Botanical Research
Vol.07 No.03(2018), Article ID:24460,8
pages
10.12677/BR.2018.73033
Dynamic Characteristics of Rhododendron maculiferum Community on North Slope of Shennongjia
Linsen Yang*, Zhiguo Jiang, Yingchun Wang, Huiliang Yu, Jinyuan Yang
Key Lab of Conservation Biology for Shennongjia Golden Monkey, Administration of Shennongjia National Park, Shennongjia Hubei

Received: Mar. 23rd, 2018; accepted: Apr. 16th, 2018; published: Apr. 23rd, 2018

ABSTRACT
The Rhododendron maculiferum community in the Abies fargesii forest belt in Shennongjia was studied via three 400 m2 quadrats. There were 84 plant species of 58 genura and 32 families in three quadrats. Rosaceae is the most in the wood layer with 19 species. The importance value of Fargesia spp. was the highest. Fragaria orientalis, Polygonum pinetorum, Saussurea populifolia were commonly seen in the herb layer. On average, there were 6 species in the tree layer, 15 species in the shrub layer, and 20 species in the herb layer, and the total was 38 species. The most individuals were in 5 -8 cm diameter class with a percentage of 61.3%, and the height of Rhododendron is mainly distributed between 4 - 7 m accounting for 80.9% of the total. Because of the high density, some young individuals died before they can grow up. The ratio of the dead to living standing trees is 7.0%. Most of the dead standing trees of Rhododendron were between the height of 3 - 5.9 m and between the diameter of 3 - 5.9 cm, respectively accounting for 75% and 81.3% of the total number of dead standing trees. With the increase of altitude, population density and tree height of Rhododendron community have decreased and the community levels tend to be simpler. The nanophanerophyte accounted for 31% and hemicryptophyte for 27.4% as the main elements of the community. This kind of life form structure adapted to the local conditions of long cold winter and humid climate. The north temperate type is the most at genus level, accounting for 76.4%. The main plant distribution type is temperate distribution. East Asia type also accounted for an important proportion. Upper Abies fargesii layer disappeared after disafforestation decades ago, so the Rhododendron maculiferum at under layer developed into single-storied stand, and then the species diversity of the tree layer, shrub tree and grass layer decreased, and the average tree height was smaller. As the elevation increased, the biodiversity of Rhododendron shrubs decreased, and the height and diameter also decreased. Rhododendron community at prime period is relatively stable, while the invasion of Fir, Betula utilis in forest or at the edge promotes the succession. The main cause of the forest gap is natural death, high wind, insect pest and animal damage. Gap caused by the dead wood or windthrow is the main reason promoting the regeneration. In general, the evolution tendency is that fir forest would displace rhododendron shrubs.
Keywords:Rhododendron maculiferum, Community Characteristic, Succession, Shennongjia
神农架北坡麻花杜鹃灌丛群落特征
杨林森*,姜治国,汪迎春,余辉亮,杨敬元
神农架国家公园管理局神农架金丝猴保育生物学湖北省重点实验室,湖北 神农架

收稿日期:2018年3月23日;录用日期:2018年4月16日;发布日期:2018年4月23日

摘 要
以3个400 m2样方对神农架亚高山麻花杜鹃(Rhododendron maculiferum)灌丛群落进行研究。群落中有植物84种,58属,32科。木本植物中蔷薇科(Rosaceae)种类最多,有19种。灌木层箭竹(Fargesia)的重要值最大。草本层常见的有东方草莓(Fragaria orientalis)、凉山悬钩子(Rubus fockeanus)、星果草(Asteropyrum peltatum)等。3个样方物种数平均为38种,乔木层、灌木层、草本层的平均物种数分别为6种、15种、20种。杜鹃高度主要分布在在4~7 m之间,占总株数的80.9%。径级构成以5~8 cm最多,占61.3%。枯立木数量与活立木株数的比值为7.0%,杜鹃枯立木占枯立木总株数的86.5%。枯死的杜鹃在高度3~5.9 m及胸径3~5.9 cm为最多,分别占枯立木的75%与81.3%。体现了群落在高度郁闭下的自疏作用。随着海拔的升高,群落层次趋向简单,种群密度降低,高度也趋于变小。矮高位芽植物占31%,地面芽植物占27.4%,是群落的主要成分。这种生活型结构与较长寒冬季节与湿润气候相适应。样地属级水平区系温带成分为76.4%,植物区系温带属性明显,尤其北温带属是主要成分,东亚植物区系占较大比重。人为干扰后上层的巴山冷杉消失,下层杜鹃层发展形成单优麻花杜鹃群落,乔、灌、草三层的物种多样性降低,平均树高变小。随着海拔的升高,杜鹃灌丛的生物多样性降低,高度与直径也变小。盛年期的单纯杜鹃群落相对稳定,但冷杉、糙皮桦(Betula utilis)等乔木在群落中或林缘的侵入推动了杜鹃群落的演替。造成林窗的成因有自然死亡、大风、虫害、兽害等。枯立木与倒木造成的林窗是促进更新作用并推动演替的主要原因,杜鹃群落有被巴山冷杉取代的演化趋势。
关键词 :麻花杜鹃,群落特征,演替,神农架

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
杜鹃属是杜鹃花科中种类最多,特有性极强的大属,广布于欧、亚、北美,主产东亚和东南亚,约960种,我国产542种 [1] [2] 。神农架杜鹃花科植物有9属36种,其中杜鹃属有23种 [3] 。麻花杜鹃(Rhododendron maculiferum)主要分布在神农架1500米以上的沟谷、山坡林下或高山地带开阔山坡。麻花杜鹃是除箭竹(Fargesia)外神农架中高海拔分布面积最广、群落最为集中的典型亚高山灌丛,常与粉红杜鹃(R. oreodoxa var. fargesii)、秀雅杜鹃(R. concinnum)等伴生。杜鹃单纯群落的自然分布一般位于林缘、山脊、山顶。巴山冷杉群落是亚高山区域的地带性植被,人为的干扰形成了较大面积的箭竹群落、杜鹃群落。神农架杜鹃花科植物是巴山冷杉林的重要成分,也是高海拔地带的重要植被类型,海拔2000米以上杜鹃林有较为集中连片的分布 [4] [5] 。冷杉-杜鹃群落是川金丝猴等珍稀动物的主要栖息地 [6] [7] 。
2. 研究区概况
神农架林区位于湖北省西北部,大巴山脉、巫山山脉、武陵山脉于此交汇,是汉水与长江的分水岭,地跨东经109˚56'~110˚58',北纬31˚15'~31˚75',辖区总面积3253平方公里。最高峰神农顶海拔为3106.2 m,最低点海拔398 m,相对高差2708.2 m。神农架属于北亚热带季风气候区,气候有温暖湿润与清凉多雨的特征 [8] 。全年辐射103.7千卡/m2,全年日照时数1858.3 h。年降水量在800~2500 mm之间,年平均降雨量为1584.5 mm。一般从9月底至次年4月底为冰霜期。区内平均年蒸发量500~800 mm,干旱指数为0.50~0.53。全年80%的时间盛行东南风。植物区系体现了亚热带向温带过渡的特性,生物多样性十分丰富,维管植物有3229种,特有性较高,保存了较多的原始种属 [9] [10] 。
3. 研究方法
于2016至2017年在神农架北坡海拔2700 m左右选取麻花杜鹃群落样地3个进行调查,样方大小为20 m × 20 m,记录样地的海拔、坡向、坡度等指标(见表1)。乔木层:调查所有DBH ≥ 3 cm植株的胸径、树高及存活状态。灌木层:记录样方内出现的全部灌木种,对其中的全部灌木分种计数,调查其高度、盖度、多度。草本层:调查样方内所有草本植物的高度、多度和盖度。多度采用Drude多度分级法调查。重要值 = 相对密度 + 相对优势度 + 相对频度 [11] 。植物区系统计采用吴征镒的科属分布区类型划分系统 [12] [13] 。
4. 研究结果
4.1. 群落物种组成
麻花杜鹃群落中有84种植物,58属,32科,其中蔷薇科(Rosaceae)20种,菊科7种,百合科、禾本科均有6种。乔木层有10种,麻花杜鹃为主要建群种,总胸高断面积麻花杜鹃占83.5%,总株数占90.1%,巴山冷杉、光叶陇东海棠也有一定比例(见表2)。灌木层37种,蔷薇科有15种,忍冬科5种,箭竹的重要值最大。草本层43种植物中,最常见的是东方草莓、松林蓼(Polygonum pinetorum)、蕨类、杨叶风毛菊,菊科、百合科、禾本科、毛茛科的植物种类较多。3个样方物种数分别是22、43、49种,平均为38种,乔木层、灌木层、草本层的平均物种数分别为6、15、20种。
大高位芽植物占1.2%,中高位芽占3.6%,小高位芽占14.3%,矮高位芽植物占31.0%,高位芽植物合计占50.0%。而地上芽植物占10.7%,地面芽植物占27.4%。矮高位芽与地面芽植物是群落的主要成分(见图1)。神农架麻花杜鹃群落生境有较长寒冬季节,因而地上芽植物、地面芽植物较多。同时夏季较好的水热条件与湿润气候,为小高位芽、矮高位芽植物及部分乔木营造了较适宜的条件。
4.2. 群落的径级结构与高阶构成
从表3可知,群落高阶在4~7 m之间分布最集中,占总株数的80.9%。径级构成以5~8 cm最多,占

Table 1. The basic characteristics of rhododendron community plots
表1. 杜鹃群落样地基本情况
Table 2. The list of tree layer
表2. 乔木层情况统计表

Figure 1. Life form spectrum of 3 plots
图1. 样地植物生活型谱
61.3%。胸径3 cm以上杜鹃枯立木45株,占枯立木总数的86.5%。杜鹃枯立木与活立木的比例为5.6%。枯死的杜鹃在高度3~5.9 m及胸径3~5.9 cm为最多,分别占枯立木的75%与81.3%,尤其在高阶4 m及径级5 cm死树最多,分别占37.5%与39.6% (见图2)。表现了杜鹃群落在高度郁闭下自疏作用的主要对象是幼树。
三个样地群落径级与高阶表现出较大的差异性。样地1、样地2、样地3的均高分别是6 m、4 m、3 m,平均胸径分别为7 cm、8 cm、5 cm。3样地的总株数分别是416、274、109株。样地1的海拔要低,土层较厚且处于避风的缓坡,因而树高明显高于其他两个样地。样地2处于山包上,风力较大,所以树形矮小但胸径较粗。样地3海拔最高,土层较薄,所以树高与胸径都要小得多,树的密度也要小很多。
Table 3. The structure of DBH class and high order of Rhododendron maculiferum
表3. 杜鹃径级与高阶结构表
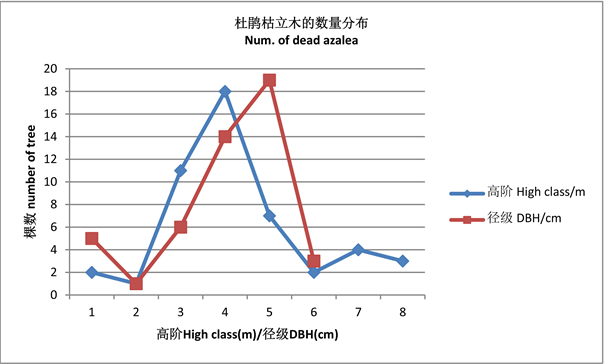
Figure 2. The structure of DBH class and high order of dead tree
图2. 杜鹃枯立木高阶与径级分布
4.3. 植物区系
从科的分布区类型来看,32科中有14科为广布种,占43.8%,温带分布10科,占31.3%,泛热带分布7科,占21.9% (见表4)。在科级水平的植物区系表现了较多的温带属性,与热带区系有较大的关联性。
在属级水平,北温带分布属有29个属,占50.0%,世界广布属14属,占24.1%,东亚分布5属,占8.6%,泛热带分布有4属占6.9%。北温带分布属占总属数占一半,植物区系北温带占主要成分(见表5)。
5. 分析与讨论
5.1. 区系分布
样地属级水平区系温带成分为76.4%,而神农架地区的温带分布属仅占33.8%。随着海拔的升高,世
Table 4. Distribution area type of Family
表4. 科的分布区类型
Table 5. Distribution area type of genura
表5. 属的分布区类型
界广布种变多,热带分布从地区的52.9%下降到4.5%,特有属数量也减少。北温带分布科百合科在样地中有7种,菊科7种中的6个属也属于温带分布,延龄草为较典型的温带分布属。可见,神农架亚高山针叶林带植物区系以温带成分为主,与其他文献的研究结果一致 [14] [15] 。神农架的珍稀植物、中国特有属植物在中海拔地带分布最多,在亚高山针叶林带有星果草属、鬼臼属等中国特有属。东亚分布属有箭竹属等5个属,以东亚为分布中心的温带分布属有黄精属、香青属等5个属,共占所有属数的17.2%,东亚植物区系占较大比重。在中国-日本分布与中国-喜马拉雅分布中,与后者的联系更紧密。神农架植物区系具有丰富性、古老性、特殊性,同时反映了过渡区域的特征 [16] 。
5.2. 径级与高阶
高海拔常绿杜鹃群落的物种多样性相对较低,在中低海拔或人为活动干扰较大的群落多样性增高 [17] 。麻花杜鹃在山顶脊部附近更容易形成单层的林相,山坡上部风力强劲,乔木树种易被风吹倒,较难形成整齐的乔木林。麻花杜鹃冠层紧凑低矮,对雨雪天气有较强抗逆性,因而在神农架顶峰一带的山坡上部形成了大面积的杜鹃灌丛。麻花杜鹃喜阳,也较耐阴,在巴山冷杉林下有较强的适应性。冷杉-杜鹃林是重要的亚高山群落类型。这一群落类型随着海拔的升高而乔木层、灌木层的树高、胸径均呈现下降的趋势,到峰顶趋于灌木化,山坡中下部乔木林下的杜鹃层趋于向小乔木发展,高度可达8~10米,平均胸径约为10 cm。杜鹃灌丛所在的地带性植被主要是巴山冷杉林,受海拔、坡向、坡度、土层厚度、附近植被分布等因素的影响较大,生境异质性造成了杜鹃群落结构差异性。这种差别反映出杜鹃生境异质性及杜鹃对环境的适应性 [18] 。径级与年龄一般呈正相关性。从神农架亚高山杜鹃群落的径级结构数据看,杜鹃种群的年龄结构属于稳定型。
5.3. 群落动态
神农架植被近代以来有着较大的变迁,本地区高海拔区域杜鹃林大多为人工干扰后形成 [5] [16] [19] 。没有人为干扰的高山杜鹃林物种多样性较低。杜鹃形成稠密的群落时,占据的生态位很大,其他物种的生境空间必然受到影响,使物种多样性减少。但从3个样地来看物种丰富度较高,尤其是灌木与草本植物还有较大的多样性。三个样地离公路10~100米,增加了人为干扰的可能性。人为干扰的主要方式是地表枯枝落叶层出现破碎与小型林窗的出现。杜鹃群落较厚的枝层落叶层使大多数种子无法萌发,但地表裸露的斑块为物种的侵入提供了机会。
神农架麻花杜鹃群落主要分布于亚高山寒温性针叶林带。杜鹃群落的重要值明显高于其他物种,属于单优势种群。3个样方均有巴山冷杉的存在,尚属幼龄阶段,可以推知,杜鹃群落向冷杉林的演替过程在数十年后会有明显的变化。但由于山地环境多样化,山峰顶部地带气候的独特性,亚高山杜鹃灌丛在神农架山地始终会有相当的比例,山坡中下部由于适合巴山冷杉、桦木等乔木层植物的生长,因而山坡中下部的杜鹃群落面积会有一定的退化或由上层植被层转变成乔木层的下层植被。巴山冷杉林与杜鹃灌丛在成熟的森林生态系统中处于不同的层次。一方面成熟的冷杉林形成良好的上层植被,林下光照不足,不利于杜鹃的生存;另一方面茂密的杜鹃灌丛也形成了整齐的单层植被,不利于冷杉等其他树种的侵入,两者都具有较强的排他性。植物-土壤的反馈作用通过凋落物的分解和矿化作用机制来实现,对植被的更新有重要影响 [20] 。有人认为杜鹃群落林下土壤中存在的化感物质可能是其天然更新障碍的重要原因之一 [21] 。
杜鹃灌丛演替过程的外界干扰因子主要有山火的影响、林窗的形成等。亨利·威尔逊在100余年前描述了神农架顶峰一带的人为砍伐、林火等现象 [19] 。几十年前的木材采伐活动将公路沿线的巴山冷杉采伐后林下的杜鹃层形成单层的灌丛植被,形成现在大部分杜鹃林。现在神农架顶峰一带的植被至少在100余年来受到较大的人为干扰。林窗的形成可能有人为的原因、动物的原因或风倒木、老死木等原因造成。林窗更新是巴山冷杉群落重要的更新方式 [22] ,杜鹃灌丛恢复为冷杉林的过程也主要受冷杉等高大乔木在杜鹃植被中形成林窗效应来实现。从风倒木及枯立木观察,巴山冷杉从幼苗生长为成熟大树需要60年以上的时间,巴山冷杉的自然寿命一般在200年以上,因而植被的转变演替过程需要较长的时间。亚高山常常具有的大风容易形成风倒木,加快了这一进程。近年来在神农架出现的华山松大小蠹、桦三节叶蜂虫害造成较多的树木枯死。野猪、熊等动物剥裂巴山冷杉树干基部树皮造成了一定数量的巴山冷杉死亡。上世纪90年代开始的箭竹枯死后更新过程也对亚高山地区冷杉与杜鹃植被有必然的影响。这些因素说明杜鹃植被的动态演替过程与巴山冷杉、箭竹等其他植被的演替过程互为影响,加上人为因素、气候变暖等原因的影响,作用过程复杂。要进一步了解群落动态与影响因子的相互作用机理,需要更广泛的多角度研究。在南北坡不同坡向、不同海拔、不同植被层次展开调查,从气候、土壤、生理学、种群遗传学等角度开展研究,会有更进一步的研究结果。神农架拥有北亚热带典型的山地垂直自然带谱,其山地植被突出反映了北亚热带生物群落的生物进化与演替的进程,是众多珍稀濒危物种和中国特有种的重要栖息地,因而开展杜鹃群落的持续监测与研究很有必要 [23] 。
致谢
野外调查、数据整理及写作过程得到同事的支持和帮助,在此一并致谢!
基金项目
本研究得到国家科技支撑计划(神农架金丝猴种质资源保育关键技术研究2013BAD03B00)及湖北省自然科学基金计划项目(神农架珍稀植物群落动态监测研究2015CFA149)资助。
文章引用
杨林森,姜治国,汪迎春,余辉亮,杨敬元. 神农架北坡麻花杜鹃灌丛群落特征
Dynamic Characteristics of Rhododendron maculiferum Community on North Slope of Shennongjia[J]. 植物学研究, 2018, 07(03): 260-267. https://doi.org/10.12677/BR.2018.73033
参考文献
- 1. 方瑞征, 闵天禄. 杜鹃属植物区系研究[M]//路安民. 种子植物科属地理. 北京: 科学出版社, 1999: 299-318.
- 2. 杨汉碧, 方明渊, 方瑞征. 杜鹃花科[M]//傅立国, 陈谭清, 朗楷永, 等. 中国高等植物(第五卷). 青岛: 青岛出版社, 2003: 557-678.
- 3. 廖明尧. 神农架地区自然资源综合调查报告[M]. 北京: 中国林业出版社, 2015: 27-61.
- 4. 朱兆泉, 宋朝枢. 神农架自然保护区科学考察集[M]. 北京: 中国林业出版社, 1999: 94-249.
- 5. 李博, 班继德. 鄂西神农架自然保护区巴山冷杉林的研究[J]. 武汉植物学研究, 1988, 4: 345-356.
- 6. Li, Y.M., Stanford, C.B. and Yang, Y.H. (2002) Winter Feeding Tree Choice in Sichuan Snub-Nosed Monkeys (Rhinopithecus roxellanae) in Shennongjia Nature Reserve, China. International Journal of Primatology, 23, 657-674.
- 7. Li, Y.M. (2006) Seasonal Variation of Diet and Food Availability in a Group of Sichuan Snub-Nosed Monkeys in Shennongjia Nature Reserve, China. American Journal of Primatology, 68, 217-233.
- 8. 湖北省神农架林区地方志编纂委员会. 神农架志[M]. 武汉: 湖北科技出版社, 1996: 1-40.
- 9. 赵常明, 陈伟烈. 神农架植被及其生物多样性基本特征[C]//生物多样性保护与区域可持续发展—第四届全国生物多样性保护与持续利用研讨会论文集: 2000年卷. 北京: 中国林业出版社, 2002: 270-279.
- 10. 应俊生, 陈梦玲. 中国植物地理[M]. 上海: 上海科学技术出版社, 2011: 175-203.
- 11. 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533-548.
- 12. 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991(S4): 1-139.
- 13. 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003, 25(3): 245-257.
- 14. 铁军, 张晶, 彭林鹏, 等. 神农架川金丝猴栖息地植物区系特征及食物资源研究[J]. 广西植物, 2009, 29(6): 736-743.
- 15. 郑重. 神农架维管植物区系初步研究[J]. 武汉植物学研究, 1993, 11(2): 137-148.
- 16. 中国科学院武汉植物研究所. 神农架植物[M]. 武汉: 湖北人民出版社, 1980: 15-33.
- 17. 杨林森. 神农架亚高山杜鹃种群的森林群落特征[J]. 湖北农业科学, 2015, 54(19): 4761-4765 + 4789.
- 18. 唐晓琴. 西藏色季拉山高寒杜鹃群落学研究[D]: [硕士学位论文]. 拉萨: 西藏大学, 2009.
- 19. Wilson, E.H. (1913) A Naturalist in Western China. Vol. 1, 42-58.
- 20. Putten, W.H., Bardgett, R.D., Bever, J.D., et al. (2013) Plant-Soil Feedbacks: The Past, the Present and Future Challenges. Journal of Ecology, 101, 265-276.
- 21. 李朝婵, 钱沉鱼, 全文选, 等. 野生露珠杜鹃林不同分解层的土壤化感潜力[J/OL]. 生态学报, 2018(13): 1-7.
- 22. 符婵娟, 刘艳红, 赵本元. 神农架巴山冷杉群落更新特点及影响因素[J]. 生态学报, 2009, 29(8): 4181-4184.
- 23. 谢宗强, 申国珍, 周友兵, 等. 神农架世界自然遗产地的全球突出普遍价值及其保护[J]. 生物多样性, 2017, 25(5): 490-497.