International Journal of Ecology
Vol.05 No.03(2016), Article ID:18451,10
pages
10.12677/IJE.2016.53007
Preliminary Investigation into Habitat-Point Selection of Rhinolophus luctus during Hibernation
Taifu Huang, Le Peng, Tao Wu, Xiaoyan Gong, Min Wang, Youxiang Zhang, Zhixiao Liu*
College of Biology and Environmental Sciences, Jishou University, Jishou Hunan

Received: Aug. 11th, 2016; accepted: Aug. 28th, 2016; published: Aug. 31st, 2016
Copyright © 2016 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



ABSTRACT
The Woolly horseshoe bat (Rhinolophus luctus), the largest species in Rhinolophidae, is a typical cave-dwelling one in the Xiangxi Tujia and Miao Autonomous Perfecture, Hunan Province, and even in the whole Wuling Mountain ranges also bordered on Chongqing, Guizhou and Hubei Provinces. From January to April of 2016, we made a preliminary field investigation into its habitat-point selection for hibernation in the Karstic caves of the Prefecture, and the results were as follows: Of the 28 investigated caves belonging to 7 counties (except Luxi), there were only 11 ones being of hibernating Woolly horseshoe bat, and totally 20 solitary inverted-roosting individuals documented. During hibernation, most of the horseshoe bat roosted in a 0.5 - 3 m height, with a very low security and special vulnerability to human interference or possible predation from other animals. Their roosting positions were relatively close to the cave entrance, mainly in the range of 20 - 40 m distance from the entrance, where the microhabitats were easily influenced by the outer-cave climatic or weather factors, so the body temperature of these bats lay between a wider range (7˚C - 17˚C) with a positive correlation by the habitat-point temperature. Finally, we inevitably put forward some corresponding population conservation recommendations roughly based on their eco- logical characteristics and endangered status.
Keywords:Rhinolophus luctus, Habitat-Point Selection, Karstic Cave, Hibernation, Wuling Mountain Ranges, Population Conservation

大菊头蝠冬眠期栖点选择的初步调查
黄太福,彭乐,吴涛,龚小燕,王敏,张佑祥,刘志霄*
吉首大学生物资源与环境科学学院,湖南 吉首

收稿日期:2016年8月11日;录用日期:2016年8月28日;发布日期:2016年8月31日

摘 要
大菊头蝠(Rhinolophus luctus)是湘西及武陵山地区体型最大而典型的洞栖型菊头蝠,2016年1~4月对其冬眠期间在喀斯特洞穴中的栖点选择模式进行了初步调查。在湘西州境内所调查的28个洞穴中,仅于11个洞穴中发现并记录了其孤居倒挂的20个个体。冬眠期间,多数大菊头蝠的栖点高度介于0.5~3 m,安全性低,易受到人为干扰或其他动物可能的捕食。栖点位置主要选择在离洞口距离20~40 m的范围内,该洞段的微环境易受到洞外天气或气象因子的影响,其体温变化的幅度也较大,介于7℃~17℃之间,并与该洞段栖点的微环境温度呈正相关。针对该物种的生存现状及生态特征,提出了相应的种群保护建议。
关键词 :大菊头蝠(Rhinolophus luctus),栖点选择,喀斯特洞穴(溶洞),冬眠期,武陵山,种群保护

1. 引言
大菊头蝠(Rhinolophus luctus)是翼手目(Chiroptera)菊头蝠科(Rhinolophidae)中体型最大的种类,其前臂长可达70 mm左右,模式产地虽远在爪哇(Temminck, 1835),却广泛分布于我国的南方(包括台湾省) [1] 。近年,由于全球气候变暖及人类的频繁强度干扰,大菊头蝠的分布区还似乎呈现向北温带地区扩散的趋势 [2] 。已知大菊头蝠有7个亚种,分布于我国境内的属于喜马拉雅亚种(R.l. perniger)、华南亚种(R.l. lanous)和海南亚种(R.l. spurcus),本次调查对象中的大菊头蝠为华南亚种 [3] 。有关大菊头蝠的研究已涉及亚种分化、核型、分子特征、系统发育关系、回声定位声波特征及相关的原理与应用等诸多方面 [4] - [11] ,但对于其栖息地选择模式的研究尚缺乏系统的报道。全球范围内,现已知200种以上的蝙蝠利用洞穴作为它们的冬眠及育仔场所 [12] ,栖居于洞内恒黑且相对稳定的处所可避免对自身不利的洞外环境因子的胁迫,并远离捕食者 [13] ,菊头蝠科正是典型的洞栖型类群。鉴于大菊头蝠在菊头蝠科中的典型性和代表性,以及冬眠在蝙蝠生命周期中的重要作用,本文对其在冬眠洞穴中的栖点选择模式进行了初步的调查与研究。
2. 调查区域与方法
2.1. 调查区域简况
湘西土家族苗族自治州(以下简称湘西州) (109˚10'~110˚22.5'E, 27˚44.5'~29˚38'N)位于湖南省的西部,地处酉水河的中游和武陵山脉的东部。武陵山脉地处云贵高原的东缘,属于我国第二阶梯向第三阶梯的过渡地带,由西南向东北斜向延伸,东南面地势较低,西北面地势较高。地貌的总体轮廓是一个以山原山地为主,兼有丘陵和小平原,并向西北突出的弧形山脉。行政区划上,湘西州包括吉首市、龙山县、永顺县、花垣县、保靖县、凤凰县、古丈县、泸溪县,与重庆市、贵州省和湖北省交界。州内生物资源丰富,具有许多孑遗植物及珍稀濒危动物 [14] 。境内石灰岩分布广泛,岩溶发育充分,多溶洞和伏流,属亚热带向暖温带过渡的季风性湿润气候,适合于蝙蝠,特别是洞栖型蝙蝠种群的栖息及繁衍。
2.2. 调查方法
2.2.1. 样洞选择
结合地图、文献及前期工作,随机访问当地群众,初步掌握所调查的区域内溶洞分布的基本情况,然后在向导的带领或指引下找到溶洞所在的位置,并对洞口周围的情况进行观察记录,之后进洞调查,直至人体很难再深入洞穴为止。调查范围涉及湘西州境内除泸溪县(因溶洞少而没有前往调查)以外的其他所有县市。根据调查结果,将溶洞简略地划分为四种类型:
I类洞:洞穴环境不适合(如洞道浅小,洞穴内的气象因子与外界相似,或虽洞道较深但洞内的气温较高、湿度较低),或者洞内旅游活动(如灯光照射、人流频繁)或洞穴周边资源开发利用(如采石场机器昼夜轰鸣、粉尘污染严重)等人为干扰强度很大,菊头蝠不在洞内栖居冬眠。
II类洞:很少或几乎无人为干扰,但洞道相对简单,仅发现个别菊头蝠栖居。
III类洞:人为干扰强度中等(偶尔有人进入洞内),但环境因子比较适合于菊头蝠栖息,有少数菊头蝠栖挂冬眠。
IV类洞:总体上,洞道结构较为复杂,洞内的环境因子比较适合于菊头蝠冬眠,菊头蝠的数量相对较多。
2.2.2. 野外鉴别
主要参考王应祥(2003)《中国哺乳动物种和亚种分类名录与分布大全》所记载,结合张佑祥(2008)、张婵(2013)等论文中描述的特征及《中国哺乳动物图鉴》(盛和林2005)进行蝙蝠物种鉴别、分布区核查及大菊头蝠鉴定。在野外调查期间,大菊头蝠较易辨别,其个体大,前臂长通常介于66~75 mm,马蹄叶发达,覆盖鼻吻部,两侧不具小附叶,鼻孔内外缘突起,并衍生成杯状的鼻间叶;鞍状叶基部向两侧扩展成翼状,使鞍状叶呈三叶形;连接叶先端低而圆,始于鞍状叶后面甚下方;顶叶狭长,呈舌状;翼膜黑褐色,不甚延长,起始于趾部上缘 [1] - [3] 。大菊头蝠不仅体型大,具有独特的鼻吻部构造,而且其栖息行为也很特别,几乎总是单只栖居,倒挂在位置较低的岩洞侧壁,并且翼膜常紧紧地包裹着身体,仅留背部长条形缝隙,易于识别,为本研究带来了方便。
2.2.3. 研究方法
使用高性能的智能手机GPS详细记录洞穴的经纬度和海拔高度,利用激光测距仪UT391+ (量程60 m,精确度±1.5 mm)测量洞穴的洞道长度、蝙蝠离地面的高度等数据,使用标智GM700 (测量范围−50℃~700℃,精确度±1.5℃)手持式激光测温仪测量蝙蝠的体温以及蝙蝠栖挂点的洞壁温度(栖点温度)。测温时,一般将测温仪置于离蝙蝠2 cm处(不能触碰到蝙蝠以免其受到刺激后惊醒),分别在翼膜、背毛、鼻及耳区测量其体温,通常在距离蝙蝠栖挂点5~15 cm的地方测定栖点温度(在该距离范围内的栖点温度不存在显著性差异P > 0.05)。
根据洞道内的人类活动痕迹、洞口植被情况,以及通过与向导和周边群众的交流访谈情况分析确定目前人类活动对洞穴的干扰强度。进洞调查期间,用强光灯扫视洞内人眼视力能够到达之处,观察记录蝙蝠的种类、悬挂方式、栖点特征,特别留意其独处或集群的方式,以及栖点处是否有滴水及其与附近水源的距离等可能影响蝙蝠冬眠的生态因子,同时观察评价蝙蝠在栖点冬眠的安全性。安全性评价分为高、中、低三级:若蝙蝠栖点离地面高,需借助从洞外带进洞内长度为1.5米以上的长树枝、长竹杆、伸缩杆或用石头投掷才可能伤害蝙蝠,或者栖点的隐蔽性很强,很难发现时,评定为“安全性高”;若栖点离地面较高,进洞之人可用1米左右的短木棍、短树枝或用石块能较为容易地伤害到蝙蝠时,评定为“安全性中”;若人可直接用手触碰到蝙蝠时,评定为“安全性低”。
采用Excel、Spss22.0等软件对数据进行统计处理。
3. 结果与分析
3.1. 大菊头蝠的分布与数量
2016年1~4月间,在湘西州除泸溪县以外的所有县市调查了28个溶洞,但仅在吉首市、古丈县、永顺县、龙山县的11个溶洞内发现了大菊头蝠,总共记录到20个孤居冬眠的个体(表1)。
在湘西州境内,大菊头蝠分布于海拔约420~1220 m的范围内(表1),平均海拔高度约为758 m,所栖溶洞多处于人为干扰相对较少的深山,II类洞占64%,在IV类洞中均有大菊头蝠栖居,但IV类洞数量很少,仅占所调查洞穴的10%。大菊头蝠均为单只倒挂栖息,且每个洞穴中的数量很少,约82%的洞穴仅发现1只,但在永顺县境内的湖南小溪国家级自然保护区七里溪的黄泥洞记录到4只,而在大坪岗金鸡洞还记录到了7只,该洞海拔较高(约920 m),有三个洞口,洞口相距15~30 m,洞道具有明显的层级,洞道宽窄高低变化多样,洞壁凹凸、皱褶及岩层纹缝繁多,洞道结构非常复杂,人为干扰也相对较小。调查期间,我们未发现大菊头蝠群居冬眠的现象。
3.2. 大菊头蝠的栖点特征
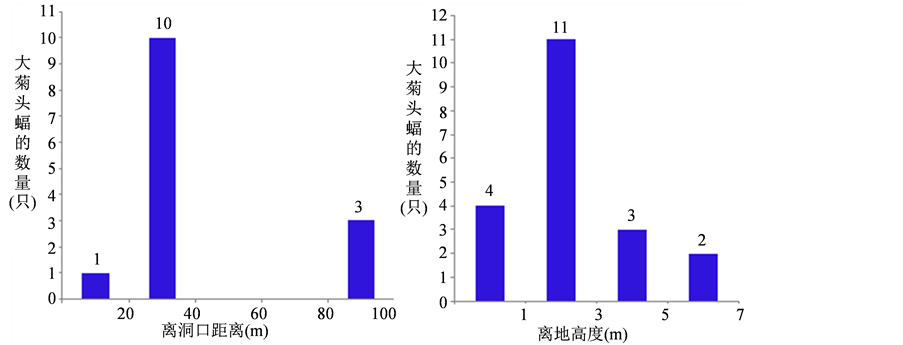
大菊头蝠冬眠期的栖点多选在洞道避风处、离洞口近、离地面低的位置,离洞口的平均距离为38 m (7~87 m, SD = 26.93)。以孤居式栖挂于离洞口较近的洞道内,主要集中于离洞口20~40 m的洞道区域,占总数(仅记录了14个个体,图1(a))的71%。冬眠期间,大菊头蝠栖点离地面的平均高度为2.24 m (0.73~6.69 m, SD = 1.73),集中栖挂于0.5~3 m高度的洞穴侧壁,占总观察数的80% (图1(b))。
从表2可见,大菊头蝠选择在洞壁凹窝处栖息的占35%,洞侧壁占40%,洞顶壁占20%。调查期间,也记录到一例选择特殊栖点冬眠的大菊头蝠,该蝠栖挂于洞道地面岩石堆成的狭小空间内,位置低矮却十分隐蔽。
多数大菊头蝠冬眠期间呈现双足垂直倒挂的姿势,但也有例外,记录到两例仅用左足悬挂的姿态(表2)。虽然其栖点周围无滴水,但选择栖息的洞穴中的空气湿度一般较高(通常在70%以上),洞内的其他区域常有滴水或积水。综合分析和评价大菊头蝠冬眠期的安全性,结果显示“安全性低”的个体占65%,“安全性高”和“安全性中”的分别为20%和15%。
3.3. 大菊头蝠的体温及栖点温度
对大菊头蝠冬眠期间身体四个部位(翼膜、背毛、耳、鼻)的温度进行测量(表3),数据经统计分析,所测量的四个部位的温度并不存在显著性的差异(P > 0.05),并且这4个部位温度的变化也不存在规律性。冬眠期间,大菊头蝠的体温介于7℃~17℃之间,与栖点温度也不存在显著性的差异,但体温与栖点温度的平均值分别为11.26℃ ± 2.4336℃ (n = 14)和10.62℃ ± 2.0517℃ (n = 14),并且呈现显著的正相关,体温与栖点温度之间的关系式为:Y = 1.12X − 0.6318 (Y-体温,X-栖点温度,X∈[7,15]) (图2)。
4. 讨论
洞穴是蝙蝠重要的栖息场所,在气温稳定、湿度较大、黑暗的洞穴环境中,蝙蝠能更好地减少能量与水分的丧失,保持其最基本的生理状态与安全性,因此蝙蝠多选择在洞穴内越冬 [12] [15] 。菊头蝠是典

Table 1. Basic information of Karstic cave investigated in the Xiangxi Tujia and Miao Autonomous Region
表1. 在湘西州境内所调查溶洞的基本情况
Continued
注:干扰强度:强——经常有人进洞或洞口附近存在持续的强干扰源;中——较少有人进入洞内或周围有弱干扰源;轻——偶尔有人进洞;无——没有人进洞。洞道长度:“?”——受限于洞内坍塌、悬崖、深水潭、地下河流等因素影响未能探索整个洞道。
 (a) (b)
(a) (b)
Figure 1. The distance of habitat-point for Rhinolophus luctus away from cave entrance (a) and the height away from cave ground (b)
图1. 大菊头蝠冬眠期栖点离洞口的距离(a)及离地面的高度(b)
Table 2. Habitat characteristics and roosting habit of Rhinolophus luctus during hibernation
表2. 大菊头蝠冬眠期的栖点特征
注:悬挂方式:B——双足挂,L——左足挂。栖点位置或形状:① 洞顶壁,② 洞侧壁,③ 洞壁狭缝处,④ 洞壁凹窝处,⑤ 其他。孤居性:“+”指单独栖息,不集群。栖点滴水情况:“-”指栖点处无滴水。
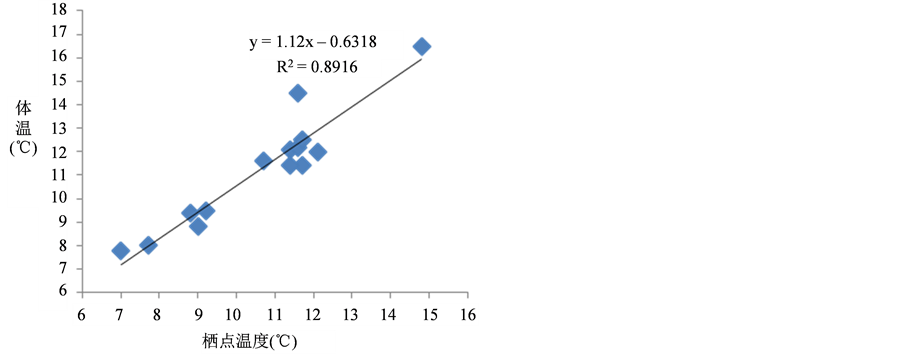
Figure 2. Linear correlation between body and hibernaculum temperature of Rhinolophus luctus in the Karstic cave
图2. 冬眠期间,大菊头蝠的体温与栖点温度之间的线性关系
Table 3. Body and hibernaculum temperature of Rhinolophus luctus (˚C)
表3. 大菊头蝠身体不同部位的体温及栖点温度(℃)
注:翼膜、背毛、耳、鼻为大菊头蝠冬眠期体温的测量部位。体温 = (翼膜 + 背毛 + 耳 + 鼻)温度/4。“-”指无法测量或者测量误差太大。
型的洞栖型蝙蝠,常栖居于洞穴的深处,但大菊头蝠冬眠期的栖点(在洞穴中实际栖挂的具体位置)多位于离洞口不远处。
调查发现,大菊头蝠多见于洞道的前段,离洞口100米以上的深洞道内未见其分布。洞穴深处的气温通常稳定在16℃左右,但洞道前段通过洞口与洞外相通,气温、湿度等气象因子波动较大。测量数据显示,大菊头蝠冬眠期的体温介于7℃~17℃之间,意味着大菊头蝠冬眠期间能适应较宽的环境温度变化,这很可能是其分布范围较广的重要原因之一 [2] [16] 。与主要在洞的深处冬眠的菲菊头蝠(Rhinolophus pusillus)、中华菊头蝠(Rhinolophus sinicus)等同属种类相比,大菊头蝠明显处于深度冬眠状态,冬眠期间对周围环境的异常变化不太敏感,不容易因人类干扰而惊醒,这可能与其受外界气温的影响较大,体温降得更低,代谢率更低,“睡得更死”而不易苏醒有关,而生理上还可能与过氧化物酶体增殖物α受体相关 [17] 。
虽然洞穴内部的小气候或微气候是影响蝙蝠栖点选择的重要因素 [18] ,但洞穴内部的物理因子等因素也不容忽视。调查发现,许多菊头蝠选择在洞顶壁或侧壁位置较高的石钟乳丛生的凹陷或凹窝处冬眠,这种微环境隐蔽性较好,不容易被看见或接触到。但大菊头蝠较为特殊,栖点的位置普遍较低,隐蔽性较差,因而冬眠期个体的安全性一般也较低,这或许也是其种群数量较少的原因之一。我们发现,与大菊头蝠栖息洞段相同的西南鼠耳蝠(Myotis altarium)常栖挂或匍匐于位置较高的洞侧壁或藏于钟乳石形成的狭缝间,隐蔽性、安全性均较高,不易被发现(除非有观察经验的人员,不然很难发现其藏身之处)或受到伤害。与西南鼠耳蝠相比,大菊头蝠体型大,可能因为重力方面的原因而不适于在高处悬挂,也无法挤进狭窄的岩缝间,并且西南鼠耳蝠不仅个体小,而且明显比菊头蝠具有较强的在岩缝间爬行的能力。总之,大菊头蝠栖点安全性“低”使它可能受到更多的环境胁迫与伤害。
在海南五指山一带,大菊头蝠分布于海拔400 m以下 [19] ,而本研究发现其冬眠洞穴的海拔高度多在400 m以上,这种差异是区域性地质地理环境差异的结果,不同的种群生活在不同的区域,总会面临着不同的生态环境条件,地势、气候及食物等条件都会影响蝙蝠种群的生存和繁衍,但蝙蝠的分布是综合因素共同作用的结果 [20] 。因此,对于洞内外的环境因子及其变化规律(包括水、气体成分、生物区系等)仍需作系统深入的数据采集与分析研究,相关工作仍在进行之中。
当前全球的生物多样性在持续下降 [21] ,而人们对翼手目动物的了解仍十分有限,其生存现状令人担忧。虽然目前大菊头蝠已被列为IUCN (2012)低危/需予以关注的物种,以及《中国物种红色名录》(蒋志刚等,2015)近危种(NT),但鉴于大菊头蝠的受胁现状及其冬眠栖点选择的特殊性,我们认为可从以下几个方面对其实施具体的保护行动:1) 将其列入国家二级重点保护野生动物名录,从法律法规上对其进行严格的保护,禁止捕杀、捕捉及伤害;2) 在洞口处张贴通告或建立宣传栏、告示牌等,宣讲有关蝙蝠(尤其是菊头蝠科)的生态知识及种群保护意义,充分调动群众保护蝙蝠的积极性和主动性;3) 在蝙蝠种类及种群数量较多并易遭受人为侵害的洞穴的洞口安置只能让蝙蝠进出而人无法通过的横栏式铁门等设施以避免人们进洞干扰。
致谢
调查期间,得到梁承远、李中炎、李永顺、刘德玉、刘德成等当地群众及本院蔡石坚老师的支持和帮助,谨此一并致以衷心地感谢。
基金项目
国家自然科学基金项目“菊头蝠科对溶洞的择居模式及对岩洞区域生态的指示作用”(项目编号:31560130)暨吉首大学&高望界–小溪国家级自然保护区校企合作省级示范性基地学生研究性学习与创新性调研及实验计划项目“高望界–小溪国家级自然保护区蝙蝠资源调查”。
文章引用
黄太福,彭 乐,吴 涛,龚小燕,王 敏,张佑祥,刘志霄. 大菊头蝠冬眠期栖点选择的初步调查
Preliminary Investigation into Habitat-Point Selection of Rhinolophus luctus during Hibernation[J]. 世界生态学, 2016, 05(03): 57-66. http://dx.doi.org/10.12677/IJE.2016.53007
参考文献 (References)
- 1. 王应祥. 中国哺乳动物种和亚种分类名录与分布大全[M]. 北京: 中国林业出版社, 2003: 34-35.
- 2. 张婵, 王艳梅, 牛红星. 河南省栾川县伏牛山发现翼手目物种大菊头蝠[J]. 动物学杂志, 2013(4): 650-654.
- 3. 张佑祥, 刘志霄, 胡开良, 等. 大菊头蝠在湖南省分布新纪录[J]. 动物学杂志, 2008(2): 141-144.
- 4. 朱斌良. 海南岛翼手目(Chiroptera)物种多样性初步调查与保护对策研究[D]: [硕士学位论文]. 海南: 海南师范大学, 2008.
- 5. 王福勋, 张智伟, 武鹏飞. 大菊头蝠对耳屏下沟的声学功能[C]//中国声学学会物理声学分会. 2009年度全国物理声学会议论文集: 2009年卷. 2009: 2.
- 6. 王福勋. 基于蝙蝠耳的仿生麦克风轮廓模型的数值分析[D]: [博士学位论文]. 济南: 山东大学, 2010.
- 7. 袁小爱, 田东, 谷晓明. 基于核基因RAG1部分序列探讨菊头蝠科和蹄蝠科的系统发育关系[J]. 四川动物, 2012(2): 191-196.
- 8. 袁小爱, 田东, 谷晓明. 基于核基因RAG2部分序列探索菊头蝠总科、蝙蝠科和鞘尾蝠科物种的系统发育关系[J]. 生命科学研究, 2012(3): 215-222.
- 9. 李德伟, 尹锋, 曾玉, 张园, 张信文. 海南岛翼手类地理分布格局的聚类分析[J]. 生物学杂志, 2010(2): 16-20.
- 10. Ellerman, J.R. and Morrison-Scott, T.C.S. (1951) Checklist of Palaeavctic and Indian Mammals. Brit Mus (Nat Hist), London, 121-122.
- 11. Marianne, V., et al. (2015) Surprising Genetic Diversity in Rhinolophus luctus (Chiroptera: Rhinolophidae) from Peninsular Malaysia: Description of a New Species Based on Genetic and Morphological Characters. Acta Chiropterologica, 17, 1-20. http://dx.doi.org/10.3161/15081109ACC2015.17.1.001
- 12. Romero, A. (2009) Cave Biology: Life in Darkness (Ecology, Biodiversity and Conservation). Cambridge University Press, USA, 291. http://dx.doi.org/10.1017/cbo9780511596841
- 13. Culver, D.C. and Pipan, T. (2009) The Biology of Caves and Other Subterranean Habitats. Oxford University Press, USA, 300.
- 14. 金旻. 湘西自治州林业建设与山区发展协调性研究[D]: [博士学位论文]. 北京: 北京林业大学, 2013.
- 15. 张成菊, 吴毅. 洞穴型蝙蝠的栖息环境选择、生态作用及保护[J]. 生物学通报, 2006, 41(5): 4-6.
- 16. Brown, C.R. and Bernard, R.T.F. (1994) Thermal Preference of Schreiber’s Long-Fingered (Miniopterus schreiberisii) and Cape Horseshoe (Rhinolophus capensis) Bats. Comparative Biochemistry and Physiology, 107A, 439-449. http://dx.doi.org/10.1016/0300-9629(94)90023-x
- 17. Han, Y.J., et al. (2015) Adaptation of Peroxisome Proliferator-Activated Receptor Alpha to Hibernation in Bats. BMC Evolutionary Biology, 15, 88. http://dx.doi.org/10.1186/s12862-015-0373-6
- 18. Neuweiler, G. (2000) The Biology of Bats. Oxford University Press, New York.
- 19. 余文华, 范凤兰, 江海声, 等. 海南五指山洞穴蝙蝠对夏季栖息地的选择[J]. 广东林业科技, 2015(5): 74-79.
- 20. 周江, 杨天友. 贵州省松桃县东部地区翼手目物种多样性[J]. 动物学杂志, 2010(2): 52-59.
- 21. Newbold, T., et al. (2016) Has Land Use Pushed Terrestrial Biodiversity beyond the Planetary Boundary? A Global Assessment. Science, 353, 288-291. http://dx.doi.org/10.1126/science.aaf2201
*通讯作者。