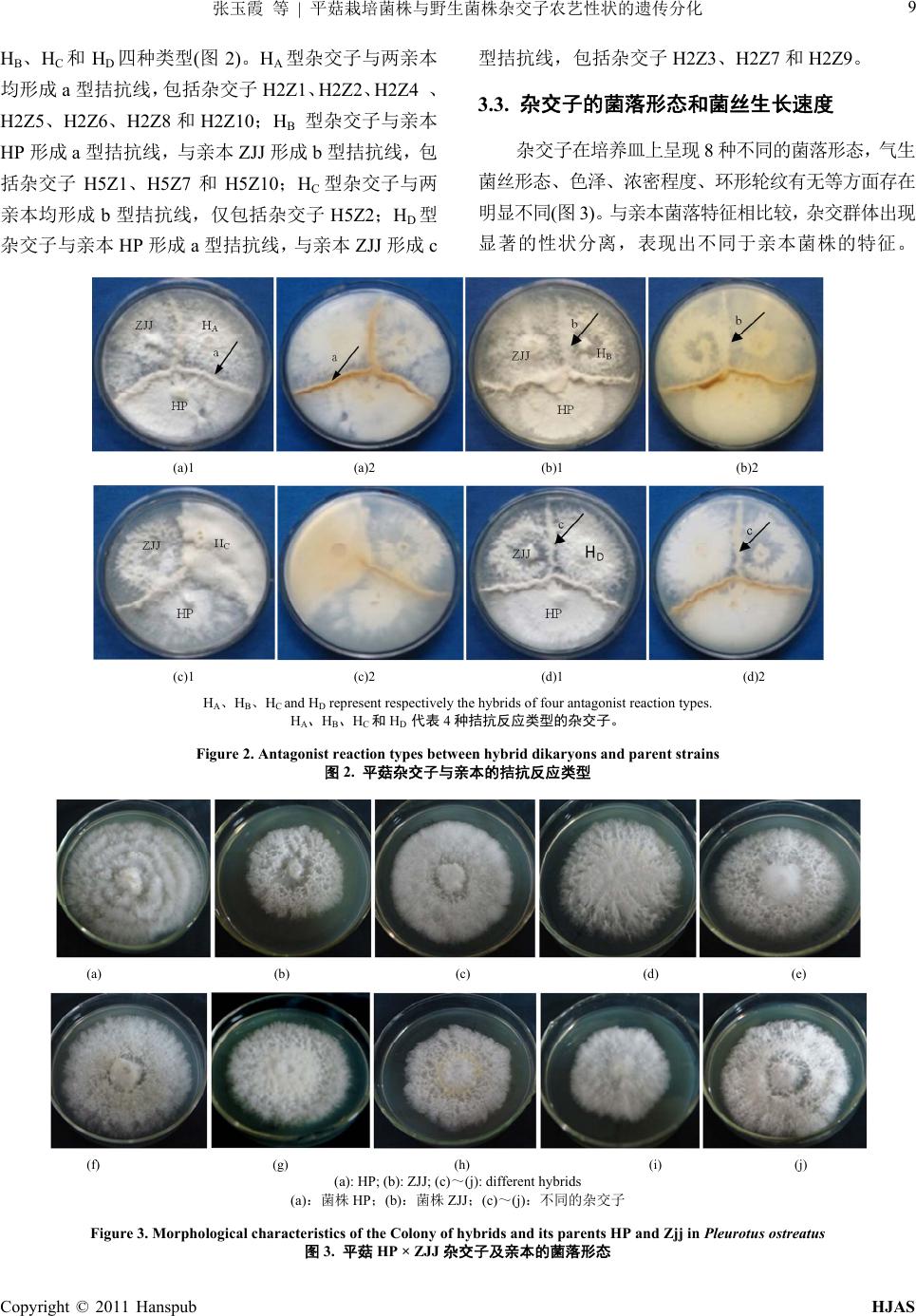
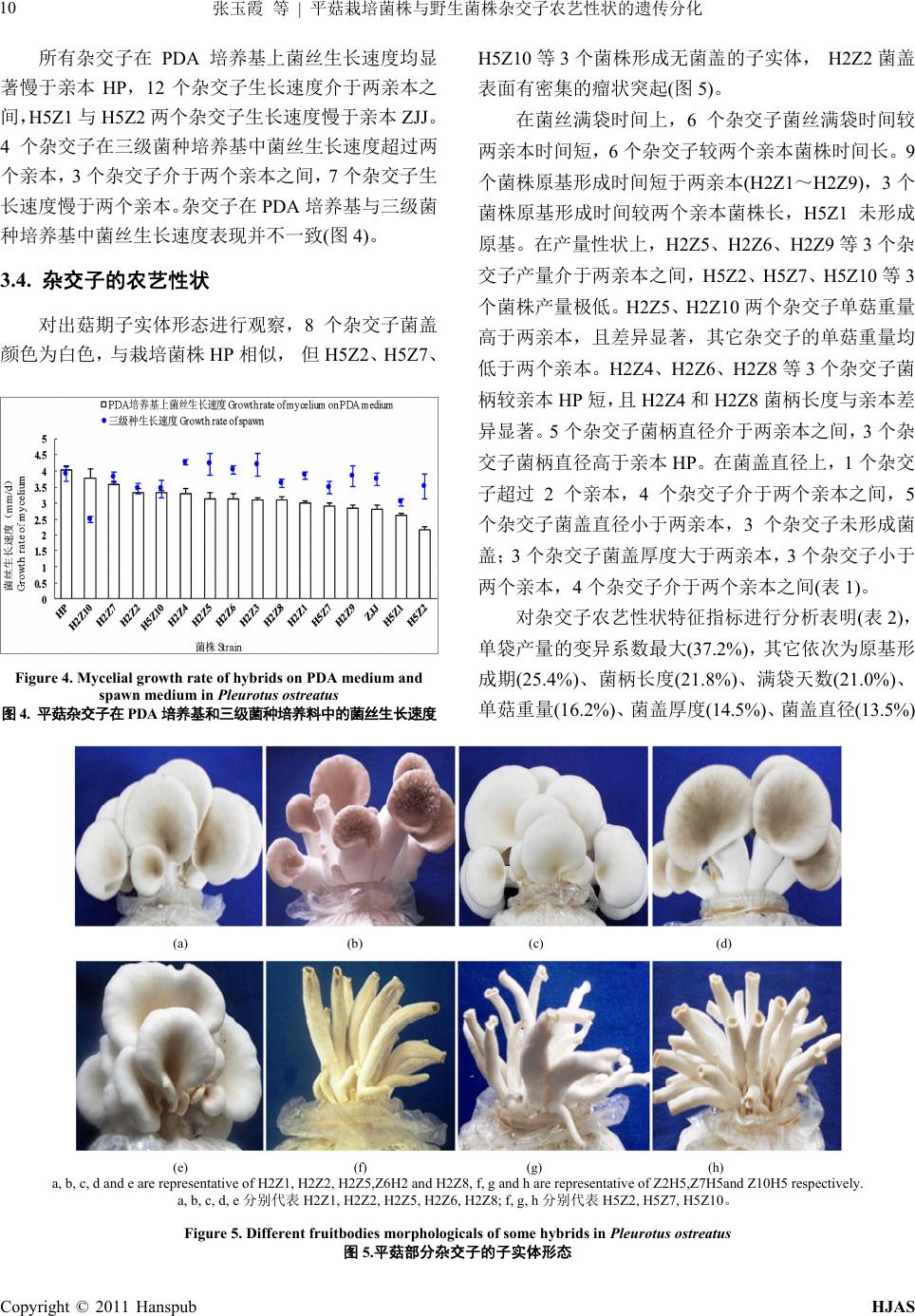
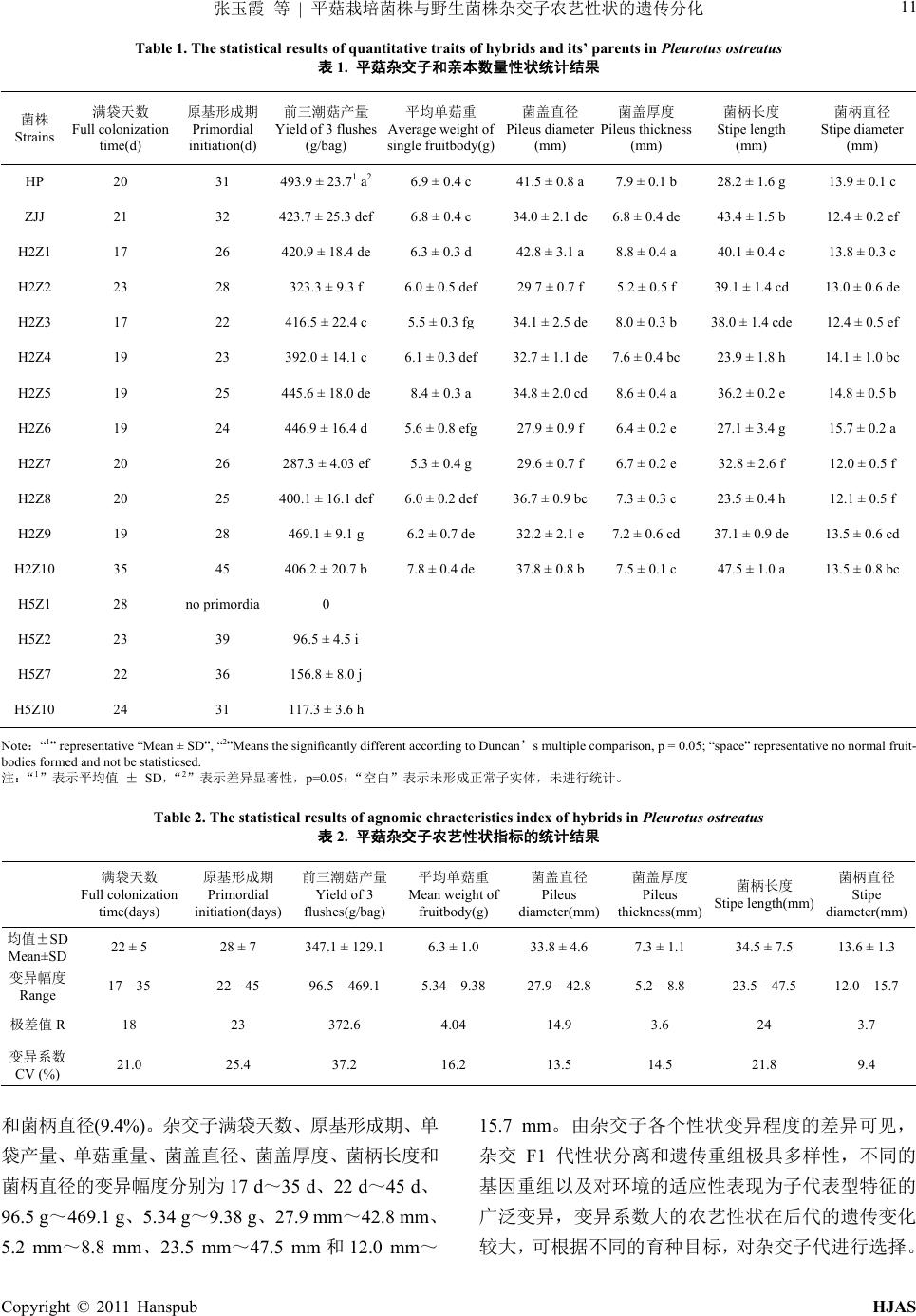
H2Z4 19 23 392.0 ± 14.1 c6.1 ± 0.3 def 32.7 ± 1.1 de7.6 ± 0.4 bc 23.9 ± 1.8 h 14.1 ± 1.0 bc
H2Z5 19 25 445.6 ± 18.0 de8.4 ± 0.3 a 34.8 ± 2.0 cd8.6 ± 0.4 a 36.2 ± 0.2 e 14.8 ± 0.5 b
H2Z6 19 24 446.9 ± 16.4 d5.6 ± 0.8 efg 27.9 ± 0.9 f 6.4 ± 0.2 e 27.1 ± 3.4 g 15.7 ± 0.2 a
H2Z7 20 26 287.3 ± 4.03 ef5.3 ± 0.4 g 29.6 ± 0.7 f 6.7 ± 0.2 e 32.8 ± 2.6 f 12.0 ± 0.5 f
H2Z8 20 25 400.1 ± 16.1 def6.0 ± 0.2 def 36.7 ± 0.9 bc7.3 ± 0.3 c 23.5 ± 0.4 h 12.1 ± 0.5 f
H2Z9 19 28 469.1 ± 9.1 g 6.2 ± 0.7 de 32.2 ± 2.1 e7.2 ± 0.6 cd 37.1 ± 0.9 de 13.5 ± 0.6 cd
H2Z10 35 45 406.2 ± 20.7 b7.8 ± 0.4 de 37.8 ± 0.8 b7.5 ± 0.1 c 47.5 ± 1.0 a 13.5 ± 0.8 bc
H5Z1 28 no primordia 0
H5Z2 23 39 96.5 ± 4.5 i
H5Z7 22 36 156.8 ± 8.0 j
H5Z10 24 31 117.3 ± 3.6 h
Note:“1” representative “Mean ± SD”, “2”Means the significantly different according to Duncan’s multiple comparison, p = 0.05; “space” representative no normal fruit-