Hans Journal of Soil Science
Vol.
07
No.
02
(
2019
), Article ID:
29545
,
12
pages
10.12677/HJSS.2019.72011
The Analysis of Vegetation and Soil Characteristics of Different Habitats in the Different Habitats of Henan County
Yanfang Deng1,2, Nengyang Xiao3, Yuanwu Yang3, Huakun Zhou2,4
1Qinghai Province Grassland Station, Xining Qinghai
2Key Laboratory of Restoration Ecology in Cold Region of Qinghai Province, Xining Qinghai
3Qinghai University College of Agriculture and Animal Husbandry, Xining Qinghai
4Northwest Plateau Institute of Biology, Chinese Academy of Sciences, Xining Qinghai
Received: Mar. 10th, 2019; accepted: Mar. 25th, 2019; published: Apr. 1st, 2019

ABSTRACT
To clarify the relationship between the characteristics of vegetation and soil environmental factors of alpine grassland, this paper studies the species diversity, the soil nutrient under different elevation changes, the soil nutrient of different vegetation and the habitat of the soil chemical properties of the different alpine habitats (shady slope, slope, wetlands, degradation and split). The results showed that species diversity of sunny slope, flat beach, shady slope, wetland and degraded land was decreasing gradually, and the contents of soil available N, available P, available K, total N, total P2O5 and total K2O were close to positive correlation. The highest soil nutrient content in habitat is the shady slope, followed by the sunny slope. The nutrient content in the flat beach soil is relatively low, and the degradation is higher than that in the wetland soil. The distribution of Potentilla fruticosa is about 3682 m higher, while the distribution of Goose down is lower than 3582 m. The soil nutrients of high-altitude habitats are relatively higher than those of low-altitude habitats. The soil nutrients of Potentilla fruticosa community are relatively high in different species.
Keywords:Alpine GRASSLAND, The Altitude, Soil Properties
河南县不同生境高寒草地植被与土壤特性分析
邓艳芳1,2,肖能洋3,杨元武3,周华坤2,4
1青海省草原总站,青海 西宁
2青海省寒区恢复生态学重点实验室,青海 西宁
3青海大学农牧学院,青海 西宁
4中国科学院西北高原生物研究所,青海 西宁
收稿日期:2019年3月10日;录用日期:2019年3月25日;发布日期:2019年4月1日

摘 要
为了阐明高寒草地植被特性与土壤环境因子的关系,本文研究了不同生境(阴坡、阳坡、湿地、退化和平滩)高寒草地物种多样性和不同海拔土壤养分的变化情况,以及不同植被土壤养分和各生境土壤的化学性质。结果表明,阳坡、平滩、阴坡、湿地和退化地物种多样性逐渐在减少,土壤速效N、速效P、速效K、全N、全P2O5和全K2O含量之间接近正相关,在不同生境中土壤养分含量最高的是阴坡,其次是阳坡,平滩土壤中养分含量是相对较低的,退化相对于湿地土壤养分是较高的。金露梅分布海拔较高约达3682 m,而鹅绒委陵菜分布海拔较低约3582 m,高海拔生境的土壤相对于低海拔生境的土壤养分相对略高,金露梅群落在不同物种中土壤养分相对较高。
关键词 :高寒草地,海拔高度,土壤特性

Copyright © 2019 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
植被与环境的关系一直是生态学研究的重点问题。随着人类活动和全球气候变化的加剧,全球生物多样性丧失及其对生态系统功能可能产生的影响引起人们普遍关注。作为世界上最为独特的生态地域单元,青海高寒草地的植被变化特征比其它地区更明显,周围地区的影响也可能更大。作为高寒草地的青海,研究土壤中化学成分对高寒草地生态系统的植被状况,生物量分配,对认识青海高原草地生态系统运行机制具有十分重要的意义。通过对植被覆盖变化与土壤的关系、不同尺度地上部生物量、地下部生物量以及根冠比与环境土壤关系的研究,确立环境效应。采用系统聚类和关联度数值分析方法,分析未退化、中度退化、重度退化和极度退化阶段下土壤养分和植被之间的关系。结果表明:当草地植被达到极度退化时,土壤养分仍保持着一定的稳定性;运用关联分析的计算表明,土壤容重和有机质对草地产量、盖度、密度有正的相关性,其关联度在0.69以上 [1] 。以青藏高原多年冻土区3种高寒草地植被为研究对象,设置6个样地,并结合附近活动层观测场环境因子数据,定量分析生物量与环境因子的关系。结果表明,各高寒草地地下生物量对总生物量的贡献率最大,而地下生物量在0~10 cm集中分布;对于总生物量和地下生物量,各因子影响程度大小次序为:土壤盐分 > 土壤含水量 > 空气温度,而对地上生物量,依次为土壤含水量 > 土壤盐分 > 空气温度;土壤温度同生物量存在负相关关系。同时,伴随多年冻土退化,活动层表层不同深度(10~50 cm)土壤温度明显升高,含水量逐渐降低,土壤盐分不断增加,从而使高寒草地植被类型出现由高寒沼泽草甸、高寒草甸至高寒草原的逆向演替过程,群落总盖度及生物量均表现出明显降低的趋势 [2] [3] 。高原鼠兔干扰与高寒草甸常见植物叶片养分含量的关系:高原鼠兔干扰与高山嵩草叶片全磷含量、垂穗披碱草和小花草玉梅叶片全氮含量呈显著的正二次函数关系,随高原鼠兔干扰水平增加,高山嵩草叶片全磷含量、垂穗披碱草和小花草玉梅叶片全氮含量先升高后降低 [4] [5] 。对甘肃省天祝藏族自治县抓喜秀龙乡境内不同生境条件下阶地、明坡、山脊、阳坡高寒草甸草原生长季中期、末期的草地植被状况进行调查研究,并进行生物多样性评价,旨在探寻生境因子对草地生产力的影响,为不同生境草地合理利用提供依据。结果表明:相同生境下草地植被在生长季中期和末期物种数量差异不显著,但其群落结构差异显著。植被盖度为阳坡、山脊、阶地、阴坡,阶地与阴坡盖度均显著高于阳坡,月底山脊盖度与阴坡差异显著,地上生物量为山脊、阳坡、阶地、阴坡,其中样地间除月初山脊与阳坡生物量差异不显著外,其余均差异显著。同一时期不同生境条件下植被物种数目、群落物种丰富度指数、物种多样性指数均为阶地、阳坡、山脊、阴坡,各生境间群落均匀度指数变化规律不明显,可能不适用于高寒草地的评价。植被群落相似性系数差异变化表明生境对草地群落组成结构的影响显著 [6] 。以青藏高原黄河源区封育三年的灌丛草地、小嵩草退化草地和黑土滩等3个高寒草地群落为研究对象,利用镜检法,对其土壤种子库的规模、物种组成、空间分布及与地上植被的相关性进行研究。结果表明,高寒草地群落围栏内外土壤种子库物种数目差异不显著,但禾本科与莎草科种子在土壤种子库中所占的比例表现为围栏内 > 围栏外,草地围栏能提高原生植被土壤种子库比例。围栏封育能够提高退化高寒草地土壤种子库数量,对退化草地的恢复有重要作用 [7] [8] [9] 。
2. 材料与方法
2.1. 研究地自然概况
研究样地设在青海省河南县南旗村、秀甲村约100 hm2高寒草地上,该大部分地区海拔在3500 m以上。研究区最高海拔4022 m,最低海拔3391 m,属高寒大陆性气候,平均气温9.2℃~14.6℃,年降水量597.1~615.5 mm。分布主要植物有分布的主要植物有矮嵩草(Humilis kobresia)、金露梅(Jin Lumei)、百里香杜鹃(Prunus thyma)、矮生蒿草(Humilis nanum)、小蒿草(Parvus humilis)、苔草(Carex)、高山嵩草(Alpinis)、针茅(Stipa capillata)、披碱草(Cover with alkali grass)、垂穗披碱草 (Elymus nutans)、早熟禾(Kentucky bluegrass)、无芋翠麦(No taro cui wheat)、洽草(Koeleria cristata)、黄芪(Astragalus membranaceus)、麻花艽(Gentiana straminea)、鹅绒委陵菜(Potentilla anserina)、瑞苓草(Saussurea nigrescens)、马先蒿属(Pedicularis)、黄帚橐吾(Ligularia virgaurea)、雪白委陵菜(Potentilla nivea)、矮火绒草(Leontopodium nanum)、高原毛茛(Ranunculus tanguticus (Maxim Orcz.))、獐牙菜(Swertia bimaculata)、美丽风毛菊(Saussurea supera)、独一味(Lamiophlomis rotata (Benth.) kudo.)、风毛菊(Saussurea japonica (Thunb.) DC.)、尖叶龙胆(Gentiana aristata Maxim)、蒲公英(Taraxacum mongolicum Hand.-Mazz.)、湿生扁蕾(Gentianopsis barbata (Froel.) Ma)、密花香薷(Elsholtzia densa Benth)、堇菜(Viola verecunda)、兰石草(Herb of Tibet Lancea)、金莲花(Trolliuschinensis bunge)、细叶亚菊(Ajania tenuifolia)、西藏微孔草(Microula tibetica)等。
2.2. 样地选择
与2016年8月植被生物量最大时期,在研究地点选择具有代表性的高寒草地,每个样地布置3个样方(1.0 m × 1.0 m)作为三次3次重复,测定样方中出现的每个物种的高度、密度、盖度、地上生物量等植被特征以及土壤有关指标。将试验样地划分为退化、阴坡、阳坡、湿地和平滩共5个生境类型(表1)。
2.3. 群落物种多样性的测定
与植物生长高峰期(2016年8月份)分别在不同生境的高寒草地上布置样方,通过测定植物高度、密度、盖度、地上生物量等来分析高寒草地物种多样性,物种多样性指数的计算公式如下 [10] :

Table 1. Dominant species of alpine grassland in different habitats
表1. 不同生境高寒草地的优势物种
物种多样性指数采用辛普森多样指数;
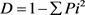 ,
,
式中Pi为第i种的个体数量占总个体数量的比例。
2.4. 数据分析
所有数据采用Excel2003软件进行单因素标准差分析,用Excel2003软件制图。
3. 结果与分析
3.1. 不同生境的高寒草地土壤化学性质
3.1.1. 不同生境高寒草地土壤中的全N和全P2O5的含量情况
阴坡全N含量是9.05 ± 2.76 g∙kg−1且分布相对于其它生境较离散,平滩全N含量是4.58 ± 0.72 g∙kg−1且分布相对于其它生境较均匀,另阴坡、退化、阳坡、湿地和平滩全N含量依次降低;阴坡全P2O5含量是2.41 ± 0.41 g∙kg−1,平滩全P2O5含量是1.55 ± 0.29 g∙kg−1,阴坡、湿地、阳坡、退化和平滩全P2O5含量依次降低,阳坡全P2O5含量分布较均匀,而阴坡相反(图1)。
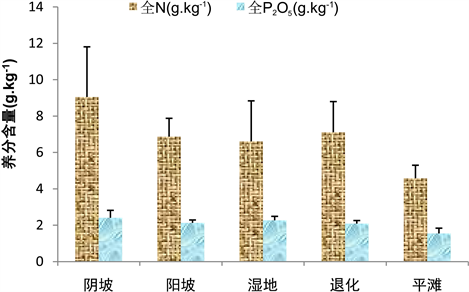
Figure 1. Total N and total P2O5 content in alpine grassland soils from different habitats
图1. 不同生境高寒草地土壤中全N和全P2O5含量情况
3.1.2. 不同生境高寒草地土壤中的全K2O含量情况
阴坡全K2O含量是25.35 ± 2.13 g∙kg−1,平滩全K2O含量是21.17 ± 2.48 g∙kg−1,其阴坡、阳坡、退化、湿平滩全P2O5含量依次降低,退化相对其它生境全K2O含量分布较均匀,而平摊较离散(图2)。

Figure 2. Total K2O content in alpine grassland soils from different habitats
图2. 不同生境高寒草地土壤中全K2O含量情况
3.1.3. 不同生境高寒草地土壤中速效N和速效K的含量情况
阴坡速效N的含量是529.04 ± 119.60 mg∙kg−1,平滩速效N的含量是334 ± 56.85 mg∙kg−1,退化速效N的含量分布较均匀,而湿地速效N的含量分布较离散,其阴坡、阳坡、退化、湿地和平滩速效N含量依次降低;阴坡速效K的含量是430.35 ± 157.04 mg∙kg−1且分布较离散,湿地速效K的含量是233.72 ± 83.40 mg∙kg−1,退化速效K的含量分布较均匀,其阴坡、阳坡、退化、平滩和湿地速效K含量依次降低(图3)。
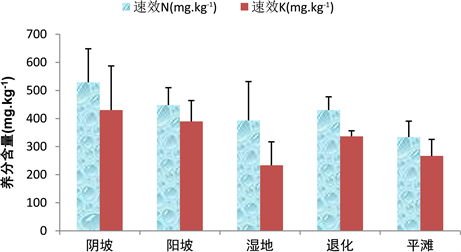
Figure 3. Contents of available N and available K in the soil of alpine grassland in different habitats
图3. 不同生境高寒草地土壤中速效N和速效K含量情况
3.1.4. 不同生境高寒草地土壤中速效P含量情况
阴坡速效P的含量是19.59 ± 10.32 mg∙kg−1,平滩速效P的含量是10.5 ± 3.75 mg∙kg−1,其阴坡、退化、阳坡、湿地和平滩速效P含量依次降低,退化速效P的含量分布相对较均匀,阴坡速效P的含量分布相对较离散(图4)。
3.2. 不同生境下的优势物种分布情况
阴坡以金露梅为优势种,阳坡以早熟禾为优势种,湿地以苔草为优势种,退化毒金露梅和鹅绒委陵菜为优势种,平滩以鹅绒委陵菜为优势种(图5)。
3.3. 不同优势种的海拔分布情况
金露梅群落分布海拔较高平均达3682.47 ± 98.79 m;其次是苔草群落分布海拔达3648.76 ± 141.37 m相对于其它优势种较离散;早熟禾群落分布海拔为3605.8 ± 67.20相对与其它优势种较均匀;而鹅绒委陵菜群落分布的海拔相对较低平均约3582.92 ± 98.84 m (图6)。
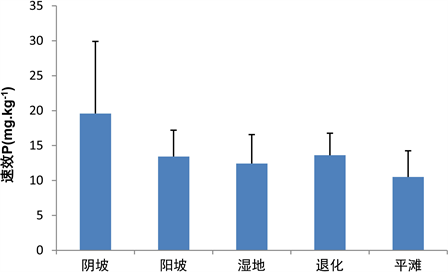
Figure 4. Available P content in alpine grassland in different habitats
图4. 不同生境高寒草地土壤中速效P含量情况

Figure 5. Distribution of dominant species in different habitats
图5. 不同生境下的优势物种分布情况
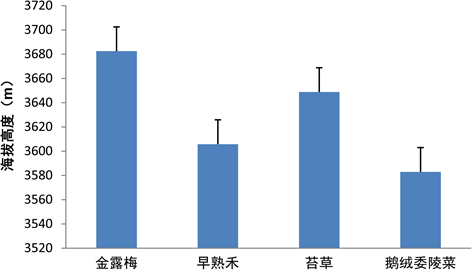
Figure 6. Elevation distribution of different dominant species
图6. 不同优势种的海拔分布情况
3.4. 不同海拔高度与土壤养分的分析
在不同海拔中海拔高度与土壤养分都有着线性关系,根据海拔变化的不同,土壤中全N含量最接近正相关,其关系式是y = 0.019x − 64.82,R2 = 0.857;而随着海拔的变化情况其次是全P2O5和全K2O,然而最不明显的是速效K,其R2 = 0.350 (图7)。
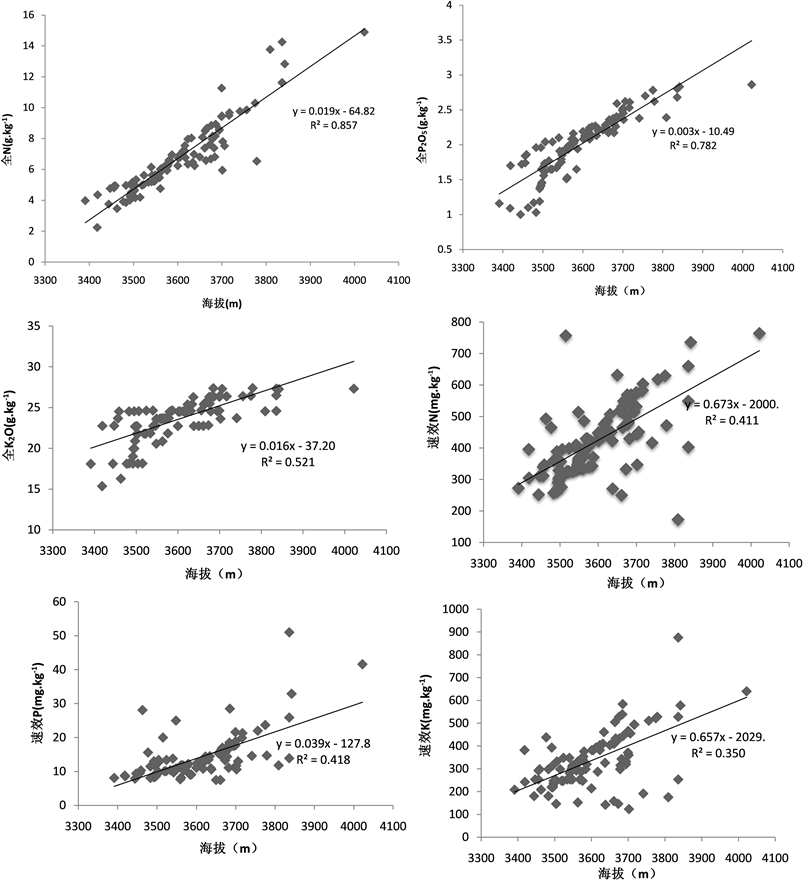
Figure 7. Relationship between different altitudes and soil nutrients
图7. 不同海拔高度与土壤养分关系情况
3.5. 不同植被与土壤养分的分析
3.5.1. 不同植被土壤中全N和P2O5的含量
金露梅群落和苔草群落的土壤全N含量相对较高,其金露梅群落的土壤含全N量为8.75 ± 2.18 g∙kg−1,苔草群落的土壤含全N量为8.35 ± 4.42 g∙kg−1。金露梅群落的土壤全N含量相对与苔草群落分布较均匀;早熟禾群落和鹅绒委陵菜群落的土壤全N含量相对较低,其含量分别为6.47 ± 0.89 g∙kg−1和6.23 ± 2.29 g∙kg−1。早熟禾群落相对与鹅绒委陵菜群落的土壤全N含量分布较均匀。金露梅群落、早熟禾群落、苔草群落和鹅绒委陵菜群落的土壤P2O5含量依次降低,其含量分别是2.3 ± 0.33 g∙kg−1,2.17 ± 0.22 g∙kg−1,2.08 ± 0.54 g∙kg−1和1.97 ± 0.40 g∙kg−1,苔草群落的土壤P2O5分布相对离散,而早熟禾群落的土壤P2O5分布相对均匀(图8)。
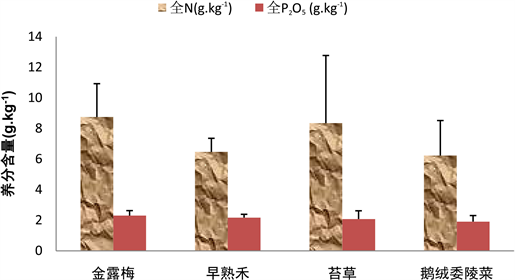
Figure 8. Contents of total N and P2O5 in different vegetation soils
图8. 不同植被土壤中全N和P2O5的含量情况
3.5.2. 不同植被土壤中K2O的含量
早熟禾群落的土壤K2O含量相对较高,早熟禾群落、金露梅群落、鹅绒委陵菜群落和苔草群落的土壤K2O含量依次降低,其土壤K2O含量分别为25.24 ± 1.20 g∙kg−1,24.69 ± 1.35 g∙kg−1,23.09 ± 1.89 g∙kg−1和21.64 ± 3.35 g∙kg−1。早熟禾群落的土壤K2O分布相对较均匀,而苔草群落的土壤K2O分布相对较离散(图9)。
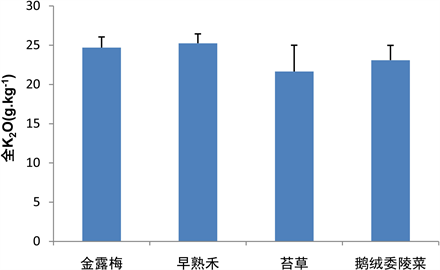
Figure 9. K2O content in different vegetation soils
图9. 不同植被土壤中K2O的含量
3.5.3. 不同植被土壤中速效N和速效K的含量
金露梅群落和早熟禾群落的土壤速效N含量相对较高,其金露梅群落的土壤速效N含量为487.71 ± 74.52 mg∙kg−1,早熟禾群落的土壤速效N含量为453.5 ± 63.81 mg∙kg−1。金露梅群落的土壤速效N含量相对与早熟禾群落分布较离散;苔草群落和鹅绒委陵菜群落的土壤速效N含量相对较低,其含量分别为443.43 ± 178.20 mg∙kg−1和381.42 ± 126.35 mg∙kg−1。鹅绒委陵菜群落相对于苔草群落的土壤速效N含量分布较均匀。早熟禾群落、金露梅群落、鹅绒委陵菜群落和苔草群落的土壤速效K含量依次降低,其含量分别是396.13 ± 120.36 mg∙kg−1,394.14 ± 215.73 mg∙kg−1,305.83 ± 106.23 mg∙kg−1和214.43 ± 58.40 mg∙kg−1,苔草群落的土壤速效K分布相对均匀,而金露梅群落的土壤速效K分布相对离散(图10)。
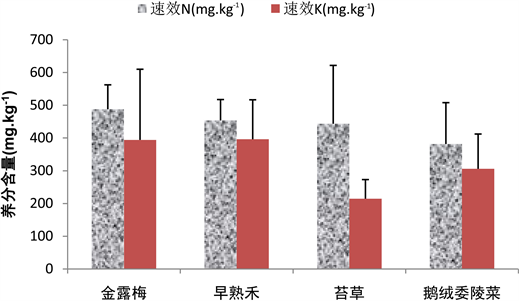
Figure 10. The content of fast-acting N and quick-acting K in different vegetations
图10. 不同植被速效N和速效K的含量
3.5.4. 不同植被土壤中速效P的含量
金露梅群落的土壤速效P含量相对较高,鹅绒委陵菜群落的土壤速效P含量相对较低,金露梅群落、苔草群落、早熟禾群落和鹅绒委陵菜群落的土壤速效P含量依次降低,其土壤速效P含量分别为21.13 ± 13.68 mg∙kg−1,17.71 ± 6.78 mg∙kg−1,15.31 ± 6.04 mg∙kg−1和12.21 ± 6.71 mg∙kg−1。早熟禾群落的土壤速效P分布相对较均匀,而金露梅群落的土壤速效P分布相对较离散(图11)。
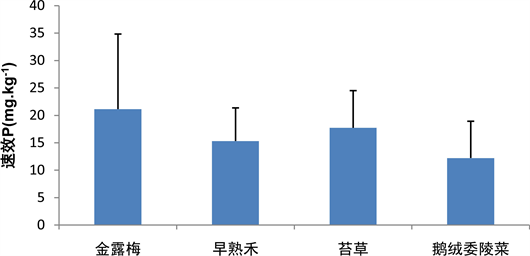
Figure 11. Content of available P in different vegetation soils
图11. 不同植被土壤中速效P的含量
4. 讨论
物种多样性反映了种间生态位的差异,当一个群落包含了更多的生物种类,且每个物种个体数分布比较均匀时,他们之间就容易形成一种复杂的关系。物种多样性具有俩种含义,其一是种的数目或丰富度,只一个群落或生境种物种数目的多寡;其二是种的均匀度,指一个群落或生境中全部物种个体数目的分配状况,它反映的是各物种个体数目分配的均匀程度。多样性指数正是反映丰富度和均匀度的综合指标 [10] [11] [12] 。表1显示在阴坡和退化地金露梅都占优势种,阳坡早熟禾占优势种,湿地苔草占优势种,而平滩鹅绒委陵菜占优势种。阳坡和平滩物种数丰富,退化地物种丰富度较低。
土壤中养分在不同生境(阴坡、阳坡和平滩)中含量是依次降低的(图1~4)。退化和湿地中土壤养分也是递减的(图1~4)。
郝文芳等人的研究结果表明,影响丰富度指数的因子从大到小依次是:土壤含水量、群落盖度、土壤有机质、坡度、坡向和海拔 [13] [14] [15] [16] 。植被盖度、坡度、海拔高度和坡向对多样性产生的影响,主要通过直接影响土壤含水量来实现的,土壤含水量是影响多样性最为关键的生态因子 [17] [18] 。本研究结果显示,全N含量阴坡 > 退化 > 阳坡 > 湿地 > 平滩,全P2O5含量阴坡 > 湿地 > 阳坡 > 退化 > 平滩,全K2O含量阴坡 > 阳坡 > 退化 > 湿地 > 平滩,速效N阴坡 > 退化 > 阳坡 > 湿地 > 平滩,速效P含量阴坡 > 退化 > 阳坡 > 湿地 > 平滩,速效K含量阴坡 > 阳坡 > 退化 > 平滩 > 湿地。其中土壤中养分阴坡在不同生镜中均大于其他不同生境,但阴坡物种数少于阳坡和平滩,所以物种数可能和温度有很大关系,而阴坡主要物种为金露梅分布海拔较高(图6),可以得出金露梅适合生长在高海拔低温的环境中。在不同海拔中海拔高度与土壤养分都有着线性关系,根据海拔变化的不同,土壤中全N含量最接近正相关,其关系式是y = 0.019x − 64.82,R2 = 0.857;而随着海拔的变化情况其次是全P2O5和全K2O,然而最不明显的是速效K,其R2 = 0.350 (图7),海拔在升高,其土壤养分在缓缓增长。金露梅群落和苔草群落的土壤全N含量相对较高,其金露梅群落的土壤含全N量为8.75 ± 2.18 g∙kg−1,苔草群落的土壤含全N量为8.35 ± 4.42 g∙kg−1。金露梅群落的土壤全N含量相对与苔草分布较均匀;早熟禾群落和鹅绒委陵菜群落的土壤全N含量相对较低,其含量分别为6.47 ± 0.89 g∙kg−1和6.23 ± 2.29 g∙kg−1。早熟禾群落相对与鹅绒委陵菜群落的土壤全N含量分布较均匀。金露梅群落、早熟禾群落、苔草群落和鹅绒委陵菜群落的土壤P2O5含量依次降低,其含量分别是2.3 ± 0.33 g∙kg−1,2.17 ± 0.22 g∙kg−1,2.08 ± 0.54 g∙kg−1和1.97 ± 0.40 g∙kg−1,苔草群落的土壤P2O5分布相对离散,而早熟禾群落的土壤P2O5分布相对均匀(图8)。早熟禾群落的土壤K2O含量相对较高,早熟禾群落、金露梅群落、鹅绒委陵菜群落和苔草群落的土壤K2O含量依次降低,其土壤K2O含量分别为25.24 ± 1.20 g∙kg−1,24.69 ± 1.35 g∙kg−1,23.09 ± 1.89 g∙kg−1和21.64 ± 3.35 g∙kg−1。早熟禾群落的土壤K2O分布相对较均匀,而苔草群落的土壤K2O分布相对较离散(图9)。金露梅群落和早熟禾群落的土壤速效N含量相对较高,其金露梅群落的土壤速效N含量为487.71 ± 74.52 mg∙kg−1,早熟禾群落的土壤速效N含量为453.5 ± 63.81 mg∙kg−1。金露梅群落的土壤速效N含量相对与早熟禾分布较离散;苔草群落和鹅绒委陵菜群落的土壤速效N含量相对较低,其含量分别为443.43 ± 178.20 mg∙kg−1和381.42 ± 126.35 mg∙kg−1。鹅绒委陵菜群落相对于苔草群落的土壤速效N含量分布较均匀。早熟禾群落、金露梅群落、鹅绒委陵菜群落和苔草群落的土壤速效K含量依次降低,其含量分别是396.13 ± 120.36 mg∙kg−1,394.14 ± 215.73 mg∙kg−1,305.83 ± 106.23 mg∙kg−1和214.43 ± 58.40 mg∙kg−1,苔草群落的土壤速效K分布相对均匀,而金露梅群落的土壤速效K分布相对离散(图10)。金露梅群落的土壤速效P含量相对较高,鹅绒委陵菜群落的土壤速效P含量相对较低,金露梅群落、苔草群落、早熟禾群落和鹅绒委陵菜群落的土壤速效P含量依次降低,其土壤速效P含量分别为21.13 ± 13.68 mg∙kg−1,17.71 ± 6.78 mg∙kg−1,15.31 ± 6.04 mg∙kg−1和12.21 ± 6.71 mg∙kg−1。早熟禾群落的土壤速效P分布相对较均匀,而金露梅群落的土壤速效P分布相对较离散(图11)。然而很难断定物种多样性的变化与土壤等环境因子变化间的因果关系,这是许多相关研究难以解答的问题 [19] [20] [21] ,因为生态系统过程本身很复杂,其生物与非生物间存在极其复杂的正负反馈机制,这些均是需要进一步研究的课题。
5. 结论
本研究分析了不同生境高寒草地植被的土壤特性和不同植被土壤养分,以及不同生境环境下的物种所生活的海拔高度情况和不同海拔与土壤养分的关系。主要结论如下:阳坡、平滩、阴坡、湿地和退化地物种多样性依次降低,土壤养分含量之间接近正相关,及其和生境的关系是阴坡、阳坡和平滩,土壤中养分含量是依次降低的,退化和湿地中土壤养分也是递减的,其中全N和速效P含量退化相对于阳坡略高。金露梅群落分布海拔较高,其土壤养分也相对较高,而鹅绒委陵菜群落分布海拔较低,其次金露梅群落适合生长在高海拔低温的环境中,随着海拔的上升土壤养分也极微的在增加。
基金项目
青海省创新平台建设专项(2017-ZJ-Y20) Qinghai innovation platform construction project。
文章引用
邓艳芳,肖能洋,杨元武,周华坤. 河南县不同生境高寒草地植被与土壤特性分析
The Analysis of Vegetation and Soil Characteristics of Different Habitats in the Different Habitats of Henan County[J]. 土壤科学, 2019, 07(02): 89-100. https://doi.org/10.12677/HJSS.2019.72011
参考文献
- 1. 王彩虹, 于瑞梅. 天山北坡中段蒿类荒漠草地土壤变化与植被退化关系的分析[J]. 草食家畜, 2009, 144(3): 65-68.
- 2. 陈生云, 赵林, 秦大河, 等. 青藏高原多年冻土区高寒草地生物量与环境因子关系的初步分析[J]. 冰川冻土, 2010, 32(2): 405-413.
- 3. 邵玺文, 韩梅, 韩忠明, 孔伟伟, 杨利民. 不同生境条件下黄芩光合日变化与环境因子的关系[J]. 生态学报, 2009, 29(3): 1470-1477.
- 4. 高三平, 李俊祥, 徐明策, 等. 天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征[J]. 生态学报, 2007, 27(3): 947-952.
- 5. 杨惠敏, 王冬梅. 草-环境系统植物碳氮磷生态化学计量学及其对环境因子的响应研究进展[J]. 草业学报, 2011, 20(2): 244-252.
- 6. 董云龙, 张德里, 陈建纲, 等. 东祁连山高寒草地不同生境条件下植物群落结构特征及多样性比较[J]. 草叶学报, 2014, 22(3): 481-487.
- 7. 李凌浩. 土地利用变化对草原生态系统土壤碳储量的影响[J]. 植物生态学报, 1998, 22(4): 300-302.
- 8. 邓斌, 国华, 刘志云, 等. 封育三年对三种高寒草地群落土壤种子库的影响[J]. 草叶学报, 2012, 21(5): 23-31.
- 9. 熊慧, 马承恩, 李乐, 曾辉, 郭大立. 不同生境条件下蕨类和被子植物的气孔形态特征及其对光强变化的响应植物[J]. 生态学报, 2014, 38(8): 868-877.
- 10. Timan, D., Wedin, D., Knops, J.O., et al. (1996) Productivity and Sustainability Influenced by Biodiversity in Grassland Ecosystems. Nature, 379, 718-720. https://doi.org/10.1038/379718a0
- 11. Jin, H.J., Li, S.X., Wang, S.L., et al. (2000) Impacts of Climatic Change on Permafrost and Cold Regions Environments in China. Acta Geographica Sinica, 55, 161-173.
- 12. 李玉霖, 毛伟, 赵学勇, 等. 北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J]. 环境科学, 2010, 31(8): 1716-1725.
- 13. 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000.
- 14. 乔有明, 王振群, 段中华. 青海湖北岸土地利用方式对土壤碳氮含量的影响[J]. 草业学报, 2009, 18(6): 105-112.
- 15. Zhang, F.W., Han, D.R., Guo, X.W., Li, Y.K. and Cao, G.M. (2011) Response of Potential Carbon Sequestration Capacity to Different Land Use Patterns in Achnatherum Splendens Grassland in Qinghai-Tibetan Plateau. Acta Botanica Boreali-Occidentalia Sinica, 31, 1866-1872.
- 16. Fang, J.Y., Liu, G.H. and Xu, S.L. (1996) Biomass and Net Production of Forest Vegetation in China. Acta Ecologica Sinica, 16, 497-508.
- 17. 庾强. 内蒙古草原植物化学计量生态学研究[D]: [博士学位论文]. 北京: 中国科学院植物研究所, 2009.
- 18. 郝文芳, 杜峰, 陈小燕, 等. 黄土丘陵区天然群落的植物组成、植物多样性及其环境因子的关系[J]. 草地学报, 2012, 20(4): 609-615.
- 19. 欧阳经政, 李以康, 林丽, 等. 不同牧压梯度下高寒草甸演替及植物物种多样性分异特征[J]. 草地学报, 2015, 23(5): 944-950.
- 20. 宋理明, 娄海萍. 青海湖地区天然草地土壤湿度研究[J]. 中国农业气象, 2006, 2(2): 151-155.
- 21. 杨元武, 李希来, 周旭辉, 等. 高寒草甸植物群落退化与土壤环境特征的关系研究[J]. 草地学报, 2016, 24(6): 1211-1217.