Botanical Research
Vol.06 No.03(2017), Article ID:20645,10
pages
10.12677/BR.2017.63016
Analysis of Esterase Isozymograms and Peroxidase Isozymograms on Two Kinds of Pods of Triquetrum-Pod Rapeseed
Yantao Zhu, Xin Zhang, Shengxue Gou, Yajun Zhao, Wenjie Chen, Bin Luo
Hybrid Rapeseed Research Center of Shaanxi Province, Rapeseed Branch of Shaanxi Province, National Oil-Crop Improvement Center, Yangling Shaanxi

Received: May 3rd, 2017; accepted: May 22nd, 2017; published: May 25th, 2017

ABSTRACT
To study the differences of Esterase (EST) and Peroxidase (POD) isozymeograms between triquetrum-siliques (tri-loculus) and ordinary siliques (bi-loculus) of triquetrum-silique materials in Bras- sica napus L., the enzyme spectrums of young siliques of length of 1 cm, 3 cm and 5 cm from 3 selfing populations were analyzed by the polyacrylamide vertical board gel electrophoresis in flowering and silique stage of rapeseed. The results show that: 1) There were no obvious differences in EST isozyme patterns between tri-loculus siliques and bi-loculus siliques in the same developmental stages for the same materials, but there were certain differences in enzyme activity for individual isozymes. In POD isozyme patterns, tri-loculus siliques and bi-loculus siliques were also similar, but there were certain differences in enzyme activity for a few isozymes. 2) The patterns of EST isozymeograms had no significant difference to the siliques in 3 developmental stages for the same materials, the patterns of POD isozymeograms had also no significant difference, but there were some differences in enzyme activity for them. 3) On EST and POD isozymeograms, the smaller the siliques were, the stronger the enzyme activity would be, i.e., the enzyme activity showed a decreasing trend along with the growing and developing of the siliques. 4) Isozyme spectrums of EST isozymeograms of 3 materials in contrast, 14ds67 and 14ds72 had high similarity, and the patterns were consistent, but 14ds78 had greatly difference with the former two. So 14ds67 and 14ds72 had close genetic relationship, and 14ds78 was far away in genetic relationship with the former two. However, isozyme spectrums of POD isozymeograms of 3 materials had no obvious difference, and the enzyme patterns with good consistency, just a few enzyme patterns had certain differences in enzyme activity. 5) The number of bands of POD isozymeograms was significantly less than that of EST isozymeograms for the same materials, and the bands moved fastest were highest in enzyme activity in the enzyme spectrums of POD isozymeograms. Therefore, there are strong consistency in EST isozyme zymogram and POD isoenzyme zymogram, respectively, between tri-loculus siliques and bi-loculus siliques in the same developmental stages for the same materials.
Keywords:Brassica napus L., Bi-Loculus Siliques, Tri-Loculus Siliques, Esterase, Peroxidase, Polyacrylamide Gel Electrophoresis
三棱角油菜两类角果的酯酶同工酶 和过氧化物酶同工酶比较分析
朱彦涛,张新,苟升学,赵亚军,陈文杰,罗斌
国家油料作物改良中心陕西油菜分中心,陕西省杂交油菜研究中心,陕西 杨凌
收稿日期:2017年5月3日;录用日期:2017年5月22日;发布日期:2017年5月25日

摘 要
本文以3个甘蓝型三棱角油菜为材料,在油菜花角期对各材料长度分别为1 cm、3 cm以及5 cm三个不同发育阶段的幼嫩三棱角果(三室)和普通角果(两室)的酯酶(EST)和过氧化物酶(POD)同工酶进行聚丙烯酰胺垂直板凝胶电泳和酶谱的比较分析,以研究三室角果与两室角果在EST和POD同工酶水平上的异同性。研究结果表明:1) 同一材料相同发育阶段三室角果和两室角果的EST同工酶,其带型表现一致,无明显差异,但个别同工酶的酶活性有一定的差异;同一材料相同发育阶段两者的POD同工酶,其带型也表现一致,只是少数同工酶的酶活性有一定的差异。2) 同一材料在三个不同发育阶段的三室角果或两室角果的EST同工酶带型无明显差异,POD同工酶带型也无明显差异,但酶活性存在一定的差异。3) 对EST同工酶和POD同工酶而言,角果越小,酶活性越强,即随着角果的生长发育其酶活性表现为减弱的趋势。4) 3个材料的EST同工酶酶谱比较而言,14ds67与14ds72的酶谱相似程度较高,带型一致,而14ds78的酶谱与前两者的差异较大,说明三棱角材料14ds67和14ds72的亲缘关系较近,而14ds78与前两者的亲缘关系较远。然而,3个材料的POD同工酶酶谱相比较无明显差异,酶带的一致性较好,只是少数条带的酶活性有一定的差异。5) POD同工酶酶谱的酶带数目明显少于相应材料的EST同工酶酶谱的酶带数目,而且迁移快的条带为强带。因此,三室角果与两室角果分别在EST同工酶酶谱和POD同工酶酶谱水平上表现出较强的一致性。
关键词 :甘蓝型油菜,两室角果,三室角果,酯酶,过氧化物酶,聚丙烯酰胺凝胶电泳

Copyright © 2017 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
油菜属于十字花科(Cruciferae)芸苔属(Brassica)植物,是主要的食用植物油来源,也是重要的工业原料和可再生能源。在油菜遗传育种中,资源的多样性是选育油菜新品种的物质基础。
近年来,多室油菜的研究日益受到人们的重视 [1] - [14] 。相对于普通的两室角果油菜而言,多室油菜是指角果为三室或多于三室的油菜,是具有多室多粒性状的一类特殊的油菜种质资源。三棱角(三室)油菜是笔者近年来发现并自主选育的一种新的甘蓝型油菜种质资源,是多室油菜的一种重要类型,具有多室、多粒和抗裂角性强的特性 [8] [13] [14] 。其角果呈三棱柱状、三室,角果皮为三瓣,角果横截面为近三角形。由于三棱角油菜的三室角果比两室角果有较多的角粒数,且三棱角角果由于独特的三棱柱状结构因而具有较强的抗裂角特性,因此三棱角油菜的选育在油菜高产育种以及机械化收获方面具有独特的优势。三棱角油菜的研究和开发利用,对丰富油菜花器官的发育和角果的形态建成理论、探讨三棱角油菜的遗传特性和分子机理,以及对丰富油菜种质资源和油菜高产育种具有重要意义。为了拓宽油菜有限的育种资源,为三棱角油菜材料的有效利用提供理论依据,进一步选育三棱角油菜新品种,探索一条多室油菜高产育种的新途径,有必要对三棱角油菜进行系统和深入的研究。
同工酶研究在植物分类学、遗传学以及作物种子纯度鉴定方面具有重要作用。长期以来,关于油菜作物酯酶(EST)同工酶和过氧化物酶(POD)同工酶的研究已有较多报道,如王保仁等 [15] 、刘忠松等 [16] 、陈松等 [17] 、伊岚等 [18] 以及林宝刚 [19] 研究认为油菜不育系和保持系之间的EST、POD同工酶谱带分别存在差异。于澄宇等 [20] 以及朱彦涛等 [21] 研究认为不育系与保持系的同工酶既受同核的控制,又受异质的影响,质核互作不可忽视。
总之,有关油菜同工酶的研究,以及有关多室油菜的研究,近年来已经取得了一定的进展,但有关多室油菜EST和POD同工酶的研究方面还未见报道。
基于此目的,本文以3个甘蓝型三棱角油菜特异种质为材料,对其不同发育阶段三室角果和两室角果的EST和POD同工酶进行聚丙烯酰胺凝胶电泳和带型的比较分析,旨在为油菜三棱角材料三室角果和三两角果的比较研究提供生理生化方面的证据,这对进一步探索油菜多室性状的遗传和分子机理,利用多室油菜为我国的油菜科研和生产服务有重要意义。
2. 材料和方法
2.1. 材料
3个甘蓝型三棱角(三室)油菜高代自交材料14ds67、14ds72、14ds78,均来源于陕西省杂交油菜研究中心(其中14ds67株高107.50 cm,三室角果率41.79%,低芥高硫,来源于早期收集的油菜三室角果07Zt1的种子;14ds72株高107.48 cm,三室角果率46.64%,低芥高硫,来源于早期收集的油菜三室角果07Zt6的种子;14ds78株高120.85 cm,长角果,三室角果率84.93%,低芥,来源于早期收集的油菜三室角果07Zt25的种子)。2014年秋季播种于本中心油菜试验田,并按照常规方法进行田间管理。次年油菜花角期,取3个材料不同发育阶段的幼嫩3室角果与2室角果,用于进行同工酶聚丙烯酰胺垂直板凝胶电泳,并对电泳结果进行比较分析。
2.2. 方法
2.2.1. EST同工酶的提取和电泳检测
聚丙烯酰胺凝胶电泳参照朱彦涛等(2009) [21] 的方法。油菜花角期,在田间选取各材料典型的无病虫害植株,在植株的主花序和上部一次分枝花序上选取长度分别为1 cm、3 cm以及5 cm的三个不同发育时期的3室角果和2室角果,将所取角果放入冰壶带回室内。先用自来水将角果冲洗干净,再用蒸馏水漂洗后晾干角果表面的水分。每样品称重0.4 g,按W(g)/V(ml) = 1:2的比例加提取液,在冰浴上研磨成浆。然后11,000 r/min离心15 min,取上清液放入冰箱保存。分离胶浓度为7%~8%,浓缩胶浓度为3.6%。每个胶板点样孔为12泳道,点样一个材料的6个样品,每样品按50 μl的点样量重复点样2个泳道。每材料至少做2个胶板的重复,各泳道从左到右依次进行点样,点样顺序如图中所示。点样完毕,在4℃冰柜中进行电泳,电泳采取稳流方式,浓缩胶电流25 mA,分离胶电流45 mA,电泳过程约4~5 h。EST同工酶采用α-醋酸萘酯 + β-醋酸萘酯+固蓝B盐溶液染色。染色完毕后,将胶板漂洗干净,对酶谱进行观察、比较和记录,并对胶板进行照相。
2.2.2. POD同工酶的提取和电泳检测
POD同工酶的提取和电泳检测方法基本与EST同工酶一致,但有以下变化:每样品按50 μl的点样量各点样一泳道,电泳过程约5~6 h,POD同工酶采用愈创木酚——联苯胺染色。
以上研究采用的同工酶电泳和分析方法,是以溴粉蓝作为前沿指示剂,以酶带迁移率的相对大小来进行定性描述的。
3. 结果与分析
3.1. EST同工酶电泳结果和酶谱分析
为了研究和描述方便,可按照EST同工酶酶谱中各条带迁移率的大小和染色深浅程度的不同将酶谱分为快、中、慢三个区域,分别命名为:I) 快速区:条带迁移率大;II) 中速区:迁移率中等、染色深的区域,位于酶谱的中间部位,酶带数量多,至少有4~5条强带,此区域一般是显示各材料特征酶带的主要区域;III) 慢速区:迁移率小、染色较深的区域。
3.1.1. 三棱角材料14ds67三室与两室角果的EST同工酶比较
三棱角材料14ds67的EST同工酶电泳结果如图1所示,由图1可知:在角果长度分别为1 cm、3 cm以及5 cm的三个发育阶段,3室角果与2室角果的EST同工酶带型一致,酶带数目相同,只是个别酶带的酶活性有一定的差异。如在I区,角果长度为1 cm时,3室角果比2室角果对应条带的酶活性要强。同样的,在图谱相同区域,角果长度为5 cm时,2室角果比3室角果对应条带的酶活性要强。其余区域酶带活性无显著差异。
另外,由图1还可知:长度分别为1 cm、3 cm以及5 cm的三个发育阶段的2室角果,其带型之间无显著差异,3室角果也是如此。但长度为1 cm的2室角果和3室角果,其酶活性明显较长度3 cm和5 cm角果的酶活性强。
3.1.2. 三棱角材料14ds72三室与两室角果的EST同工酶比较
三棱角材料14ds72的EST同工酶电泳结果如图2所示,由图2可知:在长度分别为1 cm、3 cm以及5 cm的三个发育阶段的角果中,3室角果与2室角果的EST同工酶的带型基本一致,条带数目相同,只是少数酶带的酶活性存在一定的差异。如对长度为1 cm的角果而言,3室角果比2室角果对应条带的染色要浅,说明3室角果比2室角果对应条带的酶活性要弱。又如在Ⅲ区中,对长度为5 cm的角果其迁移率最小的一条带而言,3室角果比2室角果对应条带的染色要深,说明3室角果比2室角果对应条带的酶活性要强。
另外,由图2还可知:长度分别为1 cm、3 cm以及5 cm的三个发育时期的2室角果,其带型之间无明显差异。同样地,长度分别为1 cm、3 cm以及5 cm的三个发育时期的3室角果,其带型之间也无明显差异。但是,长度为1 cm的2室角果和3室角果,其酶带染色深,说明其酶活性较长度3 cm和5 cm角果的酶活性强。
3.1.3. 三棱角材料14ds78三室与两室角果的EST同工酶比较
三棱角材料14ds78的EST同工酶电泳结果如图3所示,由图3可知:在长度分别为1 cm、3 cm以及5 cm的三个发育时期的角果中,3室角果与2室角果的同工酶带型一致,酶带数目相同,只是部分条带的染色深浅有一定的差异,即3室角果比2室角果酶带的染色程度要深,说明3室角果条带的酶活性比2室角果要强。比如,对长度为1 cm的角果而言,3室角果比2室角果对应条带的染色要深,说明3

Figure 1. Electrophoresis of EST isozymograms on 14ds67; 2R: 2 rooms; 3R: 3 rooms; Sample volumes: 50 μl, the same as follow
图1. 14ds67的EST同工酶电泳结果;2R:2室;3R:3室;点样量:50 μl。下同
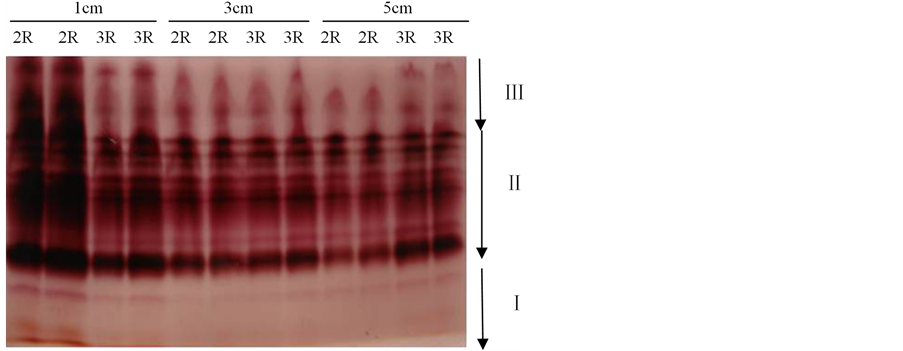
Figure 2. Electrophoresis of EST isozymograms on 14ds72
图2. 14ds72的EST同工酶电泳结果
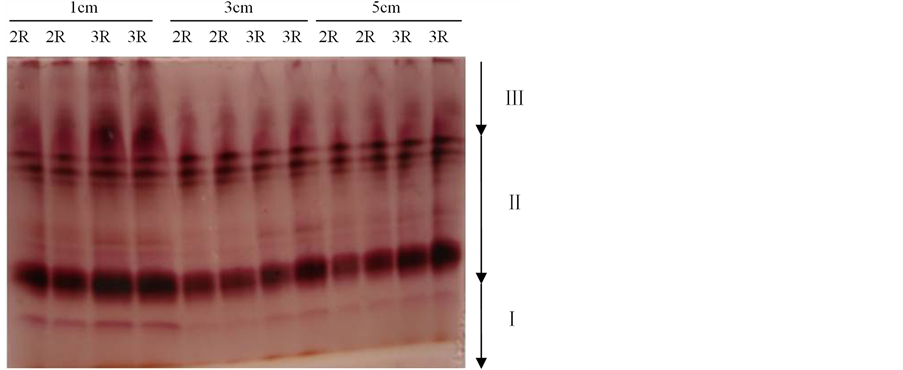
Figure 3. Electrophoresis of EST isozymograms on 14ds78
图3. 14ds78的EST同工酶电泳结果
室角果比2室角果对应条带的酶活性要强。
另外,由图3还可知:长度分别为1 cm、3 cm以及5 cm的三个发育时期的2室角果,其带型之间无明显差异。同样地,长度为1 cm、3 cm以及5 cm的3室角果,其带型之间也无明显差异。但是,长度为1 cm的2室角果和3室角果,其相应酶带染色较深,说明长度为1 cm的角果的酶活性比长度3 cm和5 cm角果的酶活性强。
3.1.4. 3个三棱角材料之间的EST同工酶酶谱比较
由图1~图3可知,14ds67、14ds72和14ds78这3个三棱角材料各样品的特征酶带主要集中在II区。比较来说,14ds67与14ds72的EST同工酶酶谱相似程度高,其带型一致,只是个别酶带的酶活性强弱有一定的差异,说明这两个材料的亲缘关系较近。然而,从带型和条带的染色程度来看,14ds78的EST同工酶酶谱与14ds67和14ds72这两个材料的EST同工酶酶谱差异较大,特别表现在第II区(中速区)中,其酶带数目明显减少,部分酶带染色程度明显变浅,说明14ds78与14ds67和14ds72这两个材料的亲缘关系较远。
3.2. POD同工酶电泳结果和酶谱分析
在三棱角材料的POD同工酶酶谱中,按迁移率大小和染色程度不同可将酶带至下而上依次分为:1条迁移率大的强带、至少2条迁移率中等且相近的较强带、至少1条迁移率小的弱带。
3.2.1. 三棱角材料14ds67三室与两室角果的POD同工酶比较
三棱角材料14ds67的POD同工酶电泳结果如图4所示,由图4可知:在长度分别为1 cm、3 cm以及5 cm的三个发育时期的角果中,3室角果与2室角果的POD同工酶带型基本一致,条带数目基本相同,只是少数条带的染色深浅有一定的差异,说明这些条带的酶活性有一定的差异。如对长度为1 cm的角果而言,3室角果比2室角果对应的条带染色深,说明3室角果比2室角果对应的条带酶活性强;对长度为5 cm的角果而言,其3室角果比2室角果对应的条带染色浅,说明3室角果比2室角果对应的条带活性弱。
另外,由图4还可知:长度分别为1 cm、3 cm以及5 cm的三个发育阶段的2室角果,其带型之间无明显差异。同样地,长度分别为1 cm、3 cm以及5 cm的三个发育阶段的3室角果,其带型之间也无明显差异。但是,长度为1 cm的3室角果,其酶带染色最深,说明其酶活性明显比长度3 cm和5 cm角果的酶活性强。
3.2.2. 三棱角材料14ds72三室与两室角果的POD同工酶比较
三棱角材料14ds72的POD同工酶电泳结果如图5所示,由图5可知:在长度分别为1 cm、3 cm以及5 cm的三个发育阶段的角果中,3室角果与2室角果的POD同工酶带型一致,条带数目相同,只是少数条带的染色深浅存在一定差异,说明这些条带的酶活性存在一定差异。如对长度为1 cm的角果而言,其3室角果比2室角果对应的条带染色深,说明3室角果比2室角果对应的条带活性强。又如对长度为5 cm的角果而言,其3室角果2条迁移率中等且相近的较强酶带比2室角果对应的条带染色要深,说明3室角果这2条较强带比2室角果对应的条带活性要强,而其3室角果1条迁移率最大的强带比2室角果对应的条带染色要浅,说明3室角果这条迁移率最大的强带比2室角果对应的条带活性要弱。
另外,由图5还可知:长度分别为3 cm和5 cm的两个发育时期的2室角果,其带型之间差异不明显。同样地,长度分别为3 cm和5 cm的两个发育时期的3室角果,其带型之间也无明显差异。但是,长度为1 cm的2室角果和3室角果,其酶带染色最深,说明长度1 cm的角果的酶活性明显比长度3 cm和5 cm角果的酶活性强,而且其在迁移率中等的区域与迁移率小的区域之间似乎还增加了1条较强带。
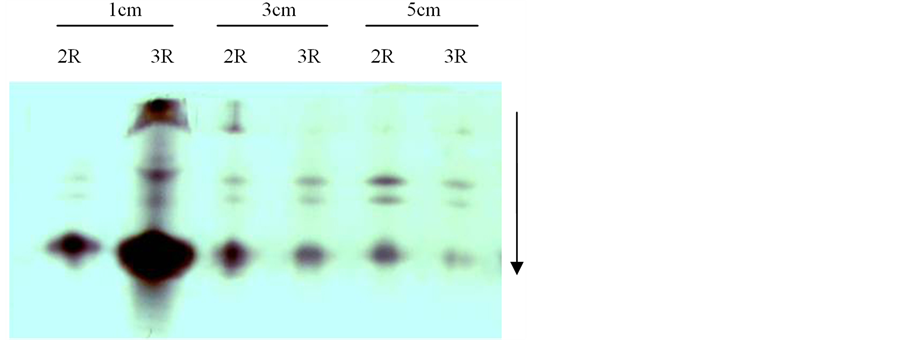
Figure 4. Electrophoresis of POD isozymograms on 14ds67; 2R: 2 rooms; 3R: 3 rooms; Sample volumes: 50 μl, the same as follow
图4. 14ds67的POD同工酶电泳结果;2R:2室;3R:3室;点样量:50 μl。下同

Figure 5. Electrophoresis of POD isozymograms on 14ds72
图5. 14ds72的POD同工酶电泳结果
3.2.3. 三棱角材料14ds78三室与两室角果的POD同工酶比较
三棱角材料14ds78的POD同工酶电泳结果如图6所示,由图6可知:在长度分别为1 cm、3 cm以及5 cm的三个发育时期的角果中,3室角果与2室角果的POD同工酶带型一致,酶带数目相同,只是少数酶带的染色深浅出现一定的差异,说明这些条带的酶活性存在一定的差异。如对长度为3 cm的角果而言,其3室角果比2室角果对应的条带染色深,说明3室角果比2室角果对应的条带酶活性强。又如对长度为5 cm的角果而言,其3室角果比2室角果对应的条带染色浅,说明3室角果比2室角果对应的条带酶活性弱。
另外,由图6还可知:长度分别为1 cm、3 cm以及5 cm的三个发育时期的2室角果,其带型之间无明显差异。同样地,长度分别为1 cm、3 cm以及5 cm的三个发育时期的3室角果,其带型之间也无明显差异。只是随着角果的生长发育,迁移率小的弱带的染色变浅,以至消失。也就是说,该酶带在角果发育早期酶活性强,但是随着角果的生长发育其酶活性水平越来越低。另外,长度为1 cm的2室角果和3室角果,其酶带染色最深,也说明长度1 cm角果的酶活性明显较长度3 cm和5 cm角果的酶活性强。
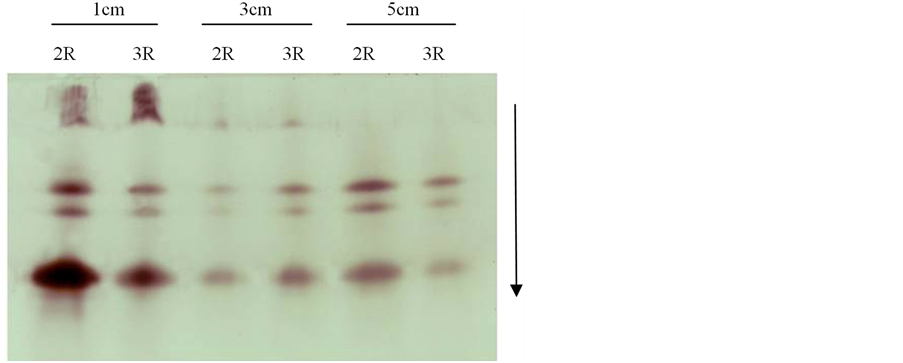
Figure 6. Electrophoresis of POD isozymograms on 14ds78
图6. 14ds78的POD同工酶电泳结果
3.2.4. 3个三棱角材料之间的POD同工酶酶谱比较
比较图4~图6可知,3个三棱角材料14ds67、14ds72以及14ds78的POD同工酶酶谱相似,带型基本一致,只是部分条带的酶活性有所不同。表现在在3个材料的POD同工酶酶谱中,分别有一条染色深的强带,其迁移率最大,位于酶谱的下部;有2条染色较深的较强带,其迁移率中等,位于酶谱的中间部位;有1条染色浅的弱带,其迁移率最小,位于酶谱的上部。而且,3室角果与2室角果的POD同工酶带型相同,没有明显差异,只是少数条带的染色程度有一定的差异,说明这些条带的酶活性存在一定的差异。
4. 结论
本文通过对甘蓝型油菜3个三棱角材料的EST和POD同工酶的凝胶电泳和酶谱分析,探讨了三棱角果(三室)和普通角果(两室)在同工酶水平上的异同性。实验结果表明:三室角果和两室角果的EST同工酶,其带型表现一致,只是个别同工酶的酶活性有一定的差异;两者的POD同工酶,其带型也表现一致,只是少数同工酶的酶活性出现一定的差异。而且,对两种同工酶而言,角果越小,酶活性越强,即随着角果的生长发育其酶活性表现为减弱的趋势。
5. 讨论
如文中所述,有关多室油菜的研究,目前已经取得了一定的进展 [1] - [14] 。然而,有关多室油菜同工酶方面的研究尚属空白,为了探讨三棱角材料三室角果与两室角果在同工酶水平上的异同性,故开展本研究工作。
三棱角(三室)油菜是笔者近年来自主选育的一种甘蓝型多室油菜类型 [8] [13] [14] ,在选育过程中,三棱角材料的植株常表现为三室角果与两室角果共存的现象,即存在三室角果与两室角果的嵌合现象。本研究首次以3个三棱角材料的三室角果与两室角果为研究对象,通过对其长度分别为1 cm、3 cm以及5 cm三个不同发育阶段幼嫩角果的EST、POD同工酶酶谱的比较分析,从而证明三棱角材料的三室角果与两室角果之间的同工酶带型分别表现一致,无明显差异,只是少数酶带的酶活性大小存在一定的差异,这为进一步研究三棱角油菜提供了参考依据。
同工酶酶谱分析表明:1) 三室角果与两室角果在长度为1 cm、3 cm以及5 cm的三个不同发育阶段的EST、POD同工酶带型分别无明显差异,只是少数酶带的活性有一定的差异;2) 同一材料不同发育阶段的两室角果的EST、POD同工酶带型分别无明显差异,只是少数条带的酶活性有一定的差异,且角果越小,酶活性越强,说明角果在由小到大的生长发育过程中酶活性表现为减弱的趋势;3) 在3个三棱角材料中,14ds67与14ds72的EST同工酶酶谱相似程度较高,带型一致,只是个别条带的酶活性大小存在一定的差异。然而,14ds78的EST同工酶酶谱与前两个材料之间有较大差异,这种差异主要表现在第II区(中速区),其中位于中间位置的2~3条强带变成极弱带或者条带消失,说明其酶活性显著降低。由此可见,三棱角材料14ds67与14ds72的亲缘关系较近,而14ds78与前两者相比较亲缘关系较远。另外,3个三棱角材料之间的POD同工酶酶谱相似程度较高,其带型一致,只是少数条带的酶活性表现出一定差异。
本研究还表明,三棱角材料的EST同工酶酶谱的条带数目较多,而POD同工酶酶谱的条带数目较少,即同一材料的EST同工酶酶谱的条带数目明显多于POD同工酶酶谱的条带数目。
基金项目
陕西省自然科学基础研究计划项目(2015JM3107)。
文章引用
朱彦涛,张新,苟升学,赵亚军,陈文杰,罗斌. 三棱角油菜两类角果的酯酶同工酶和过氧化物酶同工酶比较分析
Analysis of Esterase Isozymograms and Peroxidase Isozymograms on Two Kinds of Pods of Triquetrum-Pod Rapeseed[J]. 植物学研究, 2017, 06(03): 114-123. http://dx.doi.org/10.12677/BR.2017.63016
参考文献 (References)
- 1. 何余堂, 涂金星, 傅廷栋, 等. 中国白菜型油菜种质资源的遗传多样性研究[J]. 作物学报, 2002, 28(5): 697-703.
- 2. 何余堂, 龙卫华, 胡进平, 等. 白菜型油菜角果多室性状的遗传及解剖学研究[J]. 中国油料作物学报, 2003, 25(1): 1-4.
- 3. 何余堂. 白菜型油菜的遗传多样性及特殊种质资源的研究[D]: [博士学位论文]. 武汉: 华中农业大学, 2003.
- 4. 赵洪朝, 杜德志, 刘青元, 等. 芥菜型多室油菜的主要性状表现[J]. 西北农业学报, 2003, 12(3): 62-64.
- 5. 赵洪朝, 杜德志, 刘青元, 等. 芥菜型油菜多室性状的遗传研究[J]. 西北农林科技大学学报(自然科学版), 2003, 31(6): 90-92.
- 6. 赵洪朝, 杜德志, 刘青元, 等. 芥菜型多室油菜与甘蓝型油菜的种间远缘杂交[J]. 西北植物学报, 2003, 23(9): 1587-1591.
- 7. 赵洪朝. 芥菜型油菜多室性状的遗传表现及其与甘蓝型油菜的种间远缘杂交[D]: [硕士学位论文]. 杨凌: 西北农林科技大学, 2003.
- 8. 朱彦涛. 甘蓝型三棱角油菜新材料的遗传与选育研究初报[J]. 西北农业学报, 2005, 14(1): 23-26.
- 9. Zhao, H.C., An, F.Y. and Du, D.Z. (2007) New Idioplasmic Resource B. napus L. with Multi-Loculus Founded by Interspecific Hybridization. In: Fu, T.D., Ed., Proceedings of the 12th International Rapeseed Congress, Wuhan, Vol. 3, 294-295.
- 10. 赵文路. 种间杂交创建的甘蓝型多室油菜的多室等位性研究及其不育材料JN55A不育类型的鉴别[D]: [硕士学位论文]. 西宁: 青海大学, 2007.
- 11. 吕泽文. 芥菜型油菜多室性状的遗传分析及其基因的分子标记[D]: [硕士学位论文]. 武汉: 华中农业大学, 2011.
- 12. 吕泽文, 徐平, 张向向, 等. 芥菜型油菜多室角果的解剖特征及遗传分析[J]. 中国油料作物学报, 2012, 34(5): 461-466.
- 13. 朱彦涛, 胡选萍, 刘 湛, 等. 油菜角果多室性状及其研究进展[J]. 中国油料作物学报, 2012, 34(3): 321-325.
- 14. 朱彦涛, 田建华. 甘蓝型油菜三棱角自交群体的角果性状[J]. 中国油料作物学报, 2015, 37(5): 730-734.
- 15. 王保仁, 常桂菊, 胡能书. 湘矮型油菜三系及其杂种过氧化物酶同功酶的初步研究[J]. 湖南农业科技, 1980(6): 16-21.
- 16. 刘忠松, 官春云. 油菜雄性不育性的研究——甘蓝型油菜波里马(Polima)细胞质雄性不育系与保持系的生化比较[J]. 中国油料, 1990(3): 1-5.
- 17. 陈松, 浦惠明, 戚存扣, 等. 甘蓝型双低油菜三系及杂种酯酶同工酶分析[J]. 江苏农业科学, 1995(2): 27-28.
- 18. 伊岚, 来航线, 李殿荣, 等. 甘兰型油菜细胞质雄性不育系陕2A和保持系陕2B的生化比较[J]. 西北农业学报, 1997, 6(2): 39-41.
- 19. 林宝刚. 甘蓝型油菜细胞质雄性不育系Pol、Shan2A、Ogu和Gli的比较研究[D]: [硕士学位论文]. 杭州: 浙江大学, 2005: 38-40.
- 20. 于澄宇, 胡胜武, 郭蔼光, 等. 甘蓝型油菜5个同质CMS系及其保持系、杂交种的同工酶比较[J]. 西北农业学报, 2003, 12(1): 5-7.
- 21. 朱彦涛, 张新, 刘湛, 等. 2个油菜CMS系统的酯酶和过氧化物酶同工酶分析[J]. 西北植物学报, 2009(4): 711- 716.