的。
证:在
处线性化,令
,
,
,
。
得到如下线性化模型
(8)
寻找指数解
,
,
,
,
的稳定性取决于特征方程(9)的特征根
(9)
显然,
,
是模型(7)的两个负特征根,其余的根由方程(10)决定
。 (10)
下面证明方程(10)的所有特征根具有负实部
,
,
则只需证明方程
的所有特征根具有负实部即可。容易得到
,
。
当
时,
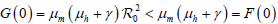 。而
关于
单调递增,
关于
单
。而
关于
单调递增,
关于
单
调递减。
,
,则方程(10)的复数根的实部都是非正的,根据Rouche定理 [22]
知,若对时滞
,
,方程(10)存在纯虚根,则导致稳定性改变。假设方程(10)存在这样的纯虚根
,我们有
。 (11)
令
y = ω 2,则有
。 (12)
易知方程(12)无正根,故不存在满足条件的
,使得
是方程(10)的解。
综上,模型(7)的无病平衡点局部渐近稳定。
4. 无病平衡点的全局吸引性
下证无病平衡点的全局吸引性。由模型(7)的第一个和第三个方程得
,
(13)
从而有
,
。 (14)
由模型(7)和(14)得
, (15)
,
由方程(15)得
,
从而有
, (16)
同理可得
。 (17)
由(16)和(17)得
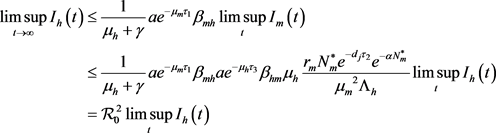 。
。
当
R0M< 1 时,
有界,故有
,
用同样的方法可证
,
再由定理2无病平衡点的局部稳定性可知
是全局渐近稳定的。
5. 地方病平衡点的全局渐近稳定性
下面证明地方病全局渐近稳定性
定理3:若
,则模型(7)的地方病平衡点
是全局渐近稳定的。
证:在
处线性化,令
,
E*,
,
,
得到如下线性化系统
(18)
寻找指数解
,
,
, 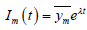 ,
,
的稳定性取决于特征方程(19)的特征根
(19)
显然,
是模型(7)的一个负特征根,其余的特征根由方程(20)决定
, (20)
其中
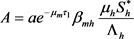 ,
,
,
,
,
。
下面证明方程(20)的所有特征根具有负实部。
令
,
。
故只需证明方程
的所有特征根具有负实部即可。
容易得到
,
,
矛盾。根据Rouche定理 [22] ,方程(20)不可能存在具有非负实部的特征根。
因此,当
时,模型(7)的地方病平衡点是局部渐近稳定的。
下证地方病平衡点的全局渐近稳定性
构造Lyapunov泛函:
,
其中
,
,
而
,
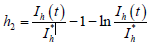 ,
,
,
,
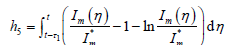 ,
。
,
。
则有
,
,
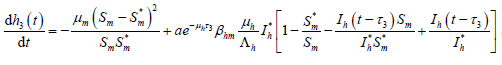 ,
,
,
,
。
从而
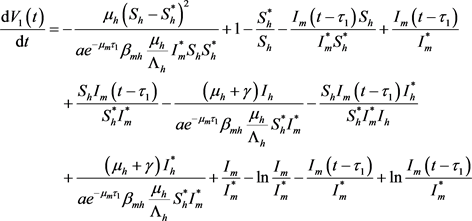 ,
,
故
(21)
令
。
需证
中最大不变集是
。由
,有
,
解上式得
。
同理可得
由(21)知,当
时,
,
。由拉萨尔不变原理知
地方病平衡点
是全局渐近稳定的。
文章引用
周瑜,郑庭庭. 具有孵化期时滞和潜伏期时滞的登革热病毒传播动力学模型研究
The Dynamical Behavior of Dengue Virus Transmission Model with Maturation Delay and Incubation Delay[J]. 应用数学进展, 2018, 07(11): 1381-1392. https://doi.org/10.12677/AAM.2018.711161
参考文献
- 1. Centers for Disease Control and Prevention. http://www.cdc.gov/dengue/
- 2. Derouich, M. and Boutayeb, A. (2006) Dengue Fever: Mathematical Modelling and Computer Simulation. Applied Mathematics and Computation, 177, 528-544. https://doi.org/10.1016/j.amc.2005.11.031
- 3. Esteva, L. and Vargas, C. (2003) Coexistence of Different Serotypes of Dengue Virus. Journal of Mathematical Biology, 46, 31-47. https://doi.org/10.1007/s00285-002-0168-4
- 4. Jousset, F.X. (1981) Geographic Aedes aegypti Strains and Dengue-2 Virus: Susceptibility, Ability to Transmit to Vertebrate and Transovarial Transmission. Annales de l’Institut Pasteur/Virologie, 132, 357-370.
https://doi.org/10.1016/S0769-2617(81)80006-8
- 5. Aldila, D., Gotz, T. and Soewono, E. (2013) An Optimal Control Problem Arising from a Dengue Disease Transmission Model. Mathematical Biosciences, 242, 9-16. https://doi.org/10.1016/j.mbs.2012.11.014
- 6. Kooi, B.W., Aguiar, M. and Stollenwerk, N. (2014) Analysis of an Asymmetric Two-Strain Dengue Model. Mathematical Biosciences, 248, 128-139. https://doi.org/10.1016/j.mbs.2013.12.009
- 7. Coutinhoa, F.A.B., Burattinia, M.N., Lopeza, L.F., et al. (2006) Threshold Conditions for a Non-Autonomous Epidemic System Describing the Population Dynamics of Dengue. Bulletin of Mathematical Biology, 68, 2263-2282.
https://doi.org/10.1007/s11538-006-9108-6
- 8. Esteva, L. and Vargas, C. (1998) Analysis of a Dengue Disease Transmission Model. Mathematical Biosciences, 150, 131-151. https://doi.org/10.1016/S0025-5564(98)10003-2
- 9. Wang, W. and Zhao, X.Q. (2011) A Nonlocal and Time-Delayed Reaction-Diffusion Model of Dengue Transmission. SIAM Journal on Applied Mathematics, 71, 147-168. https://doi.org/10.1137/090775890
- 10. Garba, S.M., Gumel, A.B. and Abu Bakar, M.R. (2008) Backward Bifurcations in Dengue Transmission Dynamics. Mathematical Biosciences, 215, 11-25. https://doi.org/10.1016/j.mbs.2008.05.002
- 11. Mishra, A. and Gakkhar, S. (2014) The Effects of Awareness and Vector Control on Two Strains Dengue Dynamics. Applied Mathematics and Computation, 246, 159-167. https://doi.org/10.1016/j.amc.2014.07.115
- 12. Yang, H.M. and Ferreira, C.P. (2008) Assessing the Effects of Vector Control on Dengue Transmission. Applied Mathematics and Computation, 198, 401-413. https://doi.org/10.1016/j.amc.2007.08.046
- 13. Wang, Z. and Zhao, X. (2012) Global Dynamics of a Time-Delayed Dengue Transmission Model. Canadian Applied Mathematics Quarterly, 20, 89-113.
- 14. Ruan, S., Xiao, D. and Beier, J.C. (2008) On the Delayed Ross-Macdonald Model for Malaria Transmission. Bulletin of Mathematical Biology, 70, 1098-1114. https://doi.org/10.1007/s11538-007-9292-z
- 15. Bayoh, M.N. and Lindsay, S.W. (2003) Effect of Temperature on the Development of the Aquatic Stages of Anopheles gambiae Sensu Stricto (Diptera: Culicidae). Bulletin of Entomological Research, 93, 375-381.
- 16. Shaman, J., Spiegelman, M., Cane, M., et al. (2006) A Hydrologically Driven Model of Swamp Water Mosquito Population Dynamics. Ecological Modelling, 194, 395-404. https://doi.org/10.1016/j.ecolmodel.2005.10.037
- 17. Cooke, K., Driessche, V.P. and Zou, X. (1999) Interaction of Maturation Delay and Nonlinear Birth in Population and Epidemic Models. Journal of Mathematical Biology, 39, 332-352. https://doi.org/10.1007/s002850050194
- 18. Velasco-Hernandez, J.X. (1994) A Model for Chagas Disease Involving Transmission by Vectors and Blood Transfusion. Theoretical Population Biology, 46, 1-31. https://doi.org/10.1006/tpbi.1994.1017
- 19. Smith, H. (1995) Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems. In: Mathematical Surveys and Monographs, Vol. 41, American Mathematical Society, Providence, 174 p.
- 20. Cooke, K.L. and Van, D.P. (1996) Analysis of an SEIRS Epidemic Model with Two Delays. Journal of Mathematical Biology, 35, 240-260. https://doi.org/10.1007/s002850050051
- 21. Van, D.P. and Watough, J. (2002) Reproduction Numbers and Sub-Threshold Endemic Equilibria for Compartmental Models of Disease Transmission. Mathematical Biosciences, 180, 29-48.
- 22. Elsgolts, L. and Norkin, S. (1973) An Introduction to the Theory and Application of Differential Equations with Deviating Argument. Academic Press, New York.
NOTES
*通讯作者。




 表示,将蚊子分为易感类、感染类,分别用
,
表示。人和蚊子的总数分别用
,
表示,将蚊子分为易感类、感染类,分别用
,
表示。人和蚊子的总数分别用
,
 。取一个Ricker型函数,在文献 [17] 和文献 [18] 中就选取过Ricker型函数来建模。蚊子的孵化期,即从卵发育成成年蚊子这个过程所需要的平均时间用常数
表示,幼蚊的死亡率用
表示。用
表示蚊子的出生率函数,其中每只蚊子的最大蚊卵日产率用
表示;忽略孵化期时滞产卵量最大时蚊子的数量用
表示。
表示
单位之前蚊子产卵的个数,
反映了潜伏期内幼蚊的存活率。即使蚊卵在潜伏期内存活下来,现时的环境也决定着它们能否转变为成年蚊子,因此在当前时间Ricker型出生率函数
。取一个Ricker型函数,在文献 [17] 和文献 [18] 中就选取过Ricker型函数来建模。蚊子的孵化期,即从卵发育成成年蚊子这个过程所需要的平均时间用常数
表示,幼蚊的死亡率用
表示。用
表示蚊子的出生率函数,其中每只蚊子的最大蚊卵日产率用
表示;忽略孵化期时滞产卵量最大时蚊子的数量用
表示。
表示
单位之前蚊子产卵的个数,
反映了潜伏期内幼蚊的存活率。即使蚊卵在潜伏期内存活下来,现时的环境也决定着它们能否转变为成年蚊子,因此在当前时间Ricker型出生率函数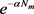 含有
。
,
含有
。
,
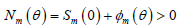 ,
。 (4)
,
。 (4)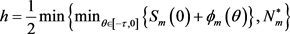 和
,则描述(3)的全局动力学行为的定理如下。
和
,则描述(3)的全局动力学行为的定理如下。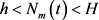 ,此外,存在一个全局渐近稳定的正解
。
,此外,存在一个全局渐近稳定的正解
。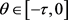 ,
,假设存在一个
,使得
,对
,
,则有
。由(3)可得
,
,假设存在一个
,使得
,对
,
,则有
。由(3)可得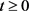 ,
。
,
。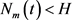 。又因为
,故由(3)得
。又因为
,故由(3)得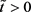 使得
,对任意的
,
,且有
。又
,故再由(3)得
使得
,对任意的
,
,且有
。又
,故再由(3)得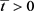 ,使得
,对任意的
,使得
,对任意的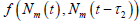 。因为
,所以
。因为
,所以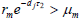 ,则有两个平衡点
和
。由定理2.3.2 [19] 知,(3)的解收敛于这两个平衡点中的一个。为了排除
是吸引子的可能,我们在
处线性化模型并利用定理A2 [20] ,可得到当
时,它是不稳定的。因此,当
时,
。
,则有两个平衡点
和
。由定理2.3.2 [19] 知,(3)的解收敛于这两个平衡点中的一个。为了排除
是吸引子的可能,我们在
处线性化模型并利用定理A2 [20] ,可得到当
时,它是不稳定的。因此,当
时,
。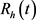 ,故模型(1)等价于如下极限系统
,故模型(1)等价于如下极限系统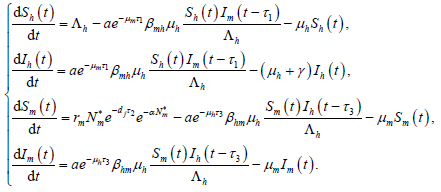 (7)
(7)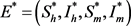 ,
,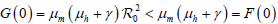 。而
关于
单调递增,
关于
单
。而
关于
单调递增,
关于
单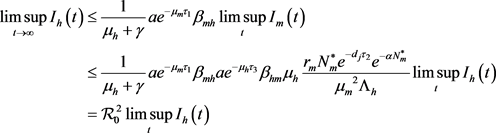 。
。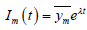 ,
,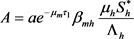 ,
,
,
,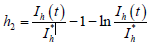 ,
,
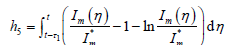 ,
。
,
。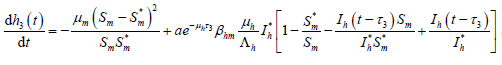 ,
,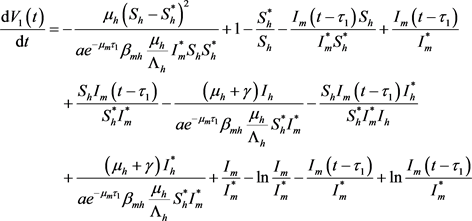 ,
,