Advances in Microbiology
Vol.
07
No.
04
(
2018
), Article ID:
27672
,
7
pages
10.12677/AMB.2018.74018
Cloning of Nudel Gene and Its Expression in COS7 Cells
Zijian Zhou1, Bangyi Wei1, Siyu Zeng1, Zhicheng Zou1, Peng Wang1, Yao Wang2, Jun Chen1*
1Bioengineering Department, Wuhan University of Science and Technology, Wuhan Hubei
2Life Science School, Huazhong University of Science and Technology, Wuhan Hubei

Received: Nov. 2nd, 2018; accepted: Nov. 15th, 2018; published: Nov. 22nd, 2018

ABSTRACT
In this study, Nudel gene was cloned and used to construct a eukaryotic expression vector harboring a red fluorescent gene. According to the Gene card data, the RPKM value of Nudel is
Keywords:Nudel Gene, cDNA clone, Gene Expression, Tumor Suppressor Gene Pten, COS7 Cell
Nudel基因的克隆及在COS7细胞中的 表达研究
周子建1,魏邦毅1,曾思宇1,邹志成1,王鹏1,汪瑶2,陈俊1*
1武汉科技大学生物工程教研室,湖北 武汉
2华中科技大学生命科学学院,湖北 武汉

收稿日期:2018年11月2日;录用日期:2018年11月15日;发布日期:2018年11月22日

摘 要
本实验克隆并构建了携带红荧光报告基因的Nudel真核表达载体,为后续研究Nudel基因的功能打下基础。查阅Gene card可知,Nudel基因在肺组织中的RPKM值为7.97,本研究以人肺癌细胞A549细胞为材料,提取总RNA并反转录出cDNA,再以cDNA为模板扩增Nudel基因;将目的基因与pMD18-T载体连接,测序鉴定并命名为pT-Nudel;然后再将Nudel基因亚克隆至含红荧光报告基因的载体pDsRed-C1,构建的表达载体命名为pRed-Nudel;提取去内毒素的超纯质粒,转染COS7细胞,检测Nudel的表达情况以及抑癌基因Pten对Nudel表达的影响。结果表明,克隆的Nudel基因经测序鉴定正确;将Nudel基因亚克隆至红荧光表达载体pDsRed-C1,成功构建pRed-Nudel,并在COS7中成功表达。同时观察到与突变型的PtenC124S相比,野生型的Pten有抑制Nudel表达的可能,需要后续的实验进一步验证。
关键词 :Nudel基因,cDNA克隆,基因表达,抑癌基因Pten,COS7细胞

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
Nudel是一类进化上保守的蛋白,在体内参与多个信号通路的调控与疾病的发生。Nudel与多种蛋白质相互作用,包括dynein、lissencephaly 1蛋白(Lis1)、精神分裂症1 (Disc1)和
2. 材料与方法
2.1. 材料
A549细胞、pDsRed-C1载体、感受态细胞DH5α由本实验室保存;pMD18-T、T4连接酶购自TaKaRa公司;内切酶Xho I、BamH I购自Thermo公司;胶回收试剂盒:E.Z.N.A Gel Extraction Kit(100)试剂盒;质粒提取试剂盒:E.Z.N.A. Plasmid Mini Kit I(100)试剂盒;超纯质粒提取试剂盒:E.Z.N.A Endo-free Plasmid Mini II试剂盒,Trizol、逆转录试剂盒M-MLV购自Invitrogen公司;转染试剂PEI购自Proteintech公司。
2.2. RT-PCR扩增Nudel目的基因
选取生长状态良好的A549细胞(培养条件:RPMI 1640培养基,10%胎牛血清,37℃,5% CO2)为材料,提取总RNA。以此RNA为模板,参照Invitrogen的说明书进行反转录获得cDNA。以cDNA为模板,扩增目的基因Nudel,扩增体系:ddH2O 15.5 μL,cDNA 0.5 μL,上下游引物各0.5 μL,dNTPs (
2.3. 构建pT-Nudel和pDsRed-Nudel载体
将扩增的目的基因克隆至pMD18-T载体上,进行测序鉴定,正确的克隆命名为pT-Nudel。以测序鉴定后的质粒pT-Nudel为模板,采用高保真酶扩增Nudel目的基因。扩增体系:ddH2O 34 μL,pT-Nudel质粒DNA稀释100倍取1 μL (约50 ng),上下游引物各1 μL,dNTPs (
2.4. Nudel基因在真核细胞中COS7中的表达
制备去内毒素的超纯pDsRed-Nudel质粒,检测DNA的浓度与纯度,转染到COS7细胞,并在荧光显微镜下进行观察。细胞转染:采用PEI转染试剂进行转染,12孔板种细胞过夜贴壁,DNA与转染试剂比例1:3 (μg:μL),转染步骤参照PEI说明书进行,转染后30小时置于显微镜下检测。
3. 结果与分析
3.1. A549细胞总RNA的提取与Nudel基因的扩增
培养A549细胞,选取生长状态良好的A549细胞为材料,提取总RNA。RNA浓度测定表明,1号样品浓度为730 ng/μL,A260/280比值为1.98,2号样品浓度为695 ng/μL,A260/280比值为1.97,说明提取的RNA 纯度比较高。提取的RNA进行电泳检测,结果见图1(A)。可以看到清晰的三条带,分别是28 S,18 S和5.8 S,表明提取的RNA完整性良好,可以用于反转录。
以总RNA为模板,反转录获得cDNA,再以cDNA为模板,扩增目的基因 Nudel。扩增结果见图1(B),电泳条带单一,对比DNA Marker,扩增的目的条带大小在1000 bp附近,与NCBI数据库里提供的Nudel基因(1038 bp)大小一致,初步判断扩增产物是目标产物,也说明A549细胞中Nudel基因有较高的表达水平。
3.2. 构建pT-Nudel和pDsRed-Nudel载体
将Nudel基因插入pMD18-T载体中,转化大肠杆菌DH5α,通过菌落PCR筛选阳性克隆,菌落PCR扩增结果见图2(A),在对应Maker1000 bp的位置有条带出现,与cDNA做模板扩增的条带大小一致。
 (A)
(A) 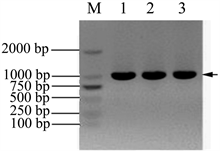 (B)
(B)
Figure 1. Amplification Nudel gene by RT-PCR. (A) The total RNA was extracted from A549 cells: 1 & 2 samples were the total RNA extracted from A549 cells; (B) Amplification of Nudel gene from the cDNA: M: DNA marker; 1, 2 & 3 samples were Nudel gene fragments amplified from the cDNA (about 1000 bp)
图1. RT-PCR扩增Nudel基因。(A) A549细胞总RNA提取电泳检测:1 & 2样品均为提取的A549细胞总RNA;(B)从A549细胞cDNA中扩增Nudel基因:M:DNA marker;1、2、3均为扩增的Nudel基因片段
 (A)
(A) 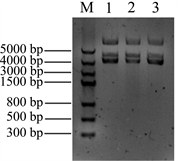 (B)
(B)
Figure 2. Construction the recombinants of pT-Nudel. (A) Identification the recombinant pT-Nudel by cloning PCR: M: DNA marker, 1, 2 & 3samples were the cloning PCR of recombinant pT-Nudel; (B) Isolation the recombinant plasmid pT-Nudel: M: DNA marker, 1, 2 & 3 samples were the recombinant pT-Nudel plasmids
图2. 构建重组质粒pT-Nudel。(A) 重组质粒pT-Nudel 菌落PCR鉴定:M:DNA marker,1、2&3为pT-Nudel菌落PCR;(B) 提取pT-Nudel重组质粒DNA:M:DNA marker,1、2&3为提取的重组质粒pT-Nudel
提取重组质粒DNA,电泳检测重组质粒大小,结果见图2(B),提取的质粒呈现3条带,中间的条带为质粒的线性条带,对比Marker,该线性条带在3000 bp和5000 bp之间,pMD18-T质粒大小为2692 bp,Nudel基因大小为1038 bp,重组质粒理论大小为3730 bp,从电泳结果判断,所提取的重组质粒大小正确,将正确的克隆进一步进行测序定。将测序结果在NCBI中用BLAST进行比对分析,本实验从A549肺泡细胞中所克隆的Nudel基因序列与NCBI数据库中的Nudel基因匹配度为100%,序列完整,并且没有发生碱基突变。
在此基础上,进一步将Nudel基因亚克隆至携带红荧光的真核表达载体pDsRed-C1上,亚克隆的限制性酶切位点为Xho I和BamH I,构建重组质粒pDsRed-Nudel。提取重组质粒,电泳检测见图3(A),pDsRed-C1载体大小为4700 bp,Nudel基因大小1038 bp,那么重组质粒pDsRed-Nudel的大小为5738 bp,由图3(A)中性条带大小判断,所提取的重组质粒大小正确。将提取的重组质粒pDsRed-Nudel用Xho I和BamH I进行双酶切鉴定,电泳结果显示(见图3(B)),在1000 bp和5000 bp的位置分别有条带出现,与预期相符,1000 bp附近的是Nudel基因片段,5000 bp附近的是载体片段。由此可见,Nudel基因成功的亚克隆到了红荧光表达载体上,构建了pDsRed-Nudel载体。
3.3. 重组质粒pDsRed-Nudel转染COS7细胞
利用PEI转染试剂,将重组质粒pDsRed-Nudel转染COS7细胞,检测其表达情况。结果如下(见图4),在平行转染条件下,pDsRed-C1空载体转染COS7细胞(见图4(B)),转染效率较重组质粒pDsRed-Nudel要高(见图4(D))。重组质粒pDsRed-Nudel转染COS7细胞的效率虽然较低,但是仍然观测到了荧光,初步判断pDsRed-Nudel在COS7细胞中能够表达。
 (A)
(A) 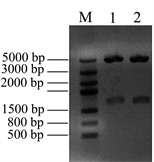 (B)
(B)
Figure 3. Construction the eukaryotic expression vector pRed-Nudel. (A) Isolation the recombinant pRed-Nudel plasmids: M: DNA marker; 1, 2, 3 & 4 samples were the recombinant pRed-Nudel plasmids; (B) Digestion the recombinant pRed-Nudel by restriction enzyme Xho I and BamH I: M: DNA marker; 1 & 2 samples were the recombinant pRed-Nudel digested by Xho I and BamH I
图3. 构建真核表达载体pRed-Nudel。(A) 提取重组质粒pRed-Nudel:M:DNA marker;1、2、3&4为提取的pRed-Nudel重组质粒;(B) 双酶切鉴定重组质粒pRed-Nudel:M:DNA marker;1&2为重组质粒pRed-Nudel双酶切
3.4. 抑癌基因Pten及其突变体对Nudel基因表达的影响
本实验克隆Nudel的目的,是为了考察Nudel对肿瘤细胞迁移的影响,而Pten基因是著名的肿瘤抑制基因,因此本实验将野生型的Pten和突变型的Pten即C124SPten (Pten第124位的氨基酸C突变成S),分别与Red-Nudel共转染COS7细胞,并在荧光显微镜下进行观察。结果发现,与突变型的C124SPten相比(见图5(D)),野生型的Pten与Red-Nudel共转染(见图5(B)),红荧光更少,推测124位的突变对Nudel的表达存在一定的影响。
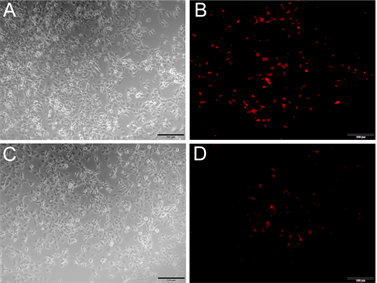
Figure 4. Transfection the expression vector pRed-Nudel into COS7 cells. (A) (B) Transfection the vector pDsRed-C1 into COS
图4. 重组质粒pRed-Nudel转染COS7细胞。(A) (B) pDsRed-C1转染COS7细胞,(A)图为白光,(B)图为激发光;(C) (D) pRed-Nudel转染COS7细胞,(C)图为白光,(D)图为激发光;注:图中比例尺为100 μm
4. 讨论
本实验以A549细胞为材料,成功获得目的基因Nudel,并构建携带红荧光报告基因的真核表达载体 pRed-Nudel。查阅Gene card可知,Nudel基因在骨髓普遍高表达,RPKM值为18.6,其次是肾上腺,RPKM
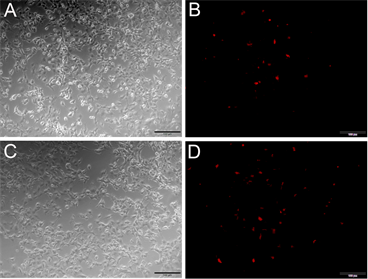
Figure 5. Co-transfection Pten or PtenC124S and pRed-Nude into COS7 cells. (A) (B) Co-transfection Pten and pRed-Nudel into COS7 cells, (A) At common optical path; (B) At fluorescence optical path; (C) (D) Co-transfection PtenC124S and pRed-Nudel into COS7 cells, (C) At common optical path; (D) At fluorescence optical path; Note: bar = 100 μm
图5. Pten和PtenC124S与pRed-Nudel共转染COS7细胞。(A) (B) Pten和pRed-Nudel共转染COS7细胞,(A)图为白光,(B)图为激发光;(C) (D) PtenC124S和pRed-Nudel共转染COS7细胞,(C)图为白光,(D)图为激发光;注:图中比例尺为100 μm
值为13.4。在另外的25种组织中均不同程度检测到Nudel的表达,具体表达情况见图6,在肺组织中的RPKM值约为7.97左右,RPKM值的大小显示的是该基因在相应的组织中表达量的高低。本实验所用的是A549细胞,该细胞为人肺泡上皮细胞。实验结果表明,Nudel基因在A549细胞中有表达,因此成功扩增到了该基因,测序结果显示,该基因在A549细胞株中,没有突变。

Figure 6. The Nudel gene expression level in different tissue
图6. Nudel基因在不同组织中的表达水平
Nudel是一种保守的中心体蛋白质,在细胞中绝大部分分布于细胞质,但在间期在中心体上有较强的定位,在有丝分裂期分布于整个纺锤体 [14] 。在体内,Nudel对于细胞运动的关键蛋白—胞质动力蛋白发挥功能至关重要,包括细胞内运输、有丝分裂和细胞迁移等。Nudel通过与Cdc42竞争结合Cdc42GAP,使得Cdc42释放而被活化,进而促进细胞迁移 [9] 。本实验初步尝试将野生型Pten和突变型PtenC124S与Red-Nudel共转染,观察到与突变型PtenC124S相比,野生型Pten对Red-Nudel的表达有抑制作用,需要进一步实验证实,如果Pten下调Nudel的表达,Nudel水平降低,则Nudel竞争结合Cdc42GAP水平降低,那么Cdc42GAP与Cdc42结合增加,则细胞迁移降低。也就是说,Pten通过下调Nudel进而抑制细胞迁移,这与Pten抑癌基因的功能是相符的 [15] 。
致谢
感谢湖北省自然科学基金资助项目(2015CFB490)的资助;感谢全国大学生创新创业大赛项目(201710488013)的资助。
文章引用
周子建,魏邦毅,曾思宇,邹志成,王 鹏,汪 瑶,陈 俊. Nudel基因的克隆及在COS7细胞中的表达研究
Cloning of Nudel Gene and Its Expression in COS7 Cells[J]. 微生物前沿, 2018, 07(04): 149-155. https://doi.org/10.12677/AMB.2018.74018
参考文献
- 1. Chansard, M., Hong, J.H., Park, Y.U., et al. (2011) Ndel1, Nudel (Noodle): Flexible in the Cell? Cytoskeleton, 68, 540-554. https://doi.org/10.1002/cm.20532
- 2. Vallee, R.B., McKenney, R.J. and Ori-McKenney, K.M. (2012) Multiple Modes of Cytoplasmic Dynein Regulation. Nature Cell Biology, 14, 224-230. https://doi.org/10.1038/ncb2420
- 3. Bradshaw, N.J., Hennah, W. and Soares, D.C. (2013) NDE1 and NDEL1: Twin Neurodevelopmental Proteins with Similar “Nature” but Different “Nurture”. Biomolecular Concepts, 4, 447-464. https://doi.org/10.1515/bmc-2013-0023
- 4. Klinman, E. and Holzbaur, E.L. (2015) Stress-Induced CDK5 Activation Disrupts Axonal Transport via Lis1/Ndel1/ Dynein. Cell Reports, 12, 462-473. https://doi.org/10.1016/j.celrep.2015.06.032
- 5. Moon, H.M., Youn, Y.H., Pemble, H., et al. (2014) LIS1 Controls Mitosis and Mitotic Spindle Organization via the LIS1-NDEL1-Dynein Complex. Human Molecular Genetics, 23, 449-466. https://doi.org/10.1093/hmg/ddt436
- 6. Bradshaw, N.J. and Hayashi, M.A. (2017) NDE1 and NDEL1 from Genes to (mal) Functions: Parallel but Distinct Roles Impacting on Neurodevelopmental Disorders and Psychiatric Illness. Cellular and Molecular Life Sciences, 74, 1191-1210. https://doi.org/10.1007/s00018-016-2395-7
- 7. Zhang, Q., Wang, F., Cao, J., et al. (2009) Nudel Promotes axonal Lysosome Clearance and Endo-Lysosome Formation via Dyne-in-Mediated Transport. Traffic, 10, 1337-1349. https://doi.org/10.1111/j.1600-0854.2009.00945.x
- 8. Lam, C., Vergnolle, M.A., Thorpe, L., et al. (2010) Functional Interplay between LIS1, NDE1 and NDEL1 in Dynein-Dependent Organelle Positioning. Journal of Cell Science, 123, 202-212. https://doi.org/10.1242/jcs.059337
- 9. Shen, Y., Li, N., Wu, S., et al. (2008) Nudel Binds Cdc42GAP to Modulate Cdc42 Activity at the Leading Edge of Migrating Cells. Developmental Cell, 14, 342-353. https://doi.org/10.1016/j.devcel.2008.01.001
- 10. Iijima, M. and Devreotes, P. (2002) Tumor Suppressor PTEN Mediates Sensing of Chemoattractant Gradients. Cell, 109, 599-610. https://doi.org/10.1016/S0092-8674(02)00745-6
- 11. Liliental, J., Moon, S.Y., Lesche, R., et al. (2000) Genetic Deletion of the Pten Tumor Suppressor Gene Promotes Cell Motility by Activation of Rac1 and Cdc42 GTPases. Current Biology, 10, 401-404. https://doi.org/10.1016/S0960-9822(00)00417-6
- 12. Mester, J. and Eng, C. (2013) When Overgrowth Bumps into Cancer: The PTEN-Opathies. American Journal of Medical Genetics Part C: Seminars in Medical Genetics, 163, 114-121. https://doi.org/10.1002/ajmg.c.31364
- 13. Kath, C., Goni-Oliver, P., Müller, R., et al. (2018) PTEN Suppresses Axon Outgrowth by Down-Regulating the Level of Detyrosinated Microtubules. PLoS One, 13, e0193257. https://doi.org/10.1371/journal.pone.0193257
- 14. Wang, S., Ketcham, S.A., Schön, A., et al. (2013) Nudel/NudE and Lis1 Promote Dynein and Dynactin Interaction in the Context of Spindle Morphogenesis. Molecular Biology of the Cell, 24, 3522-3533. https://doi.org/10.1091/mbc.e13-05-0283
- 15. Brandmaier, A., Hou, S., Demaria, S., et al. (2017) PTEN at the Interface of Immune Tolerance and Tumor Suppression. Frontiers in Biology (Beijing), 12, 163-174. https://doi.org/10.1007/s11515-017-1443-5
NOTES
*通讯作者。