Advances in Environmental Protection
Vol.
12
No.
02
(
2022
), Article ID:
50965
,
10
pages
10.12677/AEP.2022.122048
太子城河浮游植物生态化学计量学研究
李子尧,吉力,黄头生*
华北电力大学,工程生态学与非线性科学研究中心,北京
收稿日期:2022年3月18日;录用日期:2022年4月22日;发布日期:2022年4月29日

摘要
本文研究了寒冷地区山区河流浮游植物生态化学计量学特征及元素间平衡关系。在2020年5月、2020年8月、2020年11月、2021年2月采集河北省张家口市崇礼区太子城河四个季度的浮游植物样品,测定碳(C)、氮(N)、磷(P)、硫(S)、氢(H)、铁(Fe)元素含量,分析其生态化学计量学特征及相关性。结果表明,浮游植物C、N、P、S、H、Fe元素含量分别为82.14 ± 32.12 g/kg、9.22 ± 3.5g/kg、1.46 ± 0.55 g/kg、1.96 ± 0.86 g/kg、2.36 ± 1.36 g/kg、12.64 ± 10.57 g/kg。整体上C、N、P元素含量相对稳定,S、H、Fe元素含量波动较大,Fe元素变异系数高达83.62%。太子城河浮游植物元素摩尔组成为C156.00N15.41S1.54H51.17Fe5.10P,与经典Redfield比值C106N16P相比存在差异。C元素占比较高表明太子城河浮游植物对C的需求高且固C能力强,N:P比值与前人研究结果吻合。太子城河浮游植物N:P比值为15.41,表明其生长受N、P共同限制。C、N、P元素含量间显著正相关,S、H、Fe之间无明显相关性;C:P与C:N、N:P间显著正相关,而C:N、N:P间以及H:S、Fe:S、H:Fe间相关性不强,表明浮游植物不同元素间耦联性不同,C、N、P作为浮游植物重要的营养元素相关性强。本文研究为深入理解寒冷山区河流浮游植物的生态化学计量学特征及元素平衡关系提供了基础。
关键词
寒冷地区,太子城河,浮游植物,元素,化学计量学

Ecological Stoichiometry Characteristics of Phytoplankton in Taizicheng River
Ziyao Li, Li Ji, Tousheng Huang*
Research Center for Engineering Ecology and Nonlinear Science, North China Electric Power University, Beijing
Received: Mar. 18th, 2022; accepted: Apr. 22nd, 2022; published: Apr. 29th, 2022

ABSTRACT
This research performed an investigation on the phytoplankton ecological stoichiometry characteristics and the balance relationship between elements in a mountain river of cold region. The samples of phytoplankton of four seasons were collected in May 2020, August 2020, November 2020 and February 2021 from Taizicheng River in Chongli, Zhangjiakou City, Hebei Province, China. The contents of carbon (C), nitrogen (N), phosphorus (P), sulfur (S), hydrogen (H) and iron (Fe) were measured, and their ecological stoichiometry characteristics and correlation were analyzed. The results showed that, the contents of C, N, P, S, H and Fe of phytoplankton were 82.14 ± 32.12 g/kg, 9.22 ± 3.56 g/kg, 1.46 ± 0.55 g/kg, 1.96 ± 0.86 g/kg, 2.36 ± 1.36 g/kg and 12.64 ± 10.57 g/kg, respectively. Generally, the contents of C, N and P were relatively stable, while the contents of S, H and Fe fluctuated greatly, and the variation coefficient of Fe content reached as high as 83.62%. Moore of phytoplankton elements in Taizicheng River was C156.00N15.41S1.54H51.17Fe5.10P, which showed significant difference compared with the classical Redfield ratio C106N16P. The high proportion of element C indicated that phytoplankton in Taizicheng River had a high demand for C and a strong ability to consolidate C, and the N:P ratio was consistent with the results of previous studies. The ratio of N:P was 15.41, suggesting that the growth of phytoplankton in Taizicheng River was restricted by both N and P. The contents of C, N and P were positively correlated, while S, H and Fe had no significant correlation. C:P was significantly positively correlated with C:N and N:P, while C:N, N:P, H:S, Fe:S, and H:Fe were not strongly correlated, indicating that coupling relationship between phytoplankton elements was different and C, N, and P were highly correlated as important phytoplankton nutrient elements. The investigation in this research provided a basis for penetrating the phytoplankton ecological stoichiometry characteristics and elements balance relationship in a mountain river of cold region.
Keywords:Cold Region, Taizichegn River, Phytoplankton, Elements, Stoichiometry

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
浮游植物在水生态系统的生物地球化学循环中起着至关重要的作用,通过吸收C、N、P、S、H、Fe等元素用于生命活动过程 [1] [2],将元素以有机物的形式固定。这些有机物(颗粒或溶解态)一部分被下一营养级摄取吸收再排出,一部分沉淀到河流底部经微生物作用及水体交换进入自然界 [3]。在这个生态系统物质交换和生命活动相互影响 [4] 的过程中,浮游植物体内的多种化学元素平衡会形成一个特有的化学计量比。生态化学计量学是研究生命元素平衡关系和特征的重要手段 [5]。
20世纪初,Alfred Redfield证明了表层浮游生物的元素组成与海洋中溶解营养物大体一致,并确定了106C:16N:1P的Redfield摩尔比 [6]。这引出了一种假设,即海洋浮游植物已经进化到具有改变自生元素水平与其生长水环境相似的能力 [7]。但是,目前已知异养和自养水生生物的C:N:P比值不一定符合Redfield的公认比值。由于营养元素之间的相互作用和生理控制会导致生物产生特有的C:N:P化学计量比,不同环境下的同种浮游植物、相同环境下的不同浮游植物N:P比都不同。根据统计得出浮游植物最佳N:P比在8.2:1至45.0:1之间,而Redfield提出的结论更像是浮游植物平均N:P比 [8]。随着分析测试技术的发展,不仅是水生态系统,全球格局下的各种生态系统的生态化学计量学特征都产生了新的成果 [9]。
为了解寒冷山区河流生物的生态化学计量学特征及元素平衡关系,本研究以河北省张家口市崇礼区太子城河生态系统中的浮游植物为研究对象,通过对C、N、P、S、H、Fe六种元素进行研究,揭示了寒冷山区河流浮游植物多种元素平衡的化学计量学特征。
2. 材料与方法
2.1. 研究区域与野外采样
太子城河位于张家口市崇礼区,处于北纬40˚58'41~40˚51'16和东经115˚27'18~115˚15'23之间,其特殊的海拔高度和地理环境使其气候特征近似寒温带,为寒冷地区淡水山区河流。太子城河发源于太子城东侧老虎沟,向西至马丈子沟分出支流后最终注入清水河,河道曲折,流向东西,垂直落差大,海拔高度在1212 m至1864 m之间,水流速度较快,水动力强,流域年均气温低于3℃,昼夜温差大。
在2020年8月~2021年2月,为研究太子城河浮游植物生态化学计量学特征,沿河设置13个采样点(见图1),1至4号样点为太子城河上游,1号点老虎沟泉眼为整条河流源头,河水较浅,4至6号点为太子城河干流,7至9号点为太子城河下游,10至12号点为太子城河支流。采样位置用GPS定位取得经纬度海拔数据如表1,保证每次采样在同一位置。在2020年5月、2020年8月、2020年11月和2021年2月采集四个季度的河流浮游植物样品,使用13号浮游生物网嵌套25号浮游生物网拖拽法,每个点采集样品500 ml。各采样点地理位置如图1,采样点数据如表1。
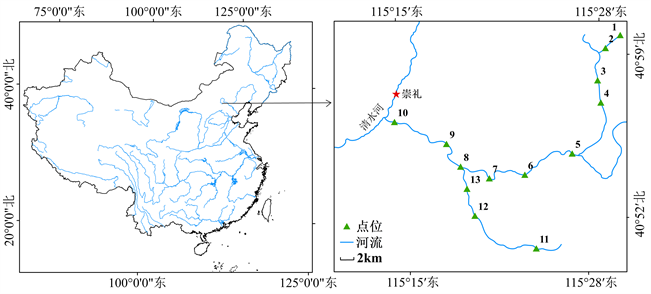
Figure 1. Location of sampling points in Taizicheng River
图1. 太子城河采样点地理位置

Table 1. Latitude and longitude coordinates and altitudes of sample points in Taizicheng River
表1. 太子城河采样点经纬度坐标及海拔高度
2.2. 研究方法
生态化学计量学对于浮游植物元素组成的研究有两种方法,一是直接测定浮游生物生物体和有机碎屑的元素组成,二是根据水体中生源元素的浓度变化进行反推。本文采用第一种方法。将采集的浮游植物样品在体视显微镜下挑拣,与河流中的颗粒悬浮物等杂物分离,将挑出的浮游植物分别装入透明小瓶中,在冷冻干燥机中干燥得到粉末状样品,进行元素测试。碳(C)、氮(N)、氢(H)、硫(S)四种元素使用Elemantar:Vario EL cube型元素分析仪测定;磷(P)、铁(Fe)元素使用安捷伦7800型ICP-MS电感耦合等离子质谱仪测定。
实验数据使用SPSS 25.0进行平均值、标准偏差、变异系数分析及单因素方差分析,对C、N、P、S、H、Fe元素及其化学计量比之间进行双侧Person相关性检验。将数据处理的结果在Excel 2007软件中制作表格,在Origin 8.5软件中绘制箱线图。
3. 结果
3.1. 浮游植物元素含量特征
太子城河浮游植物元素测试结果如表2,浮游植物六种元素含量的变化规律并不相同,C元素在2月份寒冷生长缓慢时期最高、8月份的快速生长时期最低C元素作为“骨架元素”并未受生长速率降低的影响;N元素含量在2020年5月、11月较低,在2020年8月2021年2月较高;P元素含量的分布5月份最低,逐渐升高至11月份,天气转冷,生长速率降低P元素含量也逐渐降低;S元素含量在8月到达最高值后逐渐降低,N、S两种与蛋白质相关的元素在8月份达到峰值,表明此时浮游植物体内蛋白质含量较高;H元素含量在5月和11月存在较高值,与N元素恰好相反;Fe元素含量在5月略低8月和11月随光合作用呼吸作用的加强逐渐升高,但在2月整体急剧下降,整体含量低于其他月份的十分之一。
Table 2. Content characteristics of phytoplankton C, N, P, S, H and Fe in Taizicheng River
表2. 太子城河浮游植物C、N、P、S、H、Fe的含量特征
整体上C、N、P元素含量稳定性较S、H、Fe元素略高。C、N、P因植物生命活动的需求及生长发育中蛋白质合成的原因,整体变化在一个相对较小的范围内,S、H、Fe虽然也是植物生长的必需元素,元素含量却没有C、N、P稳定,尤其Fe元素,在2月随着植物生命活动的减弱,Fe元素大量流失,化学计量占比大幅下降。
3.2. 浮游植物C、N、P化学计量学特征
太子城河浮游植物C、N、P元素的生态化学计量比采用摩尔比值。结果如图2,C:P比值春季平均值190.77,波动范围106~360;夏季平均值136.49,波动范围86~246;秋季133.16波动范围70~191;冬季平均值161.09,波动范围73~308。四个季节C:P比值变异系数分别为0.37、0.33、0.31、0.47。N:P比值时间分布为春季平均值19.23,波动范围7~35;夏季平均值15.83,波动范围6~32;冬季平均值10.22,波动范围3~16;冬季平均值15.04,波动范围6~28。四个季节N:P比值变异系数分别为0.46、0.49、0.33、0.36。C:N比值时间分布为春季平均值10.85,波动范围4~15;夏季平均值9.80,波动范围5~21;秋季平均值14.29,波动范围8~28;冬季平均值10.50,波动范围6~23。四个季节变异系数分别为0.29、0.40、0.39、0.41。太子城河浮游植物C、N、P总体化学计量学组成为C:N:P = 156.00:15.41:1。C:P、N:P比值被认为直接与生长速率相关,C:N比值就相对稳定,指数增长时期的浮游生物需要富集大量P元素以转录RNA,因此自5月份起两个比值均开始降低,到次年2月气温降低逐渐升高。太子城河浮游植物C:P比值高于海洋浮游生物的默认值106,而N:P与海洋浮游生物具有相似性。

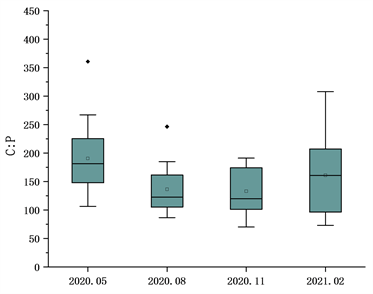
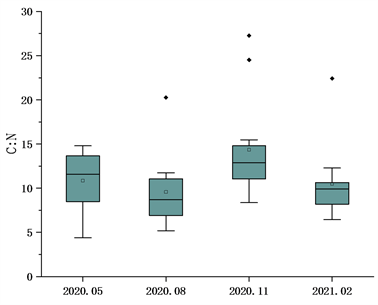
Figure 2. Mole ratio characteristics of phytoplankton C, N and P in Taizicheng River
图2. 太子城河浮游植物C、N、P的摩尔比值特征
对太子城河浮游植物所有样品C、N、P元素含量及C:P、N:P、C:N的比值进行Pearson相关性分析。结果如表3,C元素含量与N、P元素含量之间在0.01级别下显著正相关,而N、P之间仅在0.05级别下显著正相关。C:P比值与C元素含量呈显著正相关,而与P元素含量呈显著负相关;N:P比值与N元素含量呈显著正相关,而与P元素含量呈显著负相关;C:N比值与C元素含量相关性不大而与N元素含量呈显著负相关,一定程度上说明C:N比值的变化与C元素含量变化关系较小,主要取决于N元素含量变化;C:P与N:P的比值之间在0.01级别显著正相关性,与C:N的数值之间在0.05级别显著正相关,而N:P与C:N之间只有微弱的负相关。
Table 3. Pearson correlation coefficients among phytoplankton C, N, P, C:P, N:P, C:N in Taizicheng River
表3. 太子城河浮游植物C、N、P、C:P、N:P、C:N间Pearson相关系数
**在0.01级别(双尾)相关性显著;*在0.05级别(双尾)相关性显著。
3.3. 浮游植物S、H、Fe化学计量学特征
对于S、H、Fe三种元素,采取同样的方法进行处理并对太子城河浮游植物的H:S、Fe:S和H:Fe比值进行研究如图3。H:S春季平均值36.04,波动范围17.12~55.58、夏季平均值28.24,波动范围18.18~43.67;秋季平均值47.31,波动范围26.39~92.42;冬季平均值56.45波动范围32.96~121.82,变异系数分别为0.26、0.34、0.42、0.60。Fe:S比值时间分布为春季平均值3.60,波动范围0.87~12.44、夏季平均值5.06,波动范围1.75~11.74;秋季平均值7.88波动范围4.12~19.78;冬季平均值0.31,波动范围0.07~0.69,变异系数分别为0.79、0.61、0.68、0.89。H:Fe比值时间分布为春季平均值13.56,波动范围4.47~41.19,夏季平均值6.91,波动范围2.53~11.53,秋季平均值7.86,波动范围4.77~13.43、冬季平均值265.58,波动范围70.83~562.02。变异系数分别为0.38、0.45、0.44、0.66。
太子城河浮游植物S、H、Fe总体化学计量学组成为H:Fe:S = 41.47:40.28:1。H:S比值在2020年5月、8月较为稳定且S元素占比有增大趋势,而11月与次年2月由于浮游植物体内蛋白质含量的下降,元素比不稳定且逐渐升高。而Fe:S、H:Fe比值由于Fe元素含量在2月的大幅降低导致变化较大,导致2021年2月份的比值异常。Fe元素与浮游植物光合作用呼吸作用有关,在2月份随光合作用呼吸作用的减弱,其含量降低了95%。在此之前Fe:S比值稳步上升,H:Fe保持稳定,2月份H:Fe比值显著升高,Fe:S比值显著降低。H:S春季平均值36.04,波动范围17.12~55.58、夏季平均值28.24,波动范围18.18~43.67;秋季平均值47.31,波动范围26.39~92.42;冬季平均值56.45波动范围32.96~121.82,变异系数分别为0.26、0.34、0.42、0.60。Fe:S比值时间分布为春季平均值3.60,波动范围0.87~12.44、夏季平均值5.06,波动范围1.75~11.74;秋季平均值7.88波动范围4.12~19.78;冬季平均值0.31,波动范围0.07~0.69,变异系数分别为0.79、0.61、0.68、0.89。H:Fe比值时间分布为春季平均值13.56,波动范围4.47~41.19,夏季平均值6.91,波动范围2.53~11.53,秋季平均值7.86,波动范围4.77~13.43、冬季平均值265.58,波动范围70.83~562.02。变异系数分别为0.38、0.45、0.44、0.66。
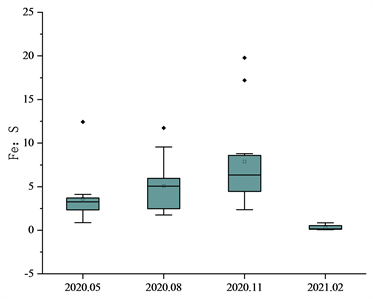

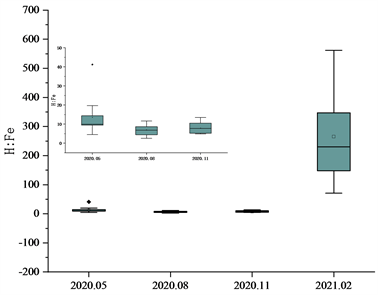
Figure 3. Mole ratio characteristics of phytoplankton S, H and Fe in Taizicheng River
图3. 太子城河浮游植物S、H、Fe的摩尔比值特征
对太子城河浮游植物所有样品的S、H、Fe三种元素含量及H:S、Fe:S、H:Fe比值进行Pearson相关性分析。结果如表4,Fe与S、H之间相关性不显著,而S、H间几乎无相关性。H:S与Fe:S无显著相关,而与H:Fe在0.05级别显著正相关;Fe:S与H:Fe比值在0.01级别显著负相关。
Table 4. Pearson correlation coefficients among phytoplankton S, H, Fe, H:S, Fe:S, H:Fe in Taizicheng River
表4. 太子城河浮游植物S、H、Fe、H:S、Fe:S、H:Fe间Pearson相关系数
**:在0.01级别(双尾)相关性显著;*:在0.05级别(双尾)相关性显著。
4. 讨论
Redfield首次研究了海洋浮游生物的元素组成,提出了C106H263N16P的元素比。根据水体生态系统水文环境地质因素的变化,浮游生物化学计量比和生物组成存在差异 [10]。Anderson [11] 通过实测提出了浮游生物新的元素组成C106H175N16P,Wollast加入了对S元素的研究,得出了C106H263N16S1.7P的分子式,徐德琳 [12] 测得太湖中草型湖区和藻型湖区元素分别为C113.14N20.06P和C105.86N14.75P。Klausmeier计算了浮游植物生长爆发期和相对平稳期的N:P比值,竞争均衡时N:P较高,生长爆发时N:P较低,得出浮游植物最佳N:P比在8.2至45.0之间,并提出了经典的N:P = 16这个比值是不同时期和环境下各个物种的平均比值。本文测得太子城河浮游植物元素组成为C156.00N15.41S1.54H51.17Fe5.10P,对比经典Redfield比例,C元素占比较高,N、P比相近,H占比大幅减小。
太子城河河水深度较浅,光照较强,这可能是导致浮游植物C元素占比较高的原因。Berger [13] 在欧洲中部选择65个湖泊研究浮游生物在不同深度的化学计量学,证明了光照与C:P比值之间的正相关关系。浮游植物群落结构的演替也会导致化学计量学的变化,如硅藻是河流浮游生物C元素组成的主要贡献者,硅藻在浮游植物群落结构中优势度越高,浮游植物C元素的化学计量比越大。硅藻正是太子城河浮游植物群落组成的主要物种之一,也是造成太子城河C元素占比较高的原因。太子城河浮游植物N:P比值为15.41,基本符合经典的Redfield比。在11月和2月P元素含量增大,N元素在2月有所上升,低温会诱导植物储存营养盐用以抵抗冬季对生长带来的不利影响。在化学计量学研究中通常以N:P比值判断限制性元素,当N:P < 14时,N是限制性营养元素,而当N:P > 16时,P则是限制性营养元素,在14至16之间时,受N、P的综合限制。结合实际数据可以得出结论:太子城河浮游植物生长受N、P共同限制。
浮游植物元素之间通过自身的生命活动相互耦合,N主要用于植物生产蛋白质、P主要参与核酸的合成,其他一些元素的储存依靠离子间平衡的有所调整。植物的生长需要合成蛋白质,而合成蛋白质必然伴随DNA转录生成RNA增加P元素的消耗,这个过程显著增强了N、P的相关性;C元素作为“骨架元素”在细胞壁中起支撑作用,随着植物的生长发育含量增加。植物的C、N、P的含量之间都显著相关,而S、H、Fe之间相关性不显著,只有S元素与N元素之间在0.01级别显著相关,两者都参与了蛋白质的合成。但是这并不能证明元素之间不存在相互影响,有研究表明,硅藻中的Fe元素可以通过影响自身生长速率和体型大小影响C:N:P比值 [14],Boyd和Geider [15] 的研究证明藻类中的铁元素可以促进藻类蛋白质和核酸合成过程中的部分酶活性和对大气中CO2的吸收,从而影响自身C、N、P的化学计量学特征。
5. 结论
本文对寒冷山区河流浮游植物C、N、P、S、H、Fe元素的生态化学计量学特征进行了研究,将研究结果与Redfield比值及前人的研究结果进行了对比分析,得出主要结论如下:
1) 太子城河浮游植物元素组成为C156.00N15.41S1.54H51.17Fe5.10P且存在季节性波动,与Redfield比值及前人研究相比,C元素占比较高、H元素占比较低,Fe元素含量变化幅度大,其余元素含量在小范围内波动;
2) 根据化学计量学限制性元素理论,太子城河浮游植物生长受N、P元素共同限制;
3) 太子城河浮游植物C、N、P元素含量及其比值间通过生命活动而耦合,相关性较强,对浮游植物的生长影响大,S、H、Fe元素三者之间则相对独立并无显著相关。
基金项目
国家水体污染控制与治理科技重大专项“冬奥会核心区生态修复与水源涵养功能提升技术与示范”(2017ZX07101002)。
文章引用
李子尧,吉 力,黄头生. 太子城河浮游植物生态化学计量学研究
Ecological Stoichiometry Characteristics of Phytoplankton in Taizicheng River[J]. 环境保护前沿, 2022, 12(02): 350-359. https://doi.org/10.12677/AEP.2022.122048
参考文献
- 1. Moreno, A.R. and Martiny, A.C. (2018) Ecological Stoichiometry of Ocean Plankton. Annual Review of Marine Science, 10, 43-69. https://doi.org/10.1146/annurev-marine-121916-063126
- 2. Urabe, J., Nakanishi, M. and Kawabata, K. (1995) Contribution of Metazoan Plankton to the Cycling of N and P in Lake Biwa. Limnology and Oceanography, 40, 232-242.
- 3. Maklno, W., Coiner, J.B. and Stemer, R.W. (2003) Are Bacteria More like Animals than Plants? Growth Rate and Resource Dependence of Bacterial C:N:P Stoichiometry. Functional Ecology, 17, 121-130. https://doi.org/10.1046/j.1365-2435.2003.00712.x
- 4. Stelzer, R.S. and Lamberti, G.A. (2002) Ecological Stoi-chiometry in Running Waters: Periphyton Chemical Composition and Snail Growth. Ecology, 83, 1039-1051. https://doi.org/10.1890/0012-9658(2002)083[1039:ESIRWP]2.0.CO;2
- 5. Elser, J. (2006) Biological Stoichi-ometry: A Chemical Bridge between Ecosystem Ecology and Evolutionary Biology. American Naturalist, 168, S25-S35. https://doi.org/10.1086/509048
- 6. Redfield, A.C. (1958) The Biological Control of Chemical Factors in the Environment. American Scientist, 46, 205-211.
- 7. Elser, J.J., Elser, M.M., MacKay, N.A., et al. (1988) Zoo-plankton-Mediated Transitions between N and P Limited Algal Growth. Limnology and Oceanography, 33, 1-14. https://doi.org/10.4319/lo.1988.33.1.0001
- 8. Downing, J.A. (1997) Marine Nitrogen: Phosphorus Stoichiometry and the Global N:P Cycle. Biogeochemistry, 37, 237-252. https://doi.org/10.1023/A:1005712322036
- 9. 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索[J]. 植物生态学报, 2005(6): 141-153.
- 10. 王保栋, 陈爱萍, 刘峰. 海洋中Redfield比值的研究[J]. 海洋科学进展, 2003(2): 232-235.
- 11. Anderson, L.A. and Sarmiento, J.L. (1994) Redfield Ratios of Remineralization Determined by Nutrient Data Analysis. Global Biogeochemical Cycles, 8, 65-88. https://doi.org/10.1029/93GB03318
- 12. 徐德琳, 林乃峰, 邹长新, 等. 太湖食物网生态化学计量学特征空间差异[J]. 中国环境科学, 2017, 37(12): 4681-4689.
- 13. Berger, S.A., Diehl, S. and Kunz, T.J. (2006) Light Supply, Plankton Biomass, and Seston Stoichiometry in a Gradient of Lake Mixing Depths. Limnology and Oceanography, 51, 1898-1905. https://doi.org/10.4319/lo.2006.51.4.1898
- 14. 邢伟, 吴昊平, 史俏, 等. 生态化学计量学理论的应用、完善与扩展[J]. 生态科学, 2015, 34(1): 190-197.
- 15. Geider, R.J. and Laroche, J. (1994) The Role of Iron in Phytoplankton Photosynthesis, and the Potential for Iron-Limitation of Primary Productivity in the Sea. Photosynthesis Research, 39, 275-301. https://doi.org/10.1007/BF00014588
NOTES
*通讯作者。