Advances in Applied Mathematics
Vol.
11
No.
12
(
2022
), Article ID:
59554
,
9
pages
10.12677/AAM.2022.1112939
具有Logistic增长的蚊群动力学模型的 稳定性分析
黄晓霞*,杨悦,吕贵臣*
重庆理工大学理学院,重庆
收稿日期:2022年11月23日;录用日期:2022年12月16日;发布日期:2022年12月27日
摘要
疟疾、登革热等蚊媒传染病的高致病率和高死亡率给全球公共卫生带来了巨大的挑战。本文建立了一个具有Logistic增长的蚊群传播动力学模型,通过计算系统的基本再生数,得到了系统平衡点的存在性,并结合单调动力系统理论,得到了系统平凡平衡点和正平衡点的全局渐近稳定性。
关键词
蚊媒传染病,蚊群动力学模型,单调动力系统理论

Stability Analysis of Mosquito Population Dynamics Model with Logistic Growth
Xiaoxia Huang*, Yue Yang, Guichen Lyu*
School of Science, Chongqing University of Technology, Chongqing
Received: Nov. 23rd, 2022; accepted: Dec. 16th, 2022; published: Dec. 27th, 2022

ABSTRACT
The high morbidity and mortality rates of mosquito-borne infectious diseases such as malaria and dengue pose a huge challenge to global public health. In this paper, a transmission dynamic model of mosquito population with Logistic growth is established. The existence of the equilibrium points is obtained by calculating the basic regeneration number of the system, and the global asymptotic stability of the trivial equilibrium point and the positive equilibrium point is obtained by combining the theory of monotone dynamic system.
Keywords:Mosquito-Borne Infectious Diseases, Mosquito Population Dynamic Model, Monotone Dynamic System Theory

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
疟疾、登革热、基孔肯雅热与寨卡等传染病都是蚊媒传染病,健康蚊虫通过吸食患病人类的血液致使自身感染,而后携带病菌的蚊虫通过吸食健康人类的血液,又将疾病传播给健康人群。蚊媒传播疾病的过程异常复杂,每年造成全球大量传染病病例,致死率较高,严重威胁着人类的健康与公共安全。
为了预防与控制蚊媒传染病,Ross提出了疟疾传播的第一个数学模型,并指出若将蚊虫数量保持在某个临界值之下,则可有效降低疟疾传染病的传播风险 [1]。在Ross工作的基础上,Macdonald改进了Ross模型,提出了Ross-Macdonald传染病模型,表明有效地减少蚊群数量对疟疾传播有一定的影响 [2]。
由于目前没有有效的疫苗来控制疾病的传播,因此常用的控制蚊虫数量的方法是使用杀虫剂。然而这种方法不仅会对环境造成污染而且还使得部分蚊虫产生抗药性。为此学者们提出了不育昆虫技术(SIT)及Wolbachia氏菌等生物基因技术来减少蚊群数量。不育昆虫技术(SIT)最早由Knipling和他的合作者们提出 [3],他们在50年代早期首次成功进行了实验,当时几乎消灭了佛罗里达州的螺旋虫种群。SIT是一种生物控制技术,它通过诱变剂来破坏蚊虫的繁殖过程,从而使雌蚊不育。这个过程一般通过向环境中释放不育的雄蚊,野生雌蚊与之交配所产卵不会孵化。若向环境中释放的雄蚊足够多,那么野生蚊虫的数量将会减少,进而导致野生蚊虫种群的灭绝。
考虑到SIT技术和Wolbachia氏菌在控制蚊群数量方面的有效性,学者尝试构建数学模型来开展SIT等生物控制技术的研究 [3] - [18]。为了对比杀虫剂与SIT方法的效果和成本,Thome等在文献 [4] 中提出了一个最优控制模型来评估这两种措施的有效性,他们通过成本最小化得到最优控制策略,并指出在蚊虫防治初期需要大量使用杀虫剂,后期再使用SIT等生物控制技术会取得更好的效果。他们的研究为蚊虫防治工作提供了一个新的思路。Anguelov等在文献 [5] 中将蚊虫生命周期分为水生阶段和成虫阶段,建立了一个野生蚊虫种群模型,而后在此模型基础上考虑SIT技术,引入人工释放的不育雄蚊,通过稳定性分析和数值模拟得到了有效控制野生蚊虫数量的一个阈值。然而,对于白蚊伊蚊这种快速繁殖的大种群物种,SIT方法的效果可能比较有限,于是一种名为“强化SIT”的新方法应运而生,Haramboure等基于SIT和强化SIT方法研究了白蚊伊蚊的种群模型,探究白蚊伊蚊的防治策略 [7]。考虑疾病在人群和蚊群的传播,Moulay等讨论了蚊群动力学和病毒向人类传播的模型,并利用Lyapunov稳定性定理和竞争系统理论,给出平衡点的稳定性及周期解的轨道稳定性 [9]。Xue等通过建立数学模型,比较了不同菌株的Wolbachia氏菌在控制基孔肯雅病毒、登革热病毒、寨卡病毒传播方面的有效性 [13]。罗育东等考虑了蚊群的强Alle效应,构造了包括野生蚊群和不育蚊群的连续时间模型 [15]。武丹在文献 [16] 中建立了Wolbachia氏菌在蚊群中传播的微分方程模型,并考虑时滞因素,提出了一个时滞微分方程模型。陈盼盼等人在文献 [17] 中建立了工程菌在蚊群中传播的模型,考虑了蚊虫间工程菌的水平传播和垂直传播,研究了工程菌对控制疟疾的有效性。张金金等建立了Wolbachia氏菌的蚊群模型,通过考虑不完全细胞质不相容情况得到了模型的鞍结点分支 [18]。
本文的主要目标是建立和分析阶段结构的蚊群动力学模型,并考虑产卵行为及生存资源对蚊群的影响。文章接下来的安排如下:第二部分给出本文主要用到的基础知识;第三部分我们提出具有生存资源限制的蚊群动力学模型,并给出模型平衡点的存在性及其稳定性;第四部分我们对全文的主要内容做一个总结。
2. 预备知识
考虑如下自治动力系统
(2.1)
其中 , 记过初值 的解为 。
引理1 [19] 假设系统(2.1)满足:对于任意的i,当 且 时,有 ,那么当 , 时,有 。
定义2 [19] 若系统(2.1)中雅可比矩阵 的非对角线上的元素均非负,即对所有的 ,当 时,有 ,则称系统(2.1)为合作系统。
引理3 [4] 若系统(2.1)满足以下条件:
a) 系统(2.1)是合作系统,
b) 存在 且 ,,满足 ,
c) 区间 内存在唯一一个平衡点p,
则p在区间 上是全局渐近稳定的。
3. 蚊群动力学模型及其稳定性分析
3.1. 模型的建立
蚊虫的生命周期包括卵、幼虫、蛹和成虫四个阶段。蚊虫从卵发育至幼虫一般需要1~2周时间,之后变成蛹,再经过两天左右成熟,变成成虫(雌蚊、雄蚊)。而雌蚊通过与雄蚊交配而产卵。雌蚊会在有水的容器中产卵,而卵需要在水中孵化变成幼虫,幼虫需要在水中发育成为蛹。卵和幼虫阶段的发育受环境和资源的限制,所以我们用Logistic增长来描述卵和幼虫的增长,Logistic增长模型又称为自我抑制性方程,常用来描述单个生物种群的数量变化 [3] [9]。
我们将某个环境中的蚊群分成6个仓室,即卵(E)、幼虫(L)、蛹(P)、不产卵的年轻雌蚊(Y)、产卵的成熟雌蚊(F)和野生雄蚊(M)。令 、 、 、 、 、 分别表示t时刻各个仓室的种群数量。该蚊虫种群的总数量用 表示,则 。
考虑到蚊虫的生命周期过程,我们作如下假设:
(A1) 蚊虫的产卵量与成熟雌蚊有关且蚊子具备制定产卵栖息地的能力,若雌蚊发现产卵地点已经存在大量的卵或是营养资源匮乏时,它将在此处产较少的卵或直接转换新的产卵地。在该假设下,我们假设平均产卵率为 ,这里的b为内禀产卵率, 为卵的环境容纳量。
(A2) 幼虫的增长率与卵的数量有关且受到营养资源的影响,而且存在种内竞争,进而其增长率为 。这里 表示蚊卵向幼虫转化的转化率, 为幼虫的环境容纳量。
(A3) 蛹的单位增长率与幼虫的数量成正比,转化系数为 。
(A4) 蛹羽化成为成虫的羽化率为 ,一部分转化为未产卵的年轻雌蚊(比例为r),剩余部分转化为雄蚊(比例为 )。
(A5) 年轻雌蚊通过与雄蚊交配,且一生只交配一次,从而转化成为产卵的成熟雌蚊,转化率为 。
基于上述假设,得到蚊群生命周期演化图如下(图1)。
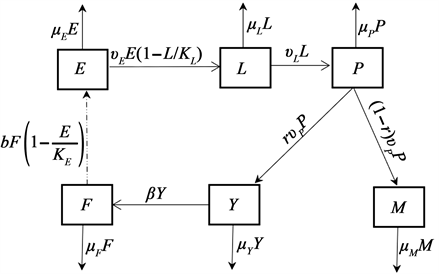
Figure 1. Flow chart of the evolution of the mosquito life cycle
图1. 蚊群生命周期演化流程图
基于流程图1,我们得到蚊群传播动力学模型:
(3.1)
其中 为相应仓室的死亡率。
3.2. 不变性
令 ,其中
则系统(3.1)可写成如下形式
(3.2)
定义集合D如下:
其中 ,,,。
定理4 集合D关于系统(3.1)是正不变的。
证明:任取 ,易知
由本文第一部分给出的引理1知,对任意的 ,系统(3.1)的解 。即集合D关于系统(3.1)是正不变的。另一方面,对于任意的 ,我们说明 。即:
1) 若 ,则 , ;
2) 若 ,则 , ;
3) 若 ,则 , ;
4) 若 ,则 , ;
5) 若 ,则 , ;
6) 若 ,则 ,。
我们只需证明情形(1)和(3)成立即可,其余情形类似可证。
对情形(1)和(3)的证明如下:
首先,证明情形(1)成立。假设结论(1)不成立,即 ,使得 。
令 ,则 且 ( ),故 ,而由模型(3.1)知
这与 矛盾,进而情形(1)成立。
然后,证明情形(3)成立。假设情形(3)不成立,即 ,使得 。
令 ,则 且 ( ),故 ,而由结论(2)知,当 时, ,我们分以下两种情况讨论:
(3a) 若 ,则根据模型(3.1)知
这与 矛盾。
(3b) 若 ,则 ,而
故由Taylor展开式知, ,
由无穷小的性质得, ,当 时,有
因此,当 时, ,这与假设矛盾,故而情形(3)成立。
定理5 集合D是系统(3.1)的全局吸引子,即 ,,当 时, 。
证明:由定理4的证明过程知, , 且集合D关于系统(3.1)是正不变的。因此,我们仅需证明,存在时刻 使得 即可。
1) 假设对于任意 ,,则 ,从而由系统(3.1)的第一个方程得
故 ,进而由迫敛性知 ,这与假设矛盾。因此,存在 使得 。
2) 若 ,则由定理4知,对任意 ,,。
若 且对任意 ,,则 ,故由系统(3.1)的第二个方程有
因此,对任意 ,,由极限理论知 ,这与假设矛盾。故存在 ,使得 。
3) 若 ,则对任意 ,。
若 ,且对任意的 ,,此时由(1)(2)知, ,,因此
进而对任意的 ,
这与假设矛盾,故存在 ,使得 。进而对任意的 ,有 ,,。
4) 若 ,则由定理4,对任意的 ,。否则类似(3)的证明可得,存在 ,使得 。进而对任意的 ,有 ,,,。
5) 若 ,则由定理4知,对任意的 ,。否则类似(3)的证明可得,存在 ,使得 。
6) 若 ,则由定理4知,对任意的 ,。否则类似(3)的证明可得,存在 ,使得 。
由(1)~(6)知,存在 ,使得 ,进而命题得证。
3.3. 平衡点的存在性与稳定性
易知系统(3.1)始终存在一个平凡平衡点 。引入阈值常数
,
容易验证,当 时,系统(3.1)存在一个正的非平凡平衡点 ,其中
,,
,,
,。
定理6 i) 当 时,系统(3.1)的平衡点 在 上是全局渐近稳定的;
ii) 当 时,系统(3.1)的平衡点 是不稳定的,平衡点 在 上是全局渐近稳定的。
证明:由定理5知,D是全局吸引子,因此我们仅需证明结论在D内成立即可。
计算可得系统(3.1)的雅可比矩阵为
易知 在D内具有非负的非对角项,所以系统(3.1)在D内是合作系统。
取
,,
,,
令 ,那么
i) 当 时,系统(3.1)在 上只有唯一一个平衡点 。在区间 上有, 。那么根据引理3可得,平衡点 在区间 上是全局渐近稳定的。另一方面,对于任意的 ,都有 ,进而
综上,当 时,系统(3.1)的平衡点 在D上是全局渐近稳定的。
ii) 当 时,任取 ,使得下列不等式成立:
令 ,从而在区间 上,有 。
在 的情况下,平衡点 是系统(3.1)在区间 上的唯一一个平衡点。从而由定理3,平衡点 在区间 上是全局渐近稳定的。另一方面,对任意的 ,可以选取充分小的 ,使得 ,进而
综上,当 时,系统(3.1)的平衡点 在D上是全局渐近稳定的。
下面再说明平衡点 的不稳定性。记矩阵A为g对x在 处的雅可比矩阵,则
记行列式 为行列式 的五阶顺序主子式,由于 ,所以
,
这说明矩阵A至少有一个实部为正的特征根,从而由文献 [20] 中的稳定性理论得,当 时,平衡点 是不稳定的。
4. 总结
本文基于蚊虫的实际生长周期情况建立了具有资源限制的蚊虫种群传播动力学模型。我们根据Smith-Waltman不变性原理证明了可行域关于系统的正不变性,在此基础上,结合数学分析的知识验证了系统的可行域为全局吸引子。通过计算和分析,得到了模型的平凡平衡点和正平衡点的存在性及稳定性的阈值条件。运用合作系统的单调系统理论和引理3进行分析和论证,构造使用定理所需要的区间条件。最后得到系统的全局动力学性态:当阈值小于等于1时,平凡平衡点是全局渐近稳定的;当阈值大于1时,平凡平衡点变得不稳定,正平衡点是全局渐近稳定的。
文章引用
黄晓霞,杨 悦,吕贵臣. 具有Logistic增长的蚊群动力学模型的稳定性分析
Stability Analysis of Mosquito Population Dynamics Model with Logistic Growth[J]. 应用数学进展, 2022, 11(12): 8908-8916. https://doi.org/10.12677/AAM.2022.1112939
参考文献
- 1. Ross, R. (1911) The Prevention of Malaria. 2nd Edition, John Murray, London.
- 2. Macdonald, G. (1957) The Epi-demiology and Control of Malaria. Oxford University Press, Oxford.
- 3. Strugarek, M., Bossin, H. and Dumont, Y. (2019) On the Use of the Sterile Insect Release Technique to Reduce or Eliminate Mosquito Populations. Applied Math-ematical Modelling, 68, 443-470. https://doi.org/10.1016/j.apm.2018.11.026
- 4. Thomé, R.C.A., Yang, H.M. and Esteva, L. (2010) Optimal Con-trol of Aedes aegypti Mosquitoes by the Sterile Insect Technique and Insecticide. Mathematical Biosciences, 223, 12-23. https://doi.org/10.1016/j.mbs.2009.08.009
- 5. Anguelov, R., Dumont, Y. and Lubuma, J. (2012) Mathematical Modeling of Sterile Insect Technology for Control of Anopheles Mosquito. Computers and Mathematics with Applica-tions, 64, 374-389. https://doi.org/10.1016/j.camwa.2012.02.068
- 6. Tapi, D., Beihle, B., Bowong, S. and Dumont, Y. (2018) Mod-els for Miridae, a Cocoa Insect Pest. Application in Control Strategies. Mathematical Methods in the Applied Sciences, 41, 8673-8696. https://doi.org/10.1002/mma.5063
- 7. Haramboure, M., Labbé, P., Baldet, T., et al. (2020) Mod-elling the Control of Aedes albopictus Mosquitoes Based on Sterile Males Release Techniques in a Tropical Environment. Ecological Modelling, 424, Article ID: 109002. https://doi.org/10.1016/j.ecolmodel.2020.109002
- 8. Dumont, Y. and Yatat-Djeumen, I. (2022) Sterile Insect Technique with Accidental Releases of Sterile Females. Impact on Mosquito-Borne Diseases Control When Viruses Are Circulating. Mathematical Biosciences, 343, Article ID: 108724. https://doi.org/10.1016/j.mbs.2021.108724
- 9. Moulay, D., Aziz-Alaoui, M. and Cadivel, M. (2011) The Chikungunya Disease: Modeling, Vector and Transmission Global Dynamics. Mathematical Biosciences, 229, 50-63. https://doi.org/10.1016/j.mbs.2010.10.008
- 10. Danbaba, U. and Garba, S. (2018) Modeling the Transmission Dynamics of Zika with Sterileinsect Technique. In: Anguelov, R. and Lachowicz, M., Eds., Mathematical Methods and Models in Biosciences, Biomath Forum, Sofia, 81-99. https://doi.org/10.11145/texts.2018.01.083
- 11. Almeida, L., Duprez, M., Privat, Y. and Vauchelet, N. (2022) Op-timal Control Strategies for the Sterile Mosquitoes Technique. Journal of Differential Equations, 311, 229-266. https://doi.org/10.1016/j.jde.2021.12.002
- 12. Bakhtiar, T., Fitri, I., Hanum, F and Kusnanto, A. (2022) Mathemat-ical Model of Pest Control Using Different Release Rates of Sterile Insects and Natural Enemies. Mathematics, 10, Arti-cle No. 883. https://doi.org/10.3390/math10060883
- 13. Xue, L., Fang, X. and Hyman, J. (2018) Comparing the Effectiveness of Different Strains of Wolbachia for Controlling Chikungunya, Dengue Fever, and Zika. PLOS Neglected Tropical Diseases, 12, e0006666. https://doi.org/10.1371/journal.pntd.0006666
- 14. 庞轶友. 不育蚊子动力学模型研究[D]: [硕士学位论文]. 吉林: 长春理工大学, 2022.
- 15. 罗育东, 张国洪. 具有强Allee效应的野生蚊子和绝育蚊子相互作用模型[J]. 西南大学学报(自然科学版), 2019, 41(1): 65-71.
- 16. 武丹. 一类Wolbachia氏菌在蚊群传播的数学模型的动力学研究[D]: [硕士学位论文]. 广州: 广州大学, 2022.
- 17. 陈盼盼, 钟浩南, 王稳地. 基于工程菌控制蚊子的动力学建模与分析[J]. 西南师范大学学报(自然科学版), 2021, 46(5): 25-31.
- 18. 张金金, 王稳地, 舒梦诗, 姚苗然. 沃尔巴克氏体在蚊子种群中的传播动态分析[J]. 西南大学学报(自然科学版), 2017, 39(3): 81-87.
- 19. 吕贵臣, 陆征一. 高维系统稳定性的几何判据[M]. 北京: 科学出版社, 2019.
- 20. 马知恩, 周义仓, 李承治. 常微分方程定性与稳定性方法[M]. 北京: 科学出版社, 2015.
NOTES
*通讯作者。