Advances in Geosciences
Vol.
12
No.
03
(
2022
), Article ID:
49595
,
12
pages
10.12677/AG.2022.123030
鄂南奥陶系平凉组微生物碳酸盐岩 特征与储集意义
韩月卿1,2*,刘仪3,岳小娟4,张军涛1,2#,金晓辉1,2,韩文彪5,赵红琴6
1中国石化深部地质与资源重点实验室,北京
2中国石化石油勘探开发研究院,北京
3中国地质大学(北京)能源学院,北京
4中国石油大学(北京)地球科学学院,北京
5中国石化油田部,北京
6中国石化中原油田分公司物探研究院,河南 濮阳
收稿日期:2022年2月21日;录用日期:2022年3月15日;发布日期:2022年3月23日

摘要
微生物碳酸盐岩储层是近年来石油地质领域研究的热点之一。微生物结构在储层储集空间的形成过程中扮演重要角色。鄂尔多斯盆地发育大量微生物碳酸盐岩,但关于鄂尔多斯盆地南部奥陶纪微生物碳酸盐岩鲜有报道。本文对鄂南陈炉任家湾剖面进行详细野外观测和薄片观察,描述了微生物岩的岩石学特征和储集空间特征,综合分析了微生物岩的沉积环境,探讨了微生物碳酸盐岩孔隙演化与成岩过程的关系。研究认为:鄂尔多斯盆地南部奥陶系平凉组发育含生屑泥晶灰岩、砾屑灰岩、砂屑灰岩和藻灰岩四种岩石类型,微生物碳酸盐岩主要发育在藻灰岩中,以凝块石最多,叠层石次之,树形石最少。陈炉任家湾剖面平凉组沉积时期属于台地边缘相–台地前缘斜坡相环境,而凝块石和叠层石主要发育在台地边缘相之中相对低能的环境,且鄂南平凉组中的礁、滩、微生物丘往往共生。微生物碳酸盐岩储层储集空间类型多样,以葡萄花边构造溶孔、凝块颗粒间溶孔、海绵体腔孔、层间溶孔、窗格孔为主。微生物碳酸盐岩储层的形成受微生物结构、成岩作用以及沉积微相等多种因素共同控制。储集空间受到微生物结构的影响,在准同生期受溶蚀改造形成,其后又经历了表生期溶蚀改造,但经过充填胶结作用后,孔隙减小或消失。沉积微相总体上控制着微生物岩储层类型及其展布特征。鄂尔多斯盆地南部奥陶系可能存在微生物岩储层,微生物碳酸盐岩储层将是今后的油气勘探中的一类重要类型。
关键词
微生物岩,凝块石,叠层石,树枝石,平凉组,奥陶系,鄂尔多斯盆地

The Characteristics and Storage Significance of Microbial Carbonate Rocks in the Ordovician Pingliang Formation of Southern Ordos Basin
Yueqing Han1,2*, Yi Liu3, Xiaojuan Yue4, Juntao Zhang1,2#, Xiaohui Jin1,2, Wenbiao Han5, Hongqin Zhao6
1Key Laboratory of Deep Geology and Resources, Sinopec, Beijing
2Sinopec Petroleum Exploration and Development Research Institute, Beijing
3School of Energy, China University of Geosciences, Beijing
4School of Earth Sciences, China University of Petroleum (Beijing), Beijing
5Department of Oil-Field, SINOPEC, Beijing
6Institue of Geophysical Exploration, Zhongyuan Oilfield Company, Puyang Henan
Received: Feb. 21st, 2022; accepted: Mar. 15th, 2022; published: Mar. 23rd, 2022

ABSTRACT
Microbial carbonate reservoir is one of the research hotspots in petroleum geology in recent years. Microbial structure plays an important role in the formation of reservoir space. A large number of microbial carbonate rocks are developed in Ordos Basin. However, there is no report on Ordovician Microbial Carbonates in the southern Ordos Basin. In this paper, the field observation and thin slice observation are carried out in the field outcrops of Chenlu Renjiawan in the southern Erdos basin. The petrological characteristics and reservoir space characteristics of the Microbial Carbonates are described. The sedimentary environment of the Microbial Carbonates is synthetically analyzed, and the relationship between the pore evolution and the diagenesis process of the Microbial Carbonates is discussed. The study suggests that: Four types of rock containing bioclastic mudstone limestone, gravel limestone, sand-clastic limestone, and algal limestone are developed in Ordovician Pingliang Formation in the southern Ordos Basin. Microbial carbonate is mainly developed in algal limestones. Most of them are thrombolites, followed by stromatolites and dentrolite. The depositional period of the Pingliang Formation in the field outcrop of Chenlu Renjiawan belongs to platform margin facies to platform foreslope facies, while the thrombolites and the stromatolites are mainly developed in the relatively low energy environment in platform margin facies. The reefs, shoals and microbial carbonate rocks in the Pingliang formation of southern Erdos are often symbiotic. There are various types of reservoir space in microbial carbonate reservoirs, mainly including grape-lace structure pores, intergranular dissolved pores of thrombolites, cavernous pores, interlaminar pores and pane pores. The formation of microbial carbonate reservoir is controlled by many factors such as microbial structure, diagenesis and sedimentary microfacies. The reservoir space is affected by the microbiological structure. At the penecontemporaneous stage, the reservoir is transformed by precipitation corrosion, followed by a supergene corrosion modification. However, after the cementation, the pores decrease or disappear. The sedimentary microfacies control the type of reservoir and its distribution characteristics in general. The microbial carbonate reservoirs in the Ordovician system in the southern Ordos basin may exist, and the microbial carbonate reservoir will be an important type of oil and gas exploration in the future.
Keywords:Microbial Rocks, Thrombolite, Stromatolite, Dentrolite, Pingliang Formation, Ordovician, Ordos Basin

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
近年来,在微生物碳酸盐岩中的油气勘探取得了重大突破。如巴西桑托斯盆地下白垩统、哈萨克斯坦的滨里海盆地、美国阿拉巴马州墨西哥湾东部和北部上侏罗统Smackover组、阿曼南部的阿曼盐盆的上震旦–下寒武统的Ara群等地区的微生物碳酸盐岩中都发现了大规模的油气储量;在我国塔里木盆地上震旦统和下寒武统、奥陶系,四川盆地震旦系灯影组,川西中三叠统雷口坡组和渤海湾盆地任丘油田中元古界雾迷山组的微生物碳酸盐岩的油气勘探也已取得了较大进展 [1] [2] [3] [4] [5]。微生物碳酸盐岩储层相关研究越来越引起地质学家的重视。
微生物碳酸盐岩是由底栖微生物群落(细菌、真菌、小型藻菌和原生动物)通过微生物生长、新陈代谢、细胞表面物质性质、胞外聚合物(Extracellular Polymeric Sub-stances, EPS)与矿物沉淀、颗粒捕获相互作用形成。微生物碳酸盐岩在整个地质历史时期中从早太古代至今均有分布,其中中新元古代、寒武纪和奥陶纪尤为发育 [6] [7] [8] [9] [10]。微生物碳酸盐岩的分类方法较多,以古微生物的生态可以划分为叠层石生态系和非叠层石生态系,前者包括叠层石、层纹石、核形石等,后者包括非造席的单细胞生物蓝细菌形成的球粒–椭球粒凝块石等 [11] [12];又可根据形态学特征分为叠层石、凝块石、树形石、均一石、纹理石和核形石 [13] [14] [15]。
凝块石和叠层石是微生物碳酸盐岩中最主要的两种岩石储层类型 [16] [17] [18] [19] [20],储层形成受控于微生物结构、白云化作用、溶蚀作用和构造作用等 [21] [22]。微生物碳酸盐岩储集空间类型受复杂微生物结构影响,包括:凝块间溶孔、葡萄花边状溶孔、喀斯特溶孔、微生物体腔孔、微生物格架孔、粒间溶孔等 [23]。
微生物碳酸盐岩沉积环境较广,台地边缘、斜坡以及台地内部都可发育,还能与生物礁、滩相伴生。不同的沉积微相和复杂的微生物结构控制着微生物碳酸盐岩储层类型及空间展布特征 [24] [25]。
鄂尔多斯盆地关于微生物岩的报道,多集中在中东部地区奥陶系马家沟组,主要以白云岩为主,分布于马五6、马五1+2、马五7、马五5等亚段,类型主要为叠层石、凝块石、核形石。近期,米探1井在马四段获得工业气流,其中主要储层段岩石为叠层石白云岩,也属于微生物碳酸盐岩。在鄂尔多斯盆地南部,前人关于生物礁的研究较多 [26] [27] [28]。从造礁生物上来看,也主要为藻类,由此可推断微生物岩可能也是鄂南平凉组重要的储集层,但是关于鄂尔多斯盆地南部奥陶纪微生物碳酸盐岩储层目前尚无报道。
本次研究通过对鄂尔多斯盆地南部典型剖面的野外和薄片观察,详细描述了微生物碳酸盐岩岩的岩石学特征和储集空间特征,试图分析鄂南奥陶纪微生物碳酸盐岩的沉积环境,探讨微生物碳酸盐岩成岩过程,为鄂南地区油气勘探提供科学依据。
2. 地质背景
鄂尔多斯盆地是典型的克拉通盆地,位于华北地台西缘,整体划分为六个构造单元:伊盟隆起、西缘复杂构造带、天环拗陷、伊陕斜坡、晋西挠褶带、渭北隆起。盆地南缘奥陶纪沉积时期受秦–祁–贺海槽的控制与古隆起制约,属于台地边缘相–台地前缘斜坡相环境。早奥陶世,鄂南地区持续海退,主要发育泥云坪和云灰坪沉积。中奥陶世,海平面波动频繁,形成了一系列海进海退旋回,主要发育台地缓坡沉积 [29] [30] [31]。晚奥陶世平凉早期,受加里东运动的影响,鄂尔多斯盆地主体部分抬升为陆,南部则继续接受沉积,在中央古隆起的西南缘形成边缘或斜坡相沉积 [32] [33] [34] [35] (图1)。背锅山期,古陆面积进一步增大,仅在西南一隅接受沉积,由开台地边缘–斜坡很快过渡到外侧的深水陆棚。受加里东运动的影响,奥陶纪之后海水整体退出,经历了1.3亿年的沉积间断,直到晚石炭世再次接受沉积。
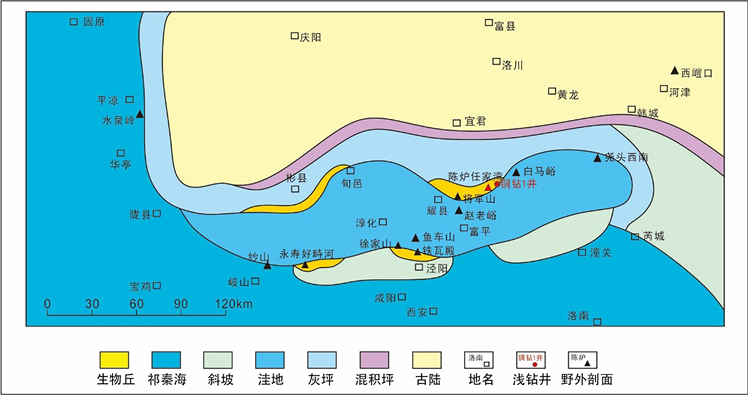
Figure 1. Geographical location map of Renjiawan in the southern Ordos basin [3]
图1. 鄂尔多斯盆地南部任家湾剖面地理位置图 [3]
鄂南陈炉任家湾剖面微生物岩最为发育,位于铜川市陈炉镇附近(图1),属于渭北隆起中心地带。此剖面以平凉组地层为主,未见底,岩性主要以灰色藻灰岩、灰白色砂屑灰岩为主,顶部在平凉组之上可见约20米厚的本溪组铝土岩层。
任家湾剖面中奥陶系平凉组整体呈一个规模较大的生物丘体(图2),由3个砂(砾)屑灰岩—藻灰岩海退旋回组成,反映了海平面频繁变化,逐渐下降的过程。第1旋回,含生屑泥晶灰岩段,含少量藻粘结颗粒和腕足类生物碎屑,层厚约8 m;藻灰岩段,分为上下两部分,下部为藻粘结灰岩,可见大量海绵生物类碎屑,上部为藻纹层灰岩。第2旋回,砂屑灰岩段,毫米级泥晶颗粒磨圆较好,含少量泥晶灰岩角砾,地层厚度为10 m;藻灰岩段,又可分为两部分,上部分为含砂屑和生物碎屑的藻粘结灰岩,厚10 m;下部分为藻纹层灰岩厚3 m。第3旋回,砂屑灰岩段,泥晶质的砂屑颗粒约几毫米,磨圆较好,含少量砾屑,总厚约4 m;藻灰岩段,又细分为三部分,中间一部分为藻纹层灰岩,上下两部分都为藻粘结灰岩,发育较多的海绵碎屑,厚12 m左右;第4旋回,砾屑灰岩段,砾屑含量约80%,棱角明显,大小1~6 cm不等,含生物碎屑、树形石和藻粘结颗粒等,层厚约6 m。
3. 微生物碳酸盐岩特征
微生物碳酸盐岩从规模尺度上可以划分为巨观构造、宏观构造和微观结构三种类型 [36]。巨观构造是大型的生物碳酸盐岩建造,如生物丘、生物礁等;宏观构造是特征尺度较小(一般为几厘米到几米),如穹状、柱状等,微观结构是借助显微镜等工具观察到的现象。通常会以宏观结构特征为依据划分微生物碳酸盐岩的类型,本文分别从宏观和微观两个方面对平凉组微生物碳酸盐岩进行描述划分。

Figure 2. Lithologic columnar diagram of Renjiawan section in the southern Ordos basin
图2. 鄂南陈炉任家湾剖面岩性柱状图
3.1. 微生物碳酸盐岩的宏观特征
任家湾剖面平凉组地层以灰岩为主,仅发育微弱白云岩化。岩石类型主要有凝块石、叠层石、树形石三种微生物碳酸盐岩,其中凝块石最多,叠层石次之,树形石最少。凝块石分布于1/2/3个沉积旋回的藻灰岩段,在砂(砾)屑灰岩段中也少量分布。凝块石一般有两部分构成,一部分是形状不规则的暗色凝块灰岩,另一部分是充填在暗色凝块之间的灰白色方解石胶结物(图3ABC)。凝块颗粒间常出现溶孔,特别是葡萄状花边构造溶孔较为发育,海绵体腔孔也较为发育,非选择性的表生溶孔也较为发育,溶孔形状不规则,从几厘米到几十厘米不等,部分被亮晶方解石充填。凝块石主要发育在潮下带,且发育位置通常比叠层石的位置深 [37] [38]。
叠层石又被称为微生物席,作为微生物群落形成的纹层状沉积构造,由亮层和暗层构成,主要分布于分布于1/2/3个沉积旋回的藻灰岩段。灰黑色的暗层为富藻层,为泥微晶结构,有机质含量高可能与微生物活动存在关联;灰白色亮层,为富屑层,微亮晶结构。藻纹层厚度差别较大(3~10厘米不等),横向延伸可达3米,局部可见丘状突起(图3DE)。叠层石仅发育少量溶蚀孔洞,沿纹层发育,多被亮晶方解石充填(图3F)。叠层石在深度不同的水体条件下均可生长,水平层状叠层石一般在潮间带–潮上带较为发育,丘状叠层石主要出现在潮间带,柱状叠石主要分布在浅潮下带,水体温度和盐度偏高 [39] [40]。
树形石仅见于第4旋回的灰色砾屑灰岩段中,呈“人”字形树枝状,枝体宽度1 mm左右,向外分叉丛生,单簇规模几厘米到几十厘米不等(图3H)。枝干颜色较暗,为灰黑色,周围基质颜色较浅,为灰白色。树形石纵向上具有明显的分段现象。这种具有树枝状的微生物形成的岩石往往被称为树形石 [41],可能由表附藻形成。树形石一般发育在比其他微生物岩更浅的水体环境中,经常暴露的潮间带环境有利于它的发育 [42]。剖面内树形石微生物碳酸盐岩中未见溶蚀孔洞发育。
![]()
Figure 3. Macroscopic tectonic characteristics of the Renjiawan section in the southern Ordos basin
图3. 鄂南任家湾剖面的宏观构造特征
A. 凝块石,葡萄状花边构造溶孔发育,葡萄状花边是由微生物包覆作用形成的,溶蚀孔洞被亮晶方解石充填,有时可见钙化的有机质海绵体发育,体腔内为亮晶方解石充填,第3旋回的藻灰岩段;B. 凝块石,富含有机质的暗色泥晶灰岩呈花斑状分布,花斑之间被乳白色微亮晶方解石胶结物充填,第3旋回藻灰岩段;C. 凝块石,凝块石颗粒之间发育溶蚀洞,充填亮晶方解石,第3旋回藻灰岩段;D. 叠层石,波状纹层,灰黑色和灰白色条纹相间,单层厚度分布不均,横向延伸2~3 m位置,第3旋回藻灰岩段。E. 叠层石,褶皱纹层亮暗条纹间颜色差别较为明显,亮层约厚2 cm,暗层约1 cm,有时在暗色条纹中可见黄色含铁物质,第3旋回藻灰岩段;F. 叠层石,顺层发育在褶皱纹层之间,被亮晶方解石充填,第2旋回藻灰岩段;G. 树形石,表附菌的菌体钙化后发生碎屑沉积,枝体表面沉淀了一层垂直颗粒表面的马牙状亮晶方解石胶结物,基质中较多碎屑颗粒,第4旋回砾屑灰岩段;H. 树形石,表附藻纵剖面上呈明显的分段,段长约1cm的人字形分枝为泥晶灰岩胶结,第4旋回砾屑灰岩段;I. 砾屑灰岩,砾屑颗粒为泥晶质,大小不一,棱角明显,杂乱无章分布,第4旋回砾屑灰岩段。
3.2. 微生物碳酸盐岩的微观特征
微生物碳酸盐岩在岩性上与宏观构造具有一致性,进一步了解微观微生物碳酸盐岩的特征:凝块石在显微镜下观察到微生物种类较多,如蓝绿藻或葛万藻等,形态千差万别,大体上可分为似球粒、团块、斑状等类型。斑状凝块石是藻粘结结构的泥晶颗粒,形状极不规则,大小不一,几百微米左右(图4A)。微生物团块(图4B)是由藻类粘结颗粒聚集形成的具有多个核心或没有核心的复合型颗粒,一般外形不规则,大小约几百微米,多形成于滨浅海碳酸盐岩沉积环境(杨仁超,2013)。似球粒结构为藻粘结颗粒,大小均匀,形态不规则,直径约100 μm左右,结构边界相对模糊,多形成于安静的水体环境,同时观察到致密泥晶中发现有暗色泥晶壁以及基座菌群落(图4C),其形成常与钙化细菌的凝集作用相关 [43]。溶蚀孔洞(图4F)较为发育,形态不规则,毫米级,部分被亮晶方解石充填。宏观剖面中观察到的暗色颗粒物质,在镜下观察为藻类粘结泥晶方解石形成的颗粒,亮色为亮晶方解石形成的胶结物。在微观层面上,凝块石存在的主要储集空间类型有藻粘结颗粒之间的溶孔、葡萄花边状构造溶孔、海绵体腔孔等。葡萄花边构造的溶蚀孔洞较为发育,花边构造主要为纤维柱状等厚环边方解石,一般未被完全充填;海绵体腔孔一般有体腔壁形成的泥晶薄层结构包裹,部分被方解石充填,残余部分孔隙。
叠层石在镜下呈波状或平直的明暗相间的条带(图4I、图4J、图4K),富藻层呈暗色纹层,由有机质含量相对较高的泥晶方解石组成;富屑层呈亮色纹层,由亮晶方解石组成。两层之间发育不规则窗格孔,大都被亮晶方解石充填(图4I、图4K),这些不规则窗格孔沿纹层规则连续分布,被称为“沙漏状(hourglass structures)结构,为蓝细菌微生物席内经过微生物光合作用导致的气泡残留结构 [44]。叠层石的形成一般与微生物的间歇性生长相关,是由蓝绿藻捕获沉积物而产生的微生物席不断叠加形成的,具有生物沉积构造。其中叠层石暗色致密泥晶纹层代表着蓝细菌与光合作用微生物结构的钙化作用有关 [45]。
树形石为暗色致密泥晶颗粒规律排列,整体由似丛生枝状体组成呈扇形,枝状体颗粒之间被亮晶方解石充填,可见发育在其中的肾型菌状的微生物膜钙化产物(图4L)。这种孤立树枝状体由分叉状树形泥晶聚集体组成,直径在2 mm,这被认为是附枝菌菌落(Ephpyon),是树形石中最具有特征性的组构,其本身具有自己的生长格架,常大量生长并与其他的骨骼有机物一起成为造岩生物,一般发育在台地边缘相 [46]。树形石中发育的两种菌落均为钙化蓝细菌化石,其周围展现由微亮晶填充的不规则的孔状结构。在寒武纪、奥陶纪的碳酸盐岩地层中较为常见。显微镜下观察到枝状体之间的亮晶胶结物之间有少量的溶蚀孔洞,内充填亮晶方解石。
A. 凝块石,斑状结构,花斑颗粒由藻粘结颗粒形成,颗粒内部有机质分布不均匀,大小200 μm左右,形状不规则,被微亮晶方解石胶结物胶结,RJW-O2P-50,单偏光;B. 凝块石,微生物团块结构,微生物团块由藻粘结泥晶颗粒被微亮晶胶结而成,RJW-O2P-22,单偏光;C. 凝块石似球粒结构,球粒大小相近,分选较好,主要由藻粘结颗粒和微亮晶胶结物构成,RJW-O2P-37,单偏光;D. 凝块石,颗粒之间发育微亮晶方解石,有沥青充填,RJW-O2P-37,单偏光;E. 凝块石,非选择性表生溶蚀洞,未被充填,RJW-O2P-37,单偏光;F. 凝块石,花边状构造,藻粘结颗粒被两期世代胶结,RJW-O2P-20,单偏光;G. 凝块石,钙化的藻菌体内发生白云石化,RJW-O2P-29,单偏光;H. 凝块石,非选择性溶蚀洞,RJW-O2P-32,单偏光;I. 叠层石,窗格孔被溶蚀后充填颗粒状方解石,RJW-O2P-24,单偏光;J. 叠层石,水平纹层状,亮暗纹层相间,单层厚度比较小,约几十微米,横向延展性好,可见小的窗格孔发育;K. 叠层石,波状纹层状亮暗相间,暗层为富含有机质的微生物泥晶灰岩,亮层为微亮晶灰岩,且顺层发育形状不规则的窗格孔构造,窗格孔内被微亮晶方解石充填;L. 树形石的横切面,富含有机质的泥晶灰岩颗粒排列规律,颗粒周
![]()
Figure 4. Microstructural characteristics of Ordovician rocks in Renjiawan section in the southern Ordos basin
图4. 鄂南陈炉任家湾剖面奥陶系岩石微观结构特征
围被亮晶方解石胶结物充填,RJW-O2P-37,单偏光。
3.3. 微生物碳酸盐岩的C、O同位素特征
鄂尔多斯盆地南部平凉组微生物岩31个样品,δ13CVPDB值与δ18OVPDB值不具有相关性(图5),δ18OVPDB值大于−5‰的样品个数为14个,δ18OVPDB平均值为−5.72‰,说明成岩过程影响小,样品数据能够反映沉积时期海水有效信息。
平凉组的样品中δ13CVPDB值分布范围为−0.6~2.1‰,平均值为1.25‰,普通灰岩δ13CVPDB平均值为0.1‰。奥陶系同期海水中的δ13CVPDB同位素的分布范围为−1~1‰。样品测试数值比正常海水沉积的碳酸盐岩值高,可能是微生物在大量繁殖过程中不断吸收水体中的12C,造成水体中13C的增加,反映发生碳
![]()
Figure 5. Isotopic characteristics of carbon and oxygen in microbial rocks of the Pingliang Formation in the southern Ordos Basin
图5. 鄂尔多斯盆地南部平凉组微生物岩中碳、氧同位素特征
酸盐岩沉积时期,微生物大量发育。样品δ18OVPDB值分布范围为−3‰~−10‰。奥陶系同期海水的δ18OVPDB值分布范围在−6.2‰~−3.7‰之间 [47]。氧同位素相较于同期海水略微偏负,可能是受到加里东期大气淡水淋滤的影响。
4. 讨论
4.1. 发育的沉积环境
在中奥陶纪时期鄂尔多斯南缘处于华北南缘边缘海与秦–祁海槽过渡位置,为浅海–半深海沉积环境。沉积演化由早奥陶纪云坪相沉积,至中–晚奥陶纪台地前缘斜坡相沉积,发育有规模较大的生物礁建造。另外,盆地南缘受构造断裂作用影响,平凉组时期发生垮塌沉积与重力流沉积。在研究剖面的藻粘结灰岩中发现了海绵等造礁生物,同时,在生物礁中含有大量藻粘结灰岩,泾阳铁瓦殿和浅钻铜钻1井之中也见到了造礁生物与凝块石相伴生,进一步说明鄂南奥陶系微生物碳酸盐岩、礁、滩三者是共生关系 [48]。因此,微生物碳酸盐岩在平凉组地层中占主导地位,为优质储层的发育奠定了良好基础。
鄂南陈炉任家湾剖面主要有四种岩石类型,分别是含生屑泥晶灰岩、砾屑灰岩、砂屑灰岩和藻灰岩。含生屑泥晶灰岩为相对低能产物,位于正常浪基面以下;砾屑灰岩指示了台地前缘斜坡相沉积(垮塌沉积),砂屑灰岩多分布在较高能的沉积环境中,藻灰岩形成环境水体能力相对较低,指示了生物丘的沉积特点。这与整体区域处于台地边缘沉积环境相吻合。剖面中不同的微生物碳酸盐岩所反映的沉积环境不同,多发育于水体动能相对较低的环境之中。凝块石主要发育在潮下带沉积环境,水平纹层状叠层石或丘状叠层石主要出现在潮间带–潮上带沉积环境,树形石主要出现在潮间带。剖面上微生物岩种类的变化,也反映了研究区水体的频繁变化,而且微生物碳酸盐岩形成于水动能较弱的水体环境之中。
4.2. 储集空间的形成与演化
从时间上,大致可以将鄂尔多斯南部奥陶系平凉组的微生物碳酸盐岩储集空间演化可为五个时期:1) 准同生或同生阶段形成原生孔隙的时期,储集空间的形成主要受到沉积相的控制以及沉积环境控制,结构性选择作用明显。凝块石的暗色团块内部缺乏孔洞,凝块颗粒间是孔洞发育的主要区域,包括粒间溶孔和葡萄花边状构造溶孔,溶蚀洞周围有微生物形成的花边状构造。海绵体腔孔在海绵体腔内部形成溶蚀孔洞,且海绵体腔壁由于其特征结构形成泥晶化、高有机质的结构;叠层石中的层间溶孔和窗格孔皆形成于微生物纹理层之间;树形石中的针状溶孔也形成于树形微生物体的周围等,这些孔隙一般都与微生物结构相关。2) 与微生物相关的孔隙结构在海水环境下被早期方解石胶结,垂直颗粒边缘形成纤状方解石胶结物,使孔隙度降低。3) 浅埋藏阶段,沉积物被埋藏,在淡水潜流带内发生淡水胶结作用,形成颗粒状的方解石,原生孔隙基本消失。4) 随着沉积物埋藏不断加深,到了深埋藏阶段,由于温度和压力的增加,微生物碳酸盐岩中有机质成熟度升高,经过特裂解等作用产生有机酸,对微生物颗粒周围的胶结物进行结构选择性溶蚀,形成埋藏溶蚀孔洞(可见溶蚀残余)。5) 鄂尔多斯南部后来又经历了加里东构造运动,使鄂尔多斯盆地大规模整体抬升暴露,进入了表生成岩阶段,受大气淡水淋滤影响,在原有残余孔隙的基础上,形成溶蚀孔洞 [49]。
在陈炉任家湾剖面的平凉组微生物岩地层中储集空间主要有:凝块石中的葡萄花边构造溶孔、凝块颗粒间溶孔、海绵体腔孔等;叠层石中主要发育层间溶孔、窗格孔等。这几种类型的储集空间特征均与微生物结构相关,其中葡萄花边构造溶孔最为常见,其与微生物(蓝绿藻)作用相关,海平面下降过程中,藻粘结灰岩受淡水淋滤,富含微生物纹层和叠层构造的泥晶灰岩遭受溶蚀,后期由微生物再次充填沉积形成的 [50]。这种溶蚀洞的形成一般都是选择性溶蚀的结果;在溶蚀作用下,凝块颗粒间的胶结物易被选择性溶蚀,凝块颗粒间的粒间溶孔也较为发育。葡萄花边、凝块颗粒和海绵体腔壁共同形成了藻粘结灰岩中的骨架结构,这种骨架在地层埋藏压实的过程中,保持原来的形态,防止孔隙变小变形,有利于形成储集性能较好的孔隙结构。这种类型的孔隙还为后期的改造提供了基础,后期容易受到埋藏和表生溶蚀改造,形成有效的储集空间。
5. 结论
1) 鄂尔多斯盆地南部的奥陶系平凉组广泛发育有微生物碳酸盐岩。主要发育在台地边缘相,相对低能环境之中,与生物礁、滩相伴生,共同构成了礁、滩、微生物碳酸盐岩组合体。主要有凝块石、叠层石、树形石。沉积相控制了微生物碳酸盐岩的类型。
2) 鄂尔多斯盆地南部奥陶系平凉组微生物岩可作为潜在的碳酸盐岩储层,主要发育在凝块石和叠层石中,储集空间有葡萄花边构造溶孔、凝块颗粒间溶孔、海绵体腔孔、层间溶孔、窗格孔等,受微生物结构、溶蚀作用和充填作用影响。微生物结构为储集空间的发育提供了物质基础,表生溶蚀和埋藏溶蚀增加了储集性能,但充填作用破坏了岩石的储集性能,寻找微生物岩中未充填区是今后探索的主要方向。
基金项目
国家自然科学基金企业创新发展联合基金项目(U19B6003);中国科学院A类战略性先导项目(XDA14010201)。
文章引用
韩月卿,刘 仪,岳小娟,张军涛,金晓辉,韩文彪,赵红琴. 鄂南奥陶系平凉组微生物碳酸盐岩特征与储集意义
The Characteristics and Storage Significance of Microbial Carbonate Rocks in the Ordovician Pingliang Formation of Southern Ordos Basin[J]. 地球科学前沿, 2022, 12(03): 286-297. https://doi.org/10.12677/AG.2022.123030
参考文献
- 1. 李朋威, 罗平, 宋金民, 金廷福, 王果谦. 微生物碳酸盐岩储层特征与主控因素——以塔里木盆地西北缘上震旦统——下寒武统为例[J]. 石油学报, 2015, 36(9): 1074-1089.
- 2. 姚根顺, 郝毅, 周进高, 蒋伟雄, 文龙, 倪超, 等. 四川盆地震旦系灯影组储层储集空间的形成与演化[J]. 天然气工业, 2014, 34(3): 31-37.
- 3. 金晓辉, 张军涛, 孙冬胜, 等. 鄂尔多斯盆地南缘上奥陶统平凉组浅钻中古岩溶洞穴的发现及其意义[J]. 石油与天然气地质2021, 42(3): 595-603.
- 4. 罗平, 王石, 李朋威, 宋金民, 金廷福, 王果谦, 等. 微生物碳酸盐岩油气储层研究现状与展望[J]. 沉积学报, 2013, 31(5): 807-823.
- 5. 施泽进, 梁平, 王勇, 胡修权, 田亚铭, 王长城. 川东南地区灯影组葡萄石地球化学特征及成因分析[J]. 岩石学报, 2011, 27(8): 2263-2271.
- 6. 李朋威, 罗平, 陈敏, 宋金民, 金廷福, 王果谦. 塔里木盆地西北缘上震旦统微生物碳酸盐岩储层特征与成因[J]. 石油与天然气地质, 2015, 36(3): 416-428.
- 7. 包洪平, 姜红霞, 吴亚生, 任军峰, 刘丽静. 鄂尔多斯盆地西南缘陕西陇县晚奥陶世背锅山组生物礁[J]. 微体古生物学报, 2016, 33(2): 152-161.
- 8. 夏明军, 郑聪斌, 毕建霞, 曾正清, 郭海霞, 魏立新, 等. 鄂尔多斯盆地奥陶系生物礁及其天然气勘探前景[J]. 天然气地球科学, 2008, 19(2): 178-182.
- 9. 姜红霞, 包洪平, 孙六一, 吴亚生, 刁建波. 鄂尔多斯盆地南缘奥陶系生物礁的珊瑚化石及其古生态[J]. 古生物学报, 2013, 52(2): 243-255.
- 10. 王文之, 杨跃明, 文龙, 罗冰, 罗文军, 夏茂龙, 等. 微生物碳酸盐岩沉积特征研究——以四川盆地高磨地区灯影组为例[J]. 中国地质, 2016, 43(1): 306-318.
- 11. 魏国齐, 沈平, 杨威, 张健, 焦贵浩, 谢武仁, 等. 四川盆地震旦系大气田形成条件与勘探远景区[J]. 石油勘探与开发, 2013, 40(2): 129-138.
- 12. 刘树根, 宋金民, 罗平, 林彤, 等. 四川盆地深层微生物碳酸盐岩储层特征及其油气勘探前景[J]. 成都理工大学学报: 自然科学版, 2016, 43(2): 129-152.
- 13. Riding, R. (2000) Microbial Carbonates: The Geological Record of Calcified Bacterial-Algal Mats and Biofilms. Sedimentology, 47, 179-214. https://doi.org/10.1046/j.1365-3091.2000.00003.x
- 14. Mancini, E.A., Parcell, W.C., Ahr, W.M., Ramirez, V.O., Llinás, J.C. and Cameron, M. (2008) Upper Jurassic Updip Stratigraphic Trap and Associated Smackover Microbial and Nearshore Carbonate Facies, Eastern Gulf Coastal Plain. AAPG Bulletin, 92, 417-442. https://doi.org/10.1306/11140707076
- 15. Burne, R.V. and Moore, L.S. (1987) Microbialites; Organosedimentary Deposits of Benthic Microbial Communities. Palaios, 2, 241-254. https://doi.org/10.2307/3514674
- 16. 梅冥相. 微生物碳酸盐岩分类体系的修订: 对灰岩成因结构分类体系的补充[J]. 地学前缘, 2007, 14(5): 222-234.
- 17. 梅冥相. 从凝块石概念的演变论微生物碳酸盐岩的研究进展[J]. 地质科技情报, 2007, 26(6): 1-9.
- 18. 许国明, 宋晓波, 冯霞, 隆轲, 王琼仙, 石国山, 等. 川西地区中三叠统雷口坡组天然气勘探潜力[J]. 天然气工业, 2013, 33(8): 8-14.
- 19. 侯方浩, 方少仙, 董兆雄, 赵敬松, 卢蜀秀, 吴诒, 等. 鄂尔多斯盆地中奥陶统马家沟组沉积环境与岩相发育特征[J]. 沉积学报, 2003, 21(1): 106-112.
- 20. 韩作振, 陈吉涛, 迟乃杰, 王兆鹏, 杨仁超, 樊爱萍. 微生物碳酸盐岩研究: 回顾与展望[J]. 海洋地质与第四纪地质, 2009, 29(4): 29-38.
- 21. 邹才能, 杜金虎, 徐春春, 汪泽成, 张宝民, 魏国齐, 等. 四川盆地震旦系-寒武系特大型气田形成分布、资源潜力及勘探发现[J]. 石油勘探与开发, 2014, 41(3): 278-293.
- 22. 郝松立. 鄂尔多斯南缘奥陶纪生物礁及其沉积环境研究[D]: [硕士学位论文]. 西安: 西北大学, 2011.
- 23. 费宝生, 汪建红. 中国海相油气田勘探实例之三渤海湾盆地任丘古潜山大油田的发现与勘探[J]. 海相油气地质, 2005, 10(3): 43-50.
- 24. 方少仙, 侯方浩. 上震旦统灯影组中非叠层石生态系兰细菌白云岩[J]. 沉积学报, 2003, 21(1): 96-105.
- 25. 席胜利, 李振宏, 王欣, 郑聪斌. 鄂尔多斯盆地奥陶系储层展布及勘探潜力[J]. 石油与天然气地质, 2006, 27(3): 405-412.
- 26. 苏中堂, 郝哲敏, 裴文超, 荣浩达. 鄂尔多斯盆地奥陶系生物礁发育特征及其油气地质意义[C]//中国矿物岩石地球化学学会岩相古地理专业委员会、沉积学专业委员会、中国地质学会沉积地质专业委员会、地层古生物专业委员会、煤田地质专业委员会、中国石油学会石油地质专业委员会. 全国古地理学及沉积学学术会议论文集. 2014: 108-109.
- 27. 付金华, 郑聪斌. 鄂尔多斯盆地奥陶纪华北海和祁连海演变及岩相古地理特征[J]. 古地理学报, 2001, 3(4): 25-34.
- 28. 董兆雄, 赵敬松, 方少仙, 侯方浩, 郑聪斌, 章贵松, 等. 鄂尔多斯盆地南部奥陶纪末端变陡缓坡沉积模式[J]. 西南石油大学学报(自然科学版), 2002, 24(1): 50-52.
- 29. 赵振宇, 郭彦如, 王艳, 林冬娟. 鄂尔多斯盆地构造演化及古地理特征研究进展[J]. 特种油气藏, 2012, 19(5): 15-20.
- 30. 冯增昭. 鄂尔多斯地区早古生代相古地理[M]. 北京: 地质出版社, 1991.
- 31. 冯增昭, 彭勇民, 金振奎, 等. 中国寒武纪和奥陶纪岩相古地理[M]. 北京: 石油工业出版社, 2004: 414.
- 32. 王超勇, 陈孟晋, 汪泽成, 郭英海. 鄂尔多斯盆地南部二叠系山西组及下石盒子组盒8段沉积相[J]. 古地理学报, 2007, 9(4): 369-378.
- 33. 李振宏, 胡健民. 鄂尔多斯盆地构造演化与古岩溶储层分布[J]. 石油与天然气地质, 2010, 31(5): 640-647.
- 34. 朱士兴. 中国叠层石[M]. 天津: 天津大学出版社, 1993.
- 35. 彭瀚霖, 刘树根, 宋金民, 孙玮, 丁一, 尹柯惟. 川北米仓山地区灯影组微生物碳酸盐岩发育特征[J]. 成都理工大学学报(自然科学版), 2014, 41(2): 181-191.
- 36. 杨仁超, 樊爱萍, 韩作振, 迟乃杰, 韩豫. 山东寒武系毛庄阶微生物团块的形态特征与成因[J]. 中国科学: 地球科学, 2013, 43(3): 423-432.
- 37. 黄思静, 梁瑞, 黄可可, 兰叶芳. 鄂尔多斯盆地上古生界碎屑岩储层中的鞍形白云石胶结物及其对储层的影响[J]. 成都理工大学学报(自然科学版), 2010, 37(4): 366-376.
- 38. 陈鹤, 巩恩普, 关长庆, 孙宝亮, 张永利. C、O、Sr同位素及微量元素组成在生物礁礁体环境研究中的应用[J]. 世界地质, 2008, 27(2): 130-136.
- 39. 周俊烈, 陈洪德, 赵俊兴, 等. 鄂尔多斯盆地西缘奥陶纪生物礁及其储集性能[J]. 成都理工大学学报(自然科学版), 2013, 40(2): 193-199.
- 40. 赵俊兴, 李凤杰, 朱广社, 苏中堂, 邹敏, 王玉萍, 等. 鄂尔多斯盆地西缘奥陶纪生物礁基本特征、分布规律及成礁模式[J]. 岩石学报, 2014, 30(3): 747-756.
- 41. 胡鑫, 田震, 朱颖, 王文婧. 微生物碳酸盐岩成因分析及其与油气的关系[J]. 天然气技术与经济, 2017, 11(4): 1-4+81
- 42. 施泽进, 王勇, 田亚铭, 王长城. 四川盆地东南部震旦系灯影组藻云岩胶结作用及其成岩流体分析[J]. 中国科学: 地球科学, 2013(2): 317-328.
- 43. Kenter, J.A., Harris, P.M.M. and Porta, G.D. (2004) Microbial and Cement Bound Stone-Dominated Flanks (and Reservoirs) of an Isolated Carbonate Platform. AAPG Annual Convention, Dallas, 18-21 April 2004.
- 44. Kershaw, S., Crasquin, S., Li, Y., Collin, P.Y., Forel, M.B., Mu, X. and Guo, L. (2012) Microbialites and Global Environmental Change across the Permian-Triassic Boundary: A Synthesis. Geobiology, 10, 25-47. https://doi.org/10.1111/j.1472-4669.2011.00302.x
- 45. Bergmann, K.D., Grotzinger, J.P. and Fischer, W.W. (2013) Biological Influences on Seafloor Carbonate Precipitation. Palaios, 28, 99-115. https://doi.org/10.2110/palo.2012.p12-088r
- 46. Warusavitharana, C. and Parcell, W. (2013) Sedimentary Features, Occurrence, and Cyclicity of Microbialite Facies in the Roubidoux and Jefferson City Formations of Missouri and Kansas Arbuckle Group Microbialites in Missouri and Kansas. AAPG Bulletin, 97, 1849-1870. https://doi.org/10.1306/06261312174
- 47. Sarg, J.F., Tänavsuu-Milkeviciene, K. and Humphrey, J.D. (2013) Lithofacies, Stable Isotopic Composition, and Stratigraphic Evolution of Microbial and Associated Carbonates, Green River Formation (Eocene), Piceance Basin, ColoradoMicrobial Carbonates, Green River Formation. AAPG Bulletin, 97, 1937-1966. https://doi.org/10.1306/07031312188
- 48. Kenward, P.A., Fowle, D.A., Goldstein, R.H., Ueshima, M., González, L.A. and Roberts, J.A. (2013) Ordered Low- Temperature Dolomite Mediated by Carboxyl-Group Density of Microbial Cell Walls. AAPG Bulletin, 97, 2113-2125. https://doi.org/10.1306/05171312168
- 49. Wright, P.V. and Racey, A. (2009) Pre-Salt Microbial Carbonate Reservoirs of the Santos Basin, Offshore Brazil. The American Association of Petroleum Geologists Annual Convention and Exhibition, Denver, 7-10 June 2009.
- 50. Riding, R. (1991) Calcareous Algae and Stromatolites. Springer-Verlag, Berlin, 21-51. https://doi.org/10.1007/978-3-642-52335-9