Botanical Research
Vol.3 No.03(2014), Article ID:13621,22 pages
DOI:10.12677/BR.2014.33017
秋海棠(Begonia grandis)的历史文化、 利用、资源多样性和研究进展
Xingjuan Li1,2, Daike Tian2*, Chun Li2, Keming Liu1*, Xiangpeng Li2, Masashi Nakata31School of Life Sciences, Hunan Normal University, Changsha
2Laboratory of Ornamental Plant Resources, Germplasm Innovation and Utilization, Shanghai Chenshan Plant Science Research Center of Chinese Academy of Sciences, Chenshan Botanical Garden, Shanghai
3Botanic Gardens of Toyama, Toyama, Japan
Email: *dktian@sibs.ac.cn
Copyright © 2014 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received: Mar. 6th, 2014; revised: Apr. 3rd, 2014; accepted: Apr. 15th, 2014
ABSTRACT
Begonia grandis Dry. has a long history in cultivation and Chinese literature due to its high values as an ornamental, medicinal, edible and cultural plant. This species is the most northerly distributed, strongest cold-resistant, the only species developing tubers in leaf axils on aerial stems and one of the widest distribution taxa in Begoniaceae. Therefore, it is especially of importance in scientific research. However, the distribution, germplasm and conservation status of this species have not been well understood before, and its intraspecific classification remains questionable. In order to better understand, protect and utilize B. grandis, the history, culture, utilization, classification, distribution, germplasm diversity, intraspecific taxonomical status and research advances on this species are thoroughly investigated, summarized and discussed, through a comprehensive review on literature, specimens in worldwide herbaria, field investigation and the online living plant images of it from the websites of Chinese Natural Herbarium (CFH), Google, etc. The results show that B. grandis is only native to China but has been widely naturalized in Japan. The altitude of its native distribution is from ca. 75 m to 3400 m. The lowest location is at Shanjuandong of Jangsu Prov., and the highest is Haba Snow Mountain of Yunnan Prov. The distribution longitude ranges from 97˚27'34''E to 121˚47'15''E, and the latitude from 22˚59'19''N to 40˚40'05''N. Hekanzi Binggou in Lingyuan County of Liaoning Prov. is the northern boundary, and Pingbian County of Yunnan is the southern boundary; Chawa Long Qin Nathon in Chayu County of Tibet is the most western boundary, and Tiantong National Forest Park in Ningbo of Zhejiang is the eastern boundary. It is for the first time to accurately define the native distribution boundaries and provinces of B. grandis. Comparing with records in Flora of China, six provinces (municipal cities and autonomous regions) were added to its distributions range, namely Chongqing, Guangdong, Jiangsu, Liaoning, Tianjin and Tibet. The wild plants of B. grandis were recently discovered in a mountain of central Taiwan, but further investigation is necessary on its native or naturalized status. It is also found that the intraspecific taxa of B. grandis are not reasonably defined based on Flora Republicae Popularis Sinicae and Flora of China, and the existing criteria don’t work in practice of identification. To solve this issue, we are conducting a comprehensive morphological and molecular study on the intraspecific diversity and scientific classification on B. grandis.
Keywords:Begonia grandis, History, Culture, Germplasm Diversity, Utilization

秋海棠(Begonia grandis)的历史文化、 利用、资源多样性和研究进展
李行娟1,2,田代科2*,李 春2,刘克明1*,李湘鹏2,中田政司3
1湖南师范大学生命科学学院,长沙
2上海辰山植物园/中国科学院上海辰山植物科学研究中心,观赏植物资源及种质创新利用实验室,上海
3日本富山县中央植物园,日本富山市
Email: *dktian@sibs.ac.cn
收稿日期:2014年3月6日;修回日期:2014年4月3日;录用日期:2014年4月15日

摘 要
秋海棠(Begonia grandis Dry.)在我国文献记载和栽培历史悠久,具有较高的观赏、药用、食用和文化价值。该种是世界秋海棠科分布最北端、抗寒性最强及分布范围最广的少数种类之一,也是唯一地上茎叶腋产生珠芽的秋海棠属植物,因此科研价值很高。然而,有关该种的分布、资源及保护现状并不非常清楚,种下分类有待商榷。为更好地了解、保护和利用这一物种资源,本研究通过文献调查、馆藏标本查阅、野外实地考察、中国自然标本馆(CFH)、谷歌等网络活体植物照片考证等综合方法,对该种的历史、文化、分类、资源及分布、利用价值和研究进展进行了全面总结和分析讨论,并对其种下多样性进行了初步探讨。研究表明,秋海棠仅原产于中国,但在日本已经大量野生化。中国原产地分布海拔为75~3400 m,最低点为江苏宜兴善卷洞,最高点为云南哈巴雪山。分布范围为东经97˚27'34''~121˚47'15'',北纬22˚59'19"~40˚40'05'',以辽宁省凌源县河坎子冰沟为最北界,云南屏边县为最南界,西藏察隅县察瓦龙秦那通为最西界,浙江宁波天童国家森林公园为最东界,较“Flora of China”记载相比,增加了重庆、广东、江苏、辽宁、天津和西藏六个省市(自治区)的分布点。近年在台湾中部山区发现该种野生植物,是原生还是自然化结果有待考证。《中国植物志》及“Flora of China”有关该种的种下单元处理均不合理,以现有的划分标准在鉴定实践中无法操作,我们正在对其种下多样性及科学分类开展综合研究。
关键词
秋海棠,历史,文化,资源多样性,应用价值

1. 引言
秋海棠(Begonia grandis Dry.),俗名断肠花、相思草等,隶属于秋海棠属(Begonia L.)、秋海棠科(Begoniaceae),为中国秋海棠科属的代表性种类。作为一种优良的庭园观赏花卉,秋海棠的记载可追溯到我国宋朝,至今已有近千年的栽培历史(据《采兰杂志》)。江户时代宽永年间(1624~1644) 该种首次从中国传入日本,现已在该国多地大量自然化[1] ,在野外开花时形成十分壮观的景象。1804年,William Kerr在中国发现秋海棠并将其引种到英国[2] ,后被各国广泛引种栽培,如作为观赏植物常见于美国诸多植物园及私家庭院[3] 。秋海棠还是我国一种传统中草药,含有多种活性成分[4] [5] ,其药用记载可追溯到1765年赵学敏编著的《本草纲目拾遗》[6] ,距今至少已有250年历史。该种也可食用[7] 。此外,秋海棠还有很高的文化价值,常见于历代文人墨客的诗词歌赋和散文小说中,也见于传统绘画、瓷器和雕刻艺术品。
秋海棠属是植物多样性最丰富的类群之一,目前已知近1600种,为全球第六大属被子植物[8] -[10] ,也是世界著名的观赏花卉。中国的秋海棠属资源十分丰富,仅次于巴西,目前已知187种和若干种下分类群,此外还有不少新类群有待挖掘发表,特别是以往该属考查较少的广西。从最近几年野外调查和新种发表趋势来看,中国野生秋海棠会很容易突破200种[11] 。秋海棠特产我国,是秋海棠属在中国分布最广的种类,也是全球最耐寒、分布最北端和唯一在直立茎上产生珠芽的秋海棠植物。因此,该种不仅具有十分重要的科研价值,还是优良的园艺育种材料。综合文献、标本查阅、野外考察和网络图片等信息,发现该种的种下多样性十分丰富,现有的种下分类混乱而不确定,资源状况及其保护现状的评估也是空白,均有待进一步研究。
为了更好地了解、合理利用和保护这一重要物种资源,本研究通过全面系统的文献调查、馆藏标本查阅、野外实地考察、网络野生植物照片及其产地考证等综合方法,对秋海棠的历史文化,分类,资源状况,观赏、药用、文化等利用价值以及各方面研究进展进行了深入分析和总结,并对其种下单元分类及多样性问题进行了初步探讨。本文中单独使用“秋海棠”一词时专指Begonia grandis及种下类群统称,特此说明。
2. 秋海棠的历史与文化艺术
2.1. 秋海棠的历史记载和早期生物学描述
西方最早关于秋海棠属的记载是从1651年西班牙僧侶Francisco Hernandez在墨西哥所绘的一种球茎秋海棠开始[12] 。直到1777年,活体秋海棠属植物开始从牙买加引种到欧洲进行栽培。1695年Plumier首次将来自加勒比海岛屿上的6种植物定名为Begonia,此后林奈将此6种合并为1种,并定名为Begonia obliqua L.,即秋海棠属的模式种[13] 。
秋海棠在我国的记载历史十分悠久,可追溯到宋朝的《采兰杂志》(作者不详,960~1279年),距今已有约千年,比其属的历史要早很多。由于历史长、分布广、多样性高,该种有多个名称,如断肠花(《采兰杂志》)、八月春(《花镜》)、相思草(《本草纲目拾遗》)、断肠草(《大观录》)、白海棠(《红楼梦》)、岩丸子(《中华本草》)、璎珞草等。其中,断肠草的名字可能起源于断肠花,为后者别名,但也可能同有毒植物的断肠草名称混淆误用。
有关秋海棠的生长习性及生物学性状在古代文献中就多有记载,如《采兰杂志》中描述了秋海棠的栽培环境及叶花等部分特征:“昔有妇人怀人不见,恒洒泪于北墙之下。后洒处生草,其花甚媚,色如妇面,其叶正绿反红,秋开,名曰断肠花,即今秋海棠也”;于若瀛(1552~1610)所著的《弗告堂集》对其生长习性、茎和叶脉、花序着花数等性状进行了清晰描述:“秋海棠喜阴生,又宜卑湿,茎岐处作浅绛,色绿,叶文似朱丝,婉媚可人,不独花也”;清朝闵梦得《万历葵丑漳州府志》(约1613年):“秋海棠岁每生苗,其茎甚脆,早秋始花,略似海棠半含,浅红可爱”,描述秋海棠地上部分为一年生、茎脆弱、早秋开花,花似蔷薇科苹果属半开粉红色的海棠花;王象晋《二如亭群芳谱》(约1621年)及汪灏(约1708年)《广群芳谱》:“一名八月春,草本,花色粉红,甚娇艳,叶绿如翠羽。此花有二种,叶下红筋者为常品,绿筋者开花更有雅趣”,指出该种叶脉有两种颜色类型;王穉登《荆溪疏》(约1583年):“善卷后洞秋时,海棠千本并著花,一壑皆丹”,描述了秋海棠喜生于溶洞旁或内,成片生长、红花盛开的壮观景象。善卷后洞位于江苏省宜兴市张渚镇善卷洞风景区,善卷洞景区分上中下三洞,经笔者实地考证,惟下洞瀑布旁岩石上长有一片秋海棠。
《学圃余疏》(曾棨,1587)中则首次记载了秋海棠的种植方法及观赏应用:“秋海棠娇好,宜于幽砌,北牕下种之,傍以古拙一峰,菖蒲、翠筠草皆其益友也”。《瓶花谱》(张丑,1595)中记载:“《花经九命升降》,吾家先哲(君讳翌)所制,可谓缩万象于笔端,实幻景于片楮矣,今谱瓶花,例当列品,录其入供者,得数十种,亦以九品九命次第之”,其中,秋海棠、山矾、夜合、赛兰、蔷薇、锦葵、杏、辛夷、各色千叶榴、佛桑、梨同为四品六命;《瓶史》(袁宏道,1599)中记载:“秋海棠娇,然有酸态,郑康成、崔秀才之侍儿也”。由此可见,秋海棠早在16世纪末就已在中国作为插花用。
清陈淏子《花镜》(1688年)卷六《秋海棠》对该种形态特征、种下多样性、生长习性、物候期和栽培管理要领等进行了全面详细的记载:“秋海棠一名八月春,为秋色中第一,本矮而叶大,背多红丝如胭脂,作界纹,花四出,以渐而开,至末朵结铃,子生桠枝,花之娇冶柔媚,真同美人倦粧。性喜阴湿,多见日色即瘁,九月收子。撒于盆内或墙下,明春自发。但老根过冬,则花更盛,不必浇肥。其异种有黄、白二色。俗传昔有女子,怀人不至,涕泪洒地,遂生此花,故色娇如女面,名为断肠花。若花谢结子后即剪去,来年花发叶稀而盛。冬亦畏冷,地上须堆以艸盖之。独昌州、定州海棠有香,诚异品也”。但是,文中提到的黄色品种存疑,因目前还未发现该种有黄花类型;最后一句指出昌州(重庆辖)、定州(河北辖)产的香花海棠并非秋海棠,我们判断其应是苹果属的海棠类(Malus Mill.),因为古时昌州号称海棠香国,最早出自南宋著名地理学家王象之《舆地纪胜》里的《静南志》。
当然,由于时代局限性和植物分类学知识的限制,中国古代文献有关秋海棠的记载肯定存在一些错误。如清关涵辑《岭南随笔》(1795年):“海棠本无香,惟清远归猿洞秋海棠、肇庆羊峡春海棠,其香特盛”中提到清远的秋海棠很可能为另种,肇庆羊峡的春海棠可能是香花秋海棠(B. handelii Irmsch.)或紫背天葵(B. fimbristipula Hance),因为后两者均在春季开花,远早于秋天开花的秋海棠,具体何种有待于进一步考证。又如:清吴升《大观录》(1712年)记载:“秋海棠亦名断肠草,其根、叶有毒,犬马食之即死,浸花水饮之害人”,名字很可能被张冠李戴,被误作马钱科的金钩吻(Gelsemium sempervirens (L.) J.St.-Hil.)、卫矛科的雷公藤(Tripterygium wilfordii Hook. f.)等某种强毒植物,这些种类也叫断肠草,而秋海棠尽管富含草酸,但低毒可食,不大量摄取则无碍。但是,秋海棠的毒性究竟多大,值得未来研究。
秋海棠在中国的药用价值记载则始于1765年的《本草纲目拾遗》:“海棠喜背阴而生,故性寒,凡大热症可用”[6] 。
据日本文献记载,秋海棠原产中国,在江户时代宽永年间(1624~1644)首次由中国传入日本[1] 。该种现已在日本广泛栽培,且很多地方早已自然化,大量成片生长,盛花时景象十分美丽壮观,如日本栃木县出流町的流ふれあいの森、埼玉县ときがわ町的椚平等成为观赏该种的著名景点。1804年,英国的William Kerr在中国发现秋海棠并将其引种到英国[2] 。随后,欧美各国不断从中国、日本引种,广泛栽培于私家庭院、植物园和公园中,并发现一些变异类型,通过直接筛选和人工杂交培育了系列品种(见7.6育种)。
尽管秋海棠在中国的历史十分悠久,分布范围广、资源丰富,且为历代文人墨客佳作的重要题材,但国内对其资源的开发利用还远不够。目前,除了部分寺庙外,国内植物园、公园和私家庭院中很少见,远不及日本、欧美少数国家栽培广泛,我国也尚未培育出一个该种起源的品种。
2.2. 秋海棠的文化艺术
2.2.1. 文化起源
作为一种观赏价值很高的庭院花卉和传统名贵草药,秋海棠的文化同其利用紧密相连、同步发展,常作为历代散文、诗词、绘画、雕刻等重要文化艺术题材。宋朝的《采兰杂志》就将秋海棠比喻成别离伤感之花,对该种的描述在《琅嬛记》(伊世珍,年代不详)及《广群芳谱》(汪灏等,约1708年)中均有引用:“昔有妇人怀人不见,恒洒泪于北墙之下。后洒处生草,其花甚媚,色如妇面,其叶正绿反红,秋开,名曰断肠花,即今秋海棠也”。
2.2.2. 诗词
秋海棠更多成为明、清文人骚客们诗歌中的意象。明代陈道复(1483~1544)的《题秋海棠》算是较早的咏秋海棠诗:“翠叶纷披花满枝,风前袅袅学低垂。墙根昨日开无数,谁说秋来少艳姿”;明王士骐(生卒年不详)题秋海棠:“是花偏灼灼,开处几丛丛,弱质不禁露,幽怀欲诉风,空庭聊取媚,傍石若为容,黄菊纷相应,餐英未许同”;明钟惺(1574~1624)咏秋海棠:“墻壁固吾分,烟霜亦是恩,光轻偏到蒂,命薄幸余根,笑泣谁能喻,荣衰不敢论,年年秋色下,幽独自相存”;明俞琬纶(生卒年不详)咏秋海棠:“薄罗初试怯风凄,小样红妆着雨低,一假妖娆描不就,非关子美不能诗。春色先应到海棠,独留此种占秋芳,稀疏点缀猩红小,堪佐黄花荐客觞”;清纳兰性德(1655~1685)《锦堂春ž秋海棠》:“帘外澹烟一缕,墙阴几簇低花。夜来微雨西风里,无力在欹斜。仿佛个人睡起,晕红不著铅华。天寒翠袖添凄楚,愁近欲栖鸦”及《临江仙ž塞上得家报云秋海棠开矣赋此》:“六曲阑干三夜雨,倩谁护取娇慵。可怜寂寞粉墙东。已分裙衩绿,犹裹泪绡红。曾记鬓边斜落下,半床凉月惺忪。旧欢如在梦魂中。自然肠欲断,何必更秋风”;清尤怡(?~1749)咏白秋海棠:“谁将清泪洒幽墀?散作瑶华别有姿。最是玉人断肠后,淡妆无语背人时”;清朱受心(生卒年不详)《白秋海棠》:“清秋湛露浥琼芳,素影风摇玉砌旁。夜静看花人独立,水晶帘外月如霜”;清黄景仁(1749~1783)《午窗偶成》:“只有断肠花一种,墙根愁雨复愁风”;清钱枚(1761~1803)《卖花声ž从友人乞秋海棠》:“微雨暗西楼,花事句留。从君分取半庭秋。属付园丁轻著力,牢护根头。花韵便盈眸,无限温柔。问花真个断肠不?侬做断肠人已惯,翻替花愁”。
与大多数其它花木相比,秋海棠的娇小柔媚和断肠花的别名,正契合了旧时代女性的性格特征和社会角色,因此也更受女性青睐。明代杨宛(?~1644)为金陵妓女,有《浪淘沙·海棠》词:“尽日若含愁,别样娇羞。晚凉香散上帘钩。带露摘来斜插鬓,一段风流。蛩语玉阶幽,又是深秋。相携闲对小妆楼。不解断肠伊似我,我似伊不?”;清代女诗人何佩芬(生卒年不详)《秋海棠用渔洋山人秋柳四首韵》之四:“绿肥红瘦总堪怜,泪洒胭脂欲化烟。不占春光仍烂漫,纵无秋思也缠绵。静含清露刚三日,艳绝西风又一年。翻羡紫薇高格好,株株占得凤池边”;清代女诗人赵韵卿(1814~1894)《诉衷情·病起看秋海棠》:“碧梧庭院暑初收,凉意逗衣篝。为忆海棠开未,呼婢卷帘钩。扶薄病,怯惊秋,强凝眸。西风萧瑟,花怜人瘦,人比花愁”。然而,鉴湖女侠秋瑾(1875~1907)赋予了秋海棠全新的意义:“栽植恩深雨露同,一丛浅淡一丛浓。平生不借春光力,几度开来斗晚风?”,表现了巾帼英豪不同凡响的英雄气概。清朝文学家袁枚(1716~1797)则以秋海棠的品格自道,表白归隐故园是在于保持自己的品格和晚节,《秋海棠》一诗咏曰:“小朵娇红窈窕姿,独含秋气发花迟。暗中自有清香在,不是幽人不得知”。关于秋海棠的传说,还有一段凄美的爱情故事。南宋爱国诗人陆游(1125~1210)一生苦恋唐婉,但唐婉家境贫寒,终不能为陆游母亲所容,最终有情人终不能成眷属。临别之际,唐琬赠送一盆秋海棠给陆游,并告诉他这是“断肠花”。陆游一听更伤心,说此花应称为“相思花”。后来陆游故地重游,触景伤情,作《秋海棠》诗一首:“横陈锦彤栏杆外,尽收红云洒盏中。贪看不辞持夜烛,倚狂直欲擅春风”。
在众多咏秋海棠诗歌中,曹雪芹的别有特色。在《红楼梦》第三十七回,大观园里成立海棠诗社,结社后的第一次活动是咏白海棠,薛宝钗、林黛玉、贾探春、贾宝玉各依所限韵脚做七言律诗一首。湘云凭《白海棠和韵》:“神仙昨日降都门,种得蓝田玉一盆。自是霜娥偏爱冷,非关倩女欲离魂。秋阴捧出何方雪,雨渍添来隔宿痕。却喜诗人吟不倦,肯令寂寞度朝昏”和“蘅芷阶通萝薜门,也宜墙角也宜盆。花因喜洁难寻偶,人为悲秋易断魂。玉烛滴干风里泪,晶帘隔破月中痕。幽情欲向嫦娥诉,无奈虚廊夜色昏”二首压倒众人。其诗用神仙种玉喻白海棠(即秋海棠白花品种),丰富的联想中,突显湘云与海棠的紧密联系[14] [15] 。
2.2.3. 戏曲
1941年,秦瘦鸥发表同名小说《秋海棠》,故事主人公秋海棠摈弃了科班中师傅为他所取的极女性化的艺名吴玉琴,将艺名改为秋海棠,取意“中国的地形,整个儿连起来恰像一片秋海棠叶”。
歌曲则分别有《秋海棠叶》(1982年倪伟德词、施光南曲)和《啊,秋海棠》(1987、史俊词,吕其明曲)、《秋海棠》(2010,苏一一词,多人作曲)等。
2.2.4. 绘画、瓷器等
秋海棠也是历代画家的创作题材之一。齐白石、张大千、吴昌硕、李苦禅等大师留下很多秋海棠名画,十分名贵,价值十万百万的不在少数,如中国嘉德国际拍卖有限公司拍卖的齐白石戊寅(1938年)的画作《秋海棠行书七言诗》成扇纸本以149.5万元成交。秋海棠题材也见于历代瓷器、油票和歌曲。瓷器中有清朝的“粉彩秋海棠小碟”到现代的茅台酒酒瓶。秋海棠在台湾常指“秋海棠地图”,即包括外蒙古等在内的中华民国地图或1947年前出版的中国地图,因其形状酷似一片近对称的秋海棠叶而得名,并非现在的公鸡状。实际上,中华民国成立前,清朝时期的中国地图外形就似秋海棠叶。
受中国文化的影响,秋海棠引种到日本后,有关该种的文化也得到萌发和发展,以秋海棠为题材的日本诗词和瓷器等也不少见。
3. 秋海棠的分类
Begonia grandis由Dryander于1791年发表,即现在广泛接受的秋海棠学名,模式标本采自日本[16] 。B. grandis Otto[17] 和B. grandis Otto ex A. D.C.[18] 皆为晚出同名而无效。
《中国植物志》52卷将国产秋海棠属划分为6个组:侧膜胎座组(Sect. Coeloecentrum)、秋海棠组(Sect. Begonia)、单座组(Sect. Reichenheimia)、四室组(Sect. Sphenanthera)、多室组(Sect. Pleiothece)、二室组(Sect. Platycentrum),秋海棠被划分到秋海棠组[19] 。而Flora of China (FOC)将国产种类划为7个组,即在《中国植物志》基础上增加了二室单座组(Sect. Monolobium),但秋海棠的归属组没变[20] 。国产秋海棠属的分组还存在一定异议,如《云南植物志》第12卷(2006)将其分为8个组,即侧膜组、小花组(Sect. Alicida)、东亚秋海棠组(Sect. Diploclinium)、单座组、小海棠组(Sect. Parvibegonia)、扁果组(Sect. Platycentrum)、棒果组(Sect. Leprosae)及无翅组(Sect. Sphenanthera),秋海棠则被划归到东亚秋海棠组[21] 。根据Doorenbos等[12] 和Shui等[22] ,秋海棠组(Sect. Begonia)在亚洲并不存在,秋海棠应属于东亚秋海棠组(Sect. Diploclinium)。
《中国植物志》[19] 将秋海棠处理为一个原亚种:秋海棠(B. grandis subsp. grandis);两个亚种:全柱秋海棠(B. grandis subsp. holostyla Irmsch.)、中华秋海棠(B. grandis subsp. sinensis (A. D.C.) Irmsch.);三个变种:单翅秋海棠(B. grandis var. unialata Irmsch.),刺毛中华秋海棠(B. grandis var. puberula Irmsch.)和柔毛中华秋海棠(B. grandis var. villosa T.C. Ku)。而FOC[20] 仅承认1原亚种、2亚种,即秋海棠(B. grandis subsp. grandis)、全柱秋海棠(B. grandis subsp. holostyla Irmsch.)及中华秋海棠(B. grandis subsp. sinensis (A. D.C.) Irmsch.),对该种分类检索如下:
1a) 花柱分离,不分支;柱头头状或肾状;雄蕊柱短于1 mm。
……全柱秋海棠B. grandis subsp. holostyla 1b) 花柱基部合生或者微合生,分支;柱头螺旋状扭曲或者U字形;雄蕊多数,雄蕊柱长于1 mm。
2a) 植株较细弱;叶背面绿色,偶尔略带红色,椭圆状卵形至三角状卵形,5~12(~20) × 3.5~9 (~13) cm;雄蕊柱短于2 mm。
……中华秋海棠B. grandis subsp. sinensis 2b) 植株较强壮;叶背面红色或至少叶脉红色,卵形至宽卵形,叶片大于中华秋海棠;雄蕊柱超过2 mm。
……秋海棠B. grandis subsp. grandis
然而,根据《中国植物志》和FOC分类标准,在实践中无法准确地对秋海棠的种下单元进行分类,即使对一株具备全部重要性状的个体开展鉴定。在考察全国各地馆藏标本时也发现该种的鉴定十分混乱,同一地区、同一地点、甚至同号的标本往往被鉴定为不同亚种。通过系统的野外调查和结合中国自然标本馆(CFH)中该种的大量野生植物照片比对后发现:除云南局部所产全柱秋海棠(B. grandis subsp. holostyla)的柱头不分支、成头状,这一特征同相邻及其它地区的居群明显不同外,按现有的亚种分类标准,全国其它地方分布的类群根本无法分开。例如,从分布最北端的辽宁凌源县到我国南部的广东阳山县、西南部的广西凤山县、云南麻栗坡县,植株均产生珠芽,同一居群有时兼有分枝和不分枝、叶背紫红和绿色二类个体,有的兼有叶背紫红、仅背脉紫红和叶背全绿3种类型植株,性状变异较大,雄蕊柱长短并无严格界限,因此,种下分类边界尚不明确,需要更全面深入地研究。尽管发现产云南麻栗坡县中越边境的居群个体较小、雄蕊数较少,但其余形态学特征同全国其他各地产的居群差异不大。当前,我们以形态学为基础,结合分子生物学手段正开展其种下多样性研究,以评估《中国植物志》和FOC有关秋海棠亚种划分的合理性,希望提出新的种下分类处理及标准,前期分子生物学研究所得结果(另行发表)印证了上述观点。
4. 秋海棠的种质资源
4.1. 种下多样性
《中国植物志》52卷将秋海棠处理为一个原亚种,两个亚种,三个变种。而FOC仅承认1原亚种、2亚种。鉴于种下单元划分还不明确和科学,暂将该种及种下类群统称为秋海棠(B. grandis),文中也如此处理。但从性状上看,其多样性比较丰富,表现在:1) 植株有大小及茎分枝多少差异(图1:A1~A3);2) 叶片明显存在背面绿色、叶肉绿色叶脉紫红、叶背面紫红3种个体类型,还有一些过渡形态,这些类型见于不同居群,有时也存在同一居群内(图1:C4~C5);3) 叶表面有绿色无斑、白斑两种类型(图1:A1~A4,C1~C3),后者少见,且斑纹多少及形态存在一定差异,但在湖南永顺县的一个居群,数公里范围内的植株叶片斑纹几乎表现一致(图1:A4,C3),说明该类个体在此地形成了稳定的遗传体系,是否作为一个新变型或变种,正在开展研究;4) 花通常为粉红色花,不同产地的居群花色深浅有一定差异,白花或近白花个体少见(《红楼梦》中也有提及)(图1:D1~D4);5) 雄蕊数多少、雄蕊柱长短、雌蕊特征等变化较大(图1:D1~D3,E1~E3),其界限有待于进一步系统调查研究;6) 因分布范围广和海拔不同,秋海棠花期差别大,为6~9月,南方早北方迟;7) 果翅,尤其是不同居群长翅的形态及大小差异大、变化丰富(图1:B1~B3),是否用于种下分类的重要标准也有待考证。
4.2. 资源及分布
秋海棠原产中国,但其具体分布范围、资源及保护状况还不很清楚。基于这一点,我们对世界各国馆藏的该种标本进行了全面收集整理,标本馆包括国际著名的英国丘园(K)、美国密苏里植物园(MO)、

Figure 1. Diversity of Begonia grandis; (A1)-(A3) Plants nonbranching to highly branching; (A4) Individual with white spotted leaf; (B1)-(B3) Shape of fruits and wings: (B1) Very short lateral wing; (B2) Longer lateral wing, abaxial wing nearly vertical to fruit; (B3) Curved abaxial wing; (C1)-(C3) Leaf shape and edge; (C4)-(C5) Diversity of abaxial leaf color, C4 is from the same population; (D1)-(D4) Difference in male flower: (D1) White flower individual; (D2)-(D4) Filaments connected 10 to 2 mm; (E1)-(E3) Difference in stigma: highly developed to simply head-shaped
图1. 秋海棠的种内多样性;(A1)-(A3) 植株由不分枝到分枝多;(A4) 白斑叶个体;(B1)-(B3) 果实及果翅形态差异:(B1) 侧翅极短;(B2) 侧翅较长、背翅近垂直果轴;(B3) 背翅明显内弯;(C1)-(C3) 叶由不裂具细齿到小齿再到浅裂粗齿;(C4)-(C5) 叶背颜色差异,其中C4表示同一居群不同个体间差异;(D1)-(D4) 雄花差异:(D1) 白花个体较少;(D2)-(D4) 为粉红花,且雄蕊柱长度由最长约1 cm到2毫米;(E1)-(E3) 雌花柱头由扭曲十分发达到头状不发达
美国国立自然历史博物馆(US)、德国自然博物馆(B)、中科院植物所(PE)、昆明植物所(KUN)、华南植物园(IBSC)、武汉植物园(HIB)等国际著名标本馆到非常小的研究机构(如国内的食品药品检验所)。到目前为止,共考察了全世界有该种标本的87个标本馆,得到1286号、2209多份标本(包括我们近3年采集的87号,319份),预计占该种所有标本的90%以上。另外,无采集号的标本有66份。
尽管秋海棠的标本采集量大,为国产秋海棠属馆藏标本量最大的种类,且有分布的地方大多有标本,但平均每号标本份数不到2,备份太少。馆藏量超过或接近100份的标本馆分别是中科院植物所(477)、四川大学(230)、华南植物园(138)、昆明植物所(115)和西北农林科技大学(96),由此可知,该种标本量总体上同馆藏单位以前植物分类科研力量成正相关(图2)。
该种标本的采集量同资源分布丰富度也大体成正相关,分布市县多的省份标本采集量也大。其中,四川分布该种的市县最多,达53个,采集量也最大,共采集标本231号480份;云南、湖北、湖南次之,都有29个市县分布,分别采集149号290份、149号248份、121号186份(图2)。采集时间段主要集中在20世纪50年代至60年代,其次为30年代至50年代。这一结果与中国的植物标本采集史相关:辛亥革命前,我国专门从事植物分类研究的本国学者寥若晨星,大规模的植物标本采集活动仅限于外国人;辛亥革命后,随着我国植物分类学研究队伍的成长和壮大,我国学者便开始了对本土植物标本的采集活动。尤其是1949年中华人民共和国成立后,植物分类学研究受到国家的高度重视,国人才真正开始独立地对本土植物资源进行大规模调查和采集。采集地点主要集中在四川、云南、湖北等地[23] 。
同时,结合标本和文献信息,我们还对全国21个省市及自治区50余县开展了野外实地考察,采集了一大批腊叶标本和分子材料,再综合中国自然标本馆(CFH)、中国数字植物标本馆(CVH)、百度、谷歌
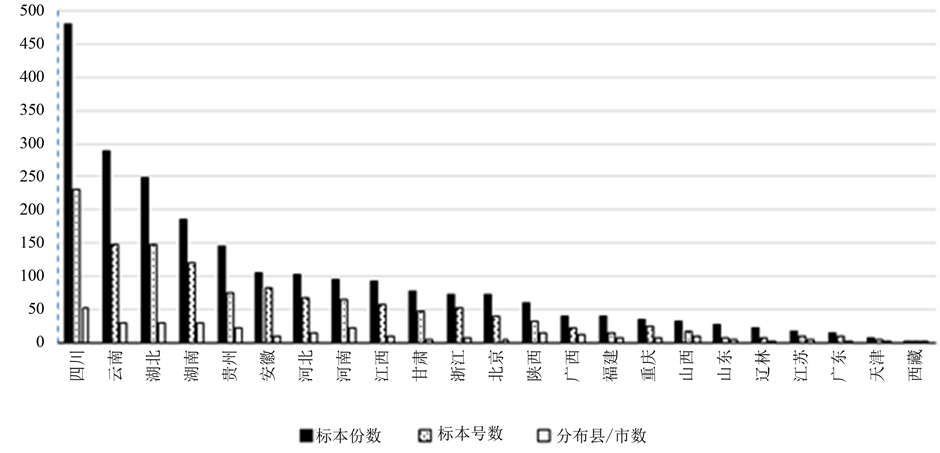
Figure 2. Native distribution of Begonia grandis based on specimens
图2. 秋海棠标本产地分布
等网站的秋海棠图片考证,不仅对原有产地记载但无标本信息的分布点进行确认,也获得一批新分布地点数据,最终得出目前最完整、最准确的秋海棠分布图(图3)。结果表明:秋海棠(B. grandis incl. subsp.)仅产中国大陆,分布海拔为75~3400 m,最低分布点为江苏宜兴张渚镇善卷洞风景区,最高为云南哈巴雪山。以浙江宁波天童国家森林公园(121˚47'15.572''E, 29˚48'9.378''N)为东界;西藏察隅县察瓦龙秦那通(97˚27'34.981''E, 28˚41'8.765''N)是西界;云南屏边县(103˚41'40.049''E, 22˚59'19.453''N)为南界;辽宁省凌源县河坎子冰沟(119˚16'14.707''E, 40˚40'5.923''N)为北界,也是秋海棠属全球分布的最北端。第一次比较准确地弄清了该种的分布界限及分布省份、市县,较FOC记载,增加了6个省市(自治区)级分布点:重庆(金佛山,城口东安乡、莱蒙区、旗杆山、后坪乡,石柱沙子镇,巫山竹贤乡,巫溪中梁乡、白果乡,秀山茅坡乡,奉节双店乡,酉阳木叶乡)、广东(阳山寨岗镇、乳源通天箩)、江苏(宜兴张渚镇,连云港云台山,江宁淳化乡)、天津(蓟县黄崖关雁东坡、八仙山)、辽宁(凌源河坎子)、西藏(察隅察瓦龙)(图3)。六省市的标本信息如下。
Chongqing(重庆):Jinfoshan(金佛山),X. L. Zhou(周喜乐) TDK639 (CSH);Chengkou(城口),Bashan Exp. (巴山采集队) 1045 (PE),T. L. Dai(戴天伦) 104348 (PE, SZ)、102478 (PE)、104107 (PE, SZ)、105908 (PE);Shizhu(石柱),W. H. Wang(王文华) 1974 (SZ);Wushan(巫山),H. F. Zhou(周洪富) 110060 (PE);Wuxi(巫溪),G. H. Yang(杨光辉) 65255 (PE, CDCM)、59113 (CDCM),Sichuan Univ.(四川大学) 59113 (PE);Xiushan (秀山),Z. Y. Yan(严铸云) 94039 (CDCM);Fengjie(奉节),H. F. Zhou et al.(周洪富等) 109233 (PE, SZ),M. Y. Fang(方明渊) 24869 (PE);Youyang(酉阳),Z. Y. Liu(刘正宇) 7016 (PE)。Guangdong(广东):Yangshan (阳山),D. K. Tian & L. Z. Tian(田代科、田礼志) TDK983 (CSH),L. Deng(邓良) 262 (IBSC)、1678 (IBSC)。Jiangsu(江苏):Yixing(宜兴),X. J. Li et al.(李行娟等) lxj028 (CSH),W. Z. Fang(方文哲等) 292 (PE);Lianyungan(连云港),F. X. Liu(刘昉勋) 10912 (PE);Jiangning(江宁),Y. Huang(黄宜) 6310 (N)。Liaoning (辽宁):Lingyuan(凌源),D. K. Tian & Y. Chen(田代科、陈岳) TDK987 (CSH)、TDK1110 (CSH)、TDK1115 (CSH)、TDK1117 (CSH),Z. F. Fang(方振富) 2961 (IFP),M. D. Liu(刘明德) 128 (IFP),S. X. Li(李书心) 5074 (IFP)。Tianjing (天津):Jixian(蓟县),K. Tian & Y. Chen(田代科、陈岳) TDK1171、TDK1172 (CSH),Anonymous(无名氏) 17209 (BNU)。Xizang(西藏):Chayu(察隅),Q. W. Wang(王启无) 65262 (PE)。
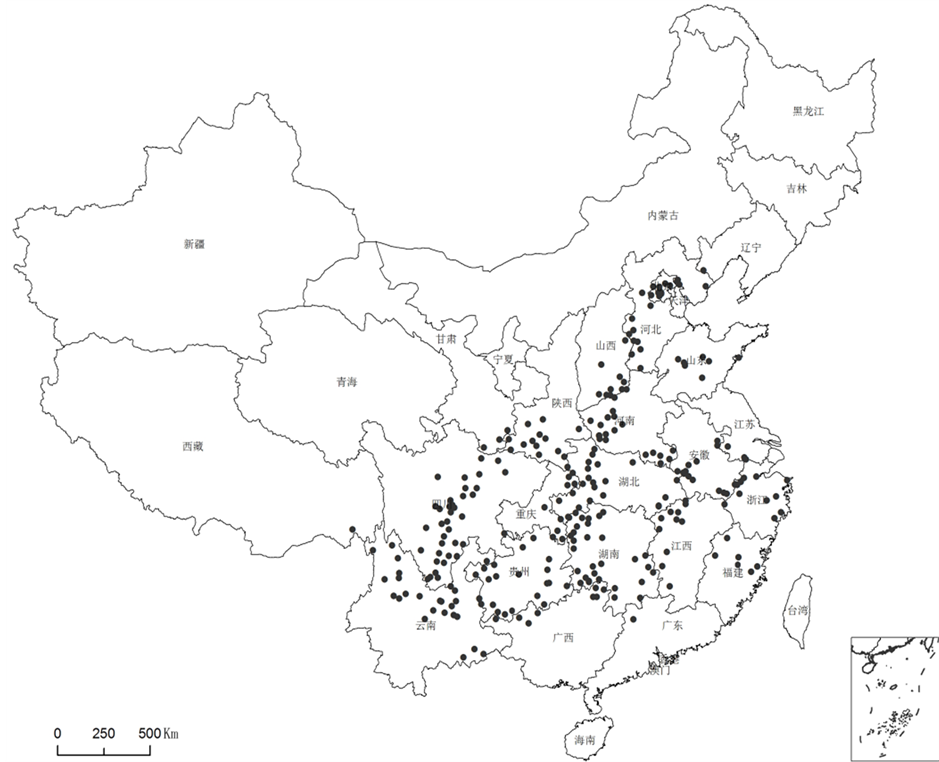
Figure 3. The distribution of Begonia grandis in China; Jiangsu is recorded in Flora Republicae Popularis Sinicae but lacked in Flora of China
图3. 秋海棠在中国大陆的分布图;《中国植物志》记载江苏有分布,但Flora of China中缺
秋海棠作为中国乃至世界秋海棠属的一个广布种,也是唯一分布于秦岭以北的耐寒种类。根据美国农业部(USDA)的标准,该种耐寒等级属6~9区,即可耐−20℃严寒(同其最北端分布点辽宁凌源县的冬季最低温相近),因此为秋海棠属抗低温最强的种类。野外考察和栽培实践发现秋海棠也较耐旱,对环境的适应性比较强。
此外,通过日本研究文献[1] 、日本语版谷歌等网上图片调查考证,我们还对秋海棠在日本的野生化状况进行了分析统计,得出该种至少在日本21个县(都、府)存在野生化及半野生化现象(图4):包括关东地区的东京、栃木、群马、埼玉、神奈川、千叶;近畿地区的滋贺、兵库、大阪、京都;中部地区的静冈、富山、岐阜、新泻、冈山;四国地方的德岛、爱媛、高知;九州的福冈;东北地区的岩手、宮城。主要集中在日本的中部,特别是栃木县辖属的都幾川町椚平、佐野市熊鹰山等多地野生化面积大,长势良好,秋季开花时形成十分美丽的自然景观,成为日本著名的秋海棠观赏景点。
秋海棠在日本栽培很广,野生化程度高,相反在我国庭园中少见,除了寺庙周边外,野外多半见于植被保持良好的保护区和风景区内,很少见明显的自然化迹象。最近,我国台湾中部山区南端一处发现有该种野生植物,并成功引种栽培,可惜原生境已被一泥石流破坏(同台湾植物学家彭镜毅交流),因此,此处该种是原产还是自然化结果,有待考证。
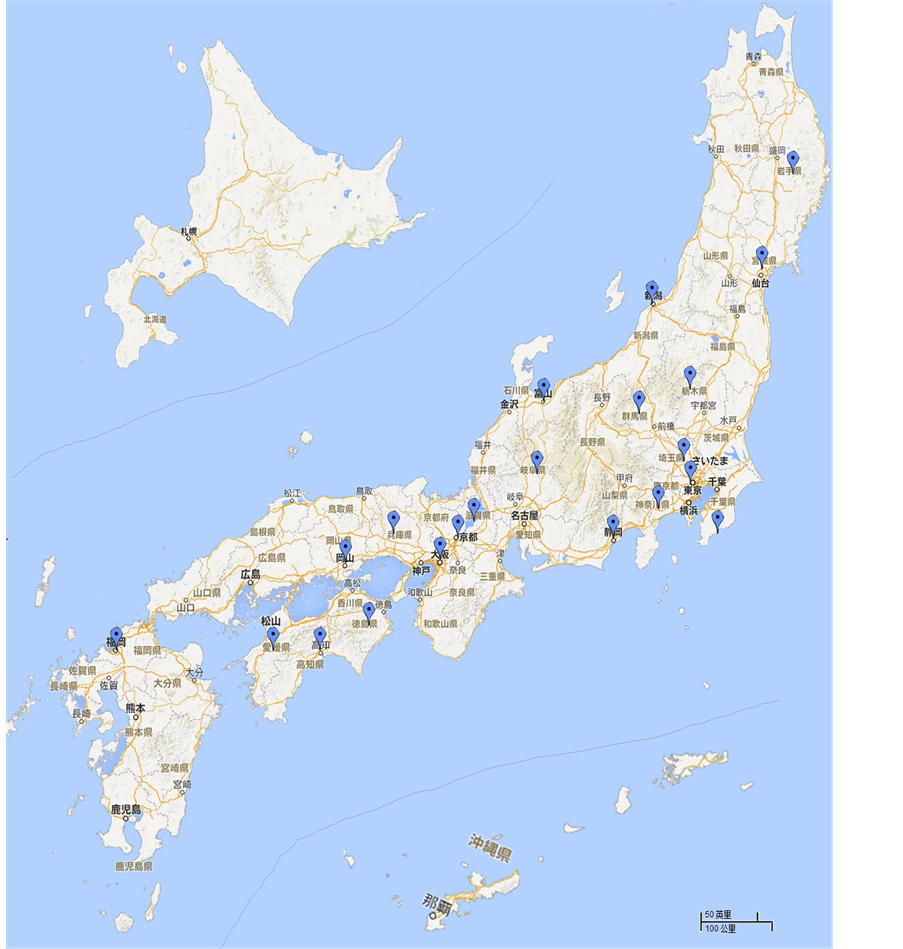
Figure 4. The distribution of naturalized B. grandis in Japan
图4. 秋海棠(B. grandis)在日本自然野生化分布图
5. 秋海棠的繁殖及散布
5.1. 秋海棠的繁殖
秋海棠常片状群生于小溪沟两侧石壁、斜坡、瀑布边、石缝、石穴、洞穴内以及人工堆砌的石墙缝等,这些地方要么植被通常良好,要么处于狭窄的峡谷,生境特殊,每天只能接受到部分日照甚至只有散射光(如溶洞内和林下),周围环境湿度适宜,土壤春夏湿润,秋冬多干燥。该种在中国、日本的寺庙周围常见栽培。
在自然界,秋海棠主要通过种子和珠芽两种方式实现自我繁殖,以块茎的增殖为补充。种子一般在8月后才逐渐成熟,随着气温变凉、植株变黄枯萎,果实变干燥形成裂缝,种子在风力或动物接触等外
力作用下掉落地上。一旦环境适宜,来年春季种子就会萌发生长。赵宝林等[24] 实验表明,中华秋海棠的种子萌发率为80%左右,不受基质类型影响,但种子萌发需要光,在暗室条件下不能萌发。
另一方面,秋海棠植株几乎每个叶腋处都常生长几个到10多个珠芽,这些珠芽随植株成熟衰老而脱离母体,一旦安全越冬,在来年春季雨季来临时同样可萌发、发育成完整植株。在野外调查过程中发现,种子和珠芽自然繁殖形成的幼苗几乎每个居群可见。秋海棠也可通过增加块茎的数量实现增殖,随着新球茎在主球茎上形成、发育成新植株,主球茎的水分和营养不断转移到新个体而最终自然衰亡[25] 。
除以上几种繁殖方式外,还可通过分株、扦插和组培等人工繁殖[24] 。但是,分株受数量限制,叶片和茎插方式更可取。作者开展了该种的叶片繁殖,发现再生十分容易。如用珍珠岩做基质,在室温条件下,扦插10天后左右长根,30天左右分化出芽,40天左右长出小叶。组培方面,赵宝林等[24] 利用采自北京植物园的中华秋海棠为材料,摸索出最佳消毒时间和培养基配方,建立了该种的组培繁殖体系。
5.2. 秋海棠的散布
秋海棠属种类十分丰富,繁殖和传播方式存在一定差别。通过大量野外考察、结合种类特征和栽培实践,可以总结出该属植物大体有5种散布或传播方式:重力、风力、水力、动物及人为传播。重力传播是指种子或珠芽脱离母体后,借自身的重力垂直落下,随生长地坡势向下方移动一定距离,繁殖成新个体后逐渐扩大居群大小和面积范围。风力传播:尽管秋海棠属的种子相对较大,且无翅,不能随风飘走,但风力对种子的散布有一定的作用,对蒴果种类,相对静止情况,风力作用会使正在掉落的种子或带翅果实飘离母株一定距离,从而对扩大居群范围起到一定效果,而对于不开裂的浆果类则无作用。水力传播:秋海棠属的很多种类生长于溪流和瀑布边或其两侧,或陡坡、石壁,其果实、种子甚至营养器官都可能被溪流和雨水带到下游,萌发生长,从而扩大了繁殖生长范围。动物传播:一方面,当动物经过干燥成熟的秋海棠果实时,通过身体的接触,细小的种子掉在动物身上被带到另外一个地方,遇到适宜的条件萌发生长;另一方面,在野外常发现被动物啃食的浆果类秋海棠果实,其种子也有可能被动物带到其他地方发育成植株。人为传播:该种方式尽管对秋海棠属的散布距离影响最大,但往往只对有重要利用价值的种类产生影响,特别是一些观赏或药用价值高的种类,通过栽培利用扩大分布范围,甚至导致逸出自然化生长(如四季秋海棠B. cucullata Willd.、秋海棠)。
秋海棠具有以上所有5种传播方式,其中动物和人为传播方式更有利于该种的远距离传播,这与动物和人的生活习性关系密切。一些四足动物可以将该种种子带到离母体较远的洞穴、石穴,这些地方往往比较阴湿,有利于种子萌发,适合秋海棠生长;而对人类来说,秋海棠不仅是一种观赏花卉,也是一种药材,被僧人或寺庙工作人员引种栽培到寺庙的概率很大,而寺庙周边的良好环境往往给该种的长期定居、逸生和种群扩散奠定了基础,这点可以部分解释为何我国及日本著名风景名胜区内的寺庙周边常见该种的缘故。
6. 秋海棠的应用价值
秋海棠不仅具有很高的观赏价值,还有一定的药用、食用和文化价值。
6.1. 观赏
秋海棠属植物以其优美的株形、奇特叶形、多样叶色及斑纹、艳丽花色成了集观花、观形、观叶、观茎及观果等提供欣赏多样性状的优良花卉,不仅适合于温室栽培,还可室外栽培观赏。全世界目前已培育出至少1万个品种,是很受欢迎的观赏植物[26] -[28] 。最著名的有四季秋海棠(B. cucullata Willd.)及其系列品种、球茎秋海棠(B. × tuberhybrida Voss)、丽格海棠(B. × hiemalis Fotsch.)等,以及观叶为主的大王秋海棠系列品种(Rex Begonia)。其中,四季秋海棠应用最广、量最大,为世界第6大地被植物,而球
茎海棠和丽格海棠主要以盆栽观赏为主。美国、加拿大、澳大利亚等国秋海棠协会每年召开年会,同时举行秋海棠品种及栽培技术展示评比;比利时布鲁塞尔每年举行一次规模宏大的秋海棠花地毯节,在广场上用秋海棠等装点成花纹“地毯”展示其美丽,吸引大量游客参观欣赏。
秋海棠的观赏价值早在一千多年前的《采兰杂志》就曾记载。从我国历代文献中也可得知,该种在全国栽培广泛,且存在少见的白花品种。目前,本种在世界各国的植物园、公园和私家庭院随处可见。秋海棠在日本多地大量自然化,形成美丽自然景观,成为秋季赏花的重要对象之一。欧美国家通过直接筛选和人工杂交培育出20多个品种(见7.6育种),根据网络记载,就有上百家园艺企业销售原种及其部分品种种球。但是,我国既没培育出一个该种相关品种,也不见有品种栽培,栽培的秋海棠基本上源于山中野草。相对美国和日本广泛栽培利用而言,该种在中国仅多见于寺庙前后,而植物园、公园、药草园、私家园林和居民家中少见栽培,因此,其观赏价值没有得到充分利用。
6.2. 药用
秋海棠属植物大部分都具有一定的药用价值,常作为民间中草药[29] -[31] ,部分种类还可以作为食物、饮料和饲料使用[29] [32] 。
秋海棠在中国的药用历史悠久,其药用记载始于1765年赵学敏编著的《本草纲目拾遗》[6] 。关于该种药用记载的书籍很多,《陆川本草》(作者及出书年代不详)、《脉药联珠药性考》(龙柏,1795)、《陕西中草药》[33] 、《贵州民间方药集》[34] 、湖南药用植物资源[35] 、《秦岭巴山天然药物志》[36] 等均有记载。秋海棠全草,包括根、茎、叶、花和果实均可入药,以根为主,指其块茎部分,又名:红白二丸、岩丸子、鸳鸯七、红黑二丸、一口血等。味苦、酸、涩,性微寒。功能有活血调经,止血止痢,镇痛。主治崩漏,月经不调,赤白带下,外伤出血,痢疾,胃痛,腹痛,腰痛,疝气痛,痛经以及跌打瘀痛等。内服:煎汤,6~15克;研末或泡酒。外用:适量,捣敷。秋海棠茎叶,味酸、辛,性微寒。具有解毒消肿,散瘀止痛,杀虫的功效。主治咽喉肿痛,疮痈溃疡,毒蛇咬伤,跌打散瘀,皮癣。外用:适量,鲜品捣敷或绞汁含漱。秋海棠花,味苦、酸,性寒。具有杀虫解毒作用,主治皮癣。外用:适量,捣汁调蜜抹。秋海棠果,味酸、涩、微辛,性凉。具有消肿解毒作用,主治毒蛇咬伤。外用:鲜品适量,捣敷或者捣汁搽[29] 。
附方有:
1) 治跌打损伤,伤后吐血:一口血、强盗绞杆子根各3克,根一钱,研成细末,用酒吞服[34] 。
2) 治痢疾:红白二丸二钱,水煎,加红白糖服[33] 。
3) 治月家病:一口血适量,用童便泡七天后,研末,每次二钱,蒸鸡吃[37] 。
4) 治崩漏,白带:红白二丸、石泽兰各二钱。水煎服[33] 。
5) 治淋浊白浊:白秋海棠块根研末,开水送服一钱,血尿用红秋海棠块根研末,开水吞服二钱[38] 。
6) 治咽喉疼痛:秋海棠根四两,加冷开水两小碗,捣烂,取汁含漱数次[34] 。
除了药用价值外,秋海棠也可食用[30] ,生食可生津解渴,但由于植株富含草酸,低毒,不宜多食。
6.3. 文化价值
秋海棠的文化价值同其观赏、药用价值息息相关。正是由于该种具有很高的观赏和药用价值,加上资源分布广、为秋海棠属最耐寒种类,被历代名人百姓所喜爱,广泛栽培,经常作为散文、诗词、绘画、雕刻等文化艺术的重要题材(见2.2秋海棠的文化艺术)。
7. 秋海棠的研究进展
有关秋海棠属的研究以往主要集中在传统的资源调查、新分类群发表、形态学分类、引种驯化和新品种培育等方面。近几年来,利用分子生物学手段从种、组、属的水平研究遗传分化、多样性式样、系统进化等逐渐增多[12] [39] -[43] ,但在种下水平的研究鲜有报道[44] 。国内外很少涉及秋海棠及其种下单位的究,仅在形态学[25] 、细胞学[1] [45] [46] 以及化学成分分析上[4] [47] -[50] 有少量报道,缺乏系统性和全面性,没有利用现代分子生物学手段研究的报道。
7.1. 分类学
秋海棠的学名“Begonia grandis”最早由Dryander于1791年发表在Transactions of the Linnean Society of London 1: 163,后来不断发表多个亚种、变种、变型,由于这些发表都是建立在简单的形态观察,甚至仅仅依据几份腊叶标本,难免导致分类不清、产生诸多异名。我们发现该种的种下分类至今还十分混乱,原有划分标准不科学,在鉴定上缺乏实践操作性。因此,有关秋海棠的分类问题主要集中在其种下划分。
《中国植物志》将该种按1种,2亚种,3变种处理,而FOC对其进行修正,分类单元减少为1种,2亚种,0变种,但缺乏足够的科学依据。因为,该修订处理并没有建立在系统科学研究的基础上,尤其是没有开展馆藏标本的全面收集鉴定和野外实地考察,更没有利用形态学和分子生物学方法对种下多样性开展研究。因此,其分类标准不科学,按FOC的分类处理,仍然无法对秋海棠种下类群进行合理鉴定和科学划分。当前,我们对全世界该种的绝大部分馆藏标本进行了收集整理,并且以居群为单位对该种在我国各地的野生分布点开展了全面调查,采集了87号300余份标本,一千余份分子样品,并将所有考察点的居群引种栽培,综合形态学和分子生物学手段进行研究,以期解决其种下分类难题,更好地理解秋海棠的种下多样性、性状遗传及演化规律。
7.2. 形态学
除了原有分类群发表文献外,针对秋海棠的形态学研究十分匮乏,仅有少量片段破碎化报道。马宏[25] 在攻读博士学位期间,曾对中国秋海棠属东亚秋海棠组(sect Diploclinium)植物的系统学与保护生物学进行了研究,调查了该种的花粉、叶表皮和毛被特征(图5)。指出:秋海棠的花粉为超长球形,平均大小为18.4 × 6.4 µm,均P/E值为2.7;萌发孔类型为三孔沟;表面纹饰为长条纹明显突起成棱,近平行,两极处交叉,无穿孔,中央处不明显缢缩;孔沟处纹饰为条纹变浅或无,萌发孔处不明显下陷。这说明该种的花粉与所属秋海棠组、单裂组和二室组的其它种类不同,而与四室组的种类高度相近,反映出秋海棠组可能通过秋海棠(或亦包括其它未研究的种类)与四室组相联系。秋海棠和少瓣秋海棠(B. wangii Yü)种皮垂周壁明显加厚,这与二室组的裂叶秋海棠(B. palmata D.Don)、赤水秋海棠(B. chishuiensis Ku)和截裂秋海棠(B. miranda Irmsch.)相似,说明秋海棠组也可能通过秋海棠和少瓣秋海棠与二室组联系,部分支持税玉民等的观点[51] 。秋海棠叶片下表皮细胞呈不规则形;垂周壁式样为平直、弯弓形;气孔器类型为不等型;气孔指数23.66;气孔大小42.79 µm;拥有3~4个副卫细胞,弓形;单列毛、多列毛偶有分叉。其副卫或保卫细胞覆有蜡质结晶体,可能与降低蒸腾作用,减少体内水分散失有关,从而适应夏季干燥或一些向阳的环境,如林缘、路边等。
7.3. 细胞学
自1936年以来,有关秋海棠属植物染色体的研究报道较多[1] [45] [46] [52] -[61] 。秋海棠染色体,最早由Matsuura和Okuno[45] 在1936年报道,2n = 26。为了探讨日本栽培及野生化秋海棠的来源、是否出现分化,Nakata等[1] 对日本12个地区18个地点的秋海棠 (包括栽培品种B. grandis ‘Alba’)以及中国云南玉溪、昆明和移栽于昆明植物所的四川雅安的3个来源秋海棠个体的染色体进行了研究,结果表明其染色体数目均为2n = 26。这与之前Matsuura & Okuno[45] 、White等[53] (B. evansiana = B. grandis)、
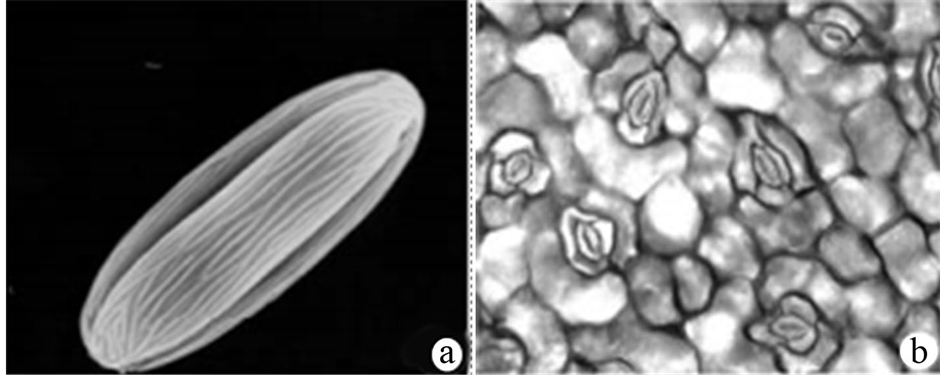
Figure 5. (a) Pollen morphology of Begonia grandis; (b) Characters of abaxial surface of leaf in B. grandis [25]
图5. (a) 秋海棠花粉形态;(b) 秋海棠叶片下表皮光学形态[25]
Legro & Doorenbos[55] (B. grandis var. evansiana = B. grandis)等研究结果相同。但是Bowden[62] 报道秋海棠(B. evansiana = B. grandis)染色体数目为2n = 24,实为计数错误。秋海棠属至今没有一个确定的染色体基数[28] 。Matsuura和Okuno[45] [52] 指出6、7和13为其染色体基数,13是一个二次起源,秋海棠(2n = 26)可能是由两个原始染色体基数为n = 6和7的祖先基因型衍生而来的异源多倍体。这一观点得到Okuno和Nagai[46] 的支持。Dewitte等[63] 研究指出秋海棠属植物基因组大小与染色体数目没有显著的相关性。因此,我们将利用流式细胞仪技术探讨秋海棠种下单位的基因组大小差异,看能否为探讨其种下单元分类及性状演化规律提供部分证据。此外,该种不同居群或种下单元的染色体形态和核型是否也存在差异,为分类提供证据,也有待进一步研究。
7.4. 化学成分
国内外对秋海棠属的化学成分研究很少,仅涉及少数几个种,如作为著名凉茶材料的紫背天葵[32] 、蕺叶秋海棠(B. limprichtii Irmsch) [64] 和秋海棠,其中对秋海棠研究最多。
国内对于秋海棠的植物化学研究主要集中于化学成分(甾醇类化合物)的分析和提取。秋海棠含有多种活性成分,包括有机酸、皂苷、黄酮、生物碱、强心苷、内酯、三萜类成分和甾醇等。植物甾醇是一种三萜醇类化合物,是秋海棠的活性成分之一[47] 。常见的植物甾醇提取方法有溶剂结晶法、络合法、皂化法[65] 、蒸馏法[66] 、柱吸附法、超临界CO2萃取法[67] 、酶法[68] 、超声波法[47] 等。秋海棠所含甾醇类物质有四种,其中β-谷甾醇含量最高,达43.42%;其次是豆甾醇,1.68%;然后是豆甾-5,24(28)-双烯-3β-醇,11.23%;麦角甾-5-烯-3β-醇,9.13% [4] [5] [47] -[50] 。秋海棠还有较强的抗大肠杆菌活性[69] 。秋海棠甾醇类物质对金黄色葡萄球菌、枯草芽孢杆菌、大肠杆菌、沙门氏菌、志贺氏菌、黑曲霉和橘青霉均有一定的抑制效果[48] ,但秋海棠的毒性问题没有进行过研究评估。
7.5. 繁殖与栽培生理
秋海棠极易繁殖,利用种子、珠芽、分根、扦插、组培均可[24] 。该种常栽培于具有散射光的半荫环境,极适合用作阴生地被植物。土壤要求湿润,富含腐殖质,微酸性(pH 6.0~6.8),排水良好,需水量应均衡而规律,不要积水[70] 。该种也耐一定的干旱和低温,冬季休眠期可耐零下20度低温。
秋海棠种子发芽的光周期习性与典型长日照开花植物相同,在29℃条件下,至少经过2~3次8 h长日照才萌芽;赤霉素可明显降低种子发芽光周期的临界值,促进萌发,但在暗条件下无作用[71] 。播种基质对种子萌发无显著影响,不同基质中发芽率均在80%以上[24] 。秋海棠在生长季节中后期几乎每一叶柄基部都会产生3~8个(或更多)珠芽,珠芽散落在母本周围,度过冬季休眠期后于次年4~6月萌发长成新植株[25] 。我们也开展了秋海棠的叶片扦插繁殖试验,发现叶片繁殖也很容易:如用珍珠岩做扦插基质,室温条件下10天左右长根,30天左右分化出芽,40天左右长出小叶。组培方面,赵宝林等[24] 利用采自北京植物园的中华秋海棠为材料,摸索出最佳消毒时间和培养基配方,建立了其组培繁殖体系。
秋海棠在当年或多年生的球茎上能直接生成新球茎,随着新球茎形成,主球茎的水分和营养不断转移到子球茎中去,并最终自然干瘪衰亡[25] 。有关该种珠芽的形成、休眠、萌发条件日本学者研究较多。珠芽形成分三个时期:夜萌芽时期——在这个时期珠芽萌发可不需光;光萌芽时期——这个时期需光;热萌芽时期——需低温[72] 。珠芽形成常是短日照的应答结果,但是当一部分光周期维持低温时,长日照条件下也会形成;如果植株在光照条件下预先接受低温处理,珠芽可在连续黑暗中形成,珠芽大小随温度降低或低温处理时间延长而增加;如果植株经过低温处理,珠芽在单个的光周期反应中也可形成,所需临界夜长低于标准值,珠芽随低温时间延长而变大;没有低温处理的植株,珠芽形成至少需要两个短日照周期;老龄植物对单个短日照无应答[73] 。在暗周期,相对低温和高温都会抑制珠芽的形成,最佳的夜温度是23℃。暗周期有两个热敏阶段:第一个阶段在暗周期开始的时候,第二个阶段则因光周期的长度不同而异,低温对这两个阶段都有影响,而高温只对后阶段有影响[74] 。
秋海棠珠芽的萌发受珠芽成熟度、光照、温度等条件影响。不成熟珠芽在光照条件下可萌芽,而成熟珠芽必须经低温处理、打破休眠后才可萌芽[75] 。氧气是珠芽萌发必须的,也是在2℃~5℃春化过程中打破珠芽休眠所需。在蓝光下,低到3%浓度的氧气条件即可显著打破成熟珠芽的休眠,但在更低浓度氧条件下才促进未成熟珠芽发芽。当氧张力增加时,红光可诱导未成熟珠芽休眠并延长成熟珠芽休眠,黑暗条件下17℃低温处理也可诱导不成熟珠芽休眠[76] 。光质和温度对珠芽休眠有明显调控作用。蓝光和远红外光诱导光萌芽并促进打破经低温(2℃~5℃)处理的成熟珠芽休眠,但在室温条件下不能打破成熟珠芽的休眠,相反15℃~17℃条件下促进不成熟珠芽休眠;红光抑制未成熟珠芽的光萌芽,但在低温下打破成熟珠芽休眠[75] 。红光抑制珠芽生长,从而刺激植物生长。蓝光和远红外光也抑制珠芽生长,但很少刺激植物生长,远红外光在黑暗条件下抑制珠芽形成[77] 。在暗周期之前或暗周期照射近红外光,将抑制珠芽形成[78] 。长日照和短日照对秋海棠珠芽的形成和萌芽起拮抗作用,对一片叶子的短日照处理会被对另一片叶子的长日照处理抑制,不管位置是上还是下,幼叶更明显,而短日照对幼叶的作用很小[79] 。红光对减少光周期反应最有效,在低强度下几乎完全抑制块茎的形成,12小时光周期下,随后经蓝光和远红外照射可逆,8 h光周期下则不可逆[80] 。
脱落酸(3 × 10−6至10−5 M)可抑制秋海棠珠芽萌芽,但一些实验表明10−6 M脱落酸促进萌芽,脱落酸在长日照条件下诱导珠芽形成无效[81] 。赤霉素处理后,导致可溶性乙酸乙酯和正丁醇等生长抑制剂增加,从而抑制珠芽萌芽,当抑制剂积累到一定程度,珠芽休眠[82] 。在高氧浓度下,赤霉素对珠芽萌芽的抑制作用加强,在低氧浓度下,其抑制减弱;对硝基酚、间苯二酚、水杨醛肟、2,4-二硝基苯酚、叠氮化钠和环己酰亚胺可降低赤霉素对珠芽萌芽的抑制作用[83] 。吲哚乙酸、萘乙酸和吲哚丁酸在暗周期处理叶片,可抑制珠芽形成[84] ;2,4-二硝基苯酚、对硝基苯酚、叠氮化纳抑制珠芽休眠[76] 。
吲哚乙酸氧化酶在短日照(SD)处理两天后达到最大值,当SD处理被夜间断,6天后达到最大值,该酶与吲哚乙酸呈负相关[85] ,表明短日照处理通过影响珠芽内部激素水平而影响休眠和萌发。在SD处理开始到第10天,赤霉素(GA3)显著抑制珠芽形成,并促进发育中的珠芽萌发,但是10天后,GA3加速并延长珠芽休眠,GA3对分离的珠芽休眠促进作用随其成熟度增加而增强[86] 。形态素与细胞分裂素一样,促进成熟和未成熟的珠芽萌芽,但与后者不同,形态素不刺激珠芽变大,可打破赤霉素对珠芽萌芽

Table 1. Cultivars from Begonia grandis and its hybrids
表1. 秋海棠的栽培品种
*Begonia grandis subsp. evansiana按Flora of China处理应为Begonia grandis subsp. grandis,这里仍按RHS的处理,作为独立亚种;中文名释根据英文品种名翻译而来;“-”表示信息不详。的抑制作用,促进顶端优势[87] 。
Ahn和Park[88] 指出:组培时期开始就用Cu2+预处理的秋海棠根部能积累大量Cu2+,表明秋海棠有较强的Cu2+耐受力和积累力,所以,该种可用于Cu2+富集的水土环境修复。
7.6. 育种
作为秋海棠属唯一耐寒性种类,秋海棠在西方常被称为耐寒秋海棠(hardy begonia)。加上观赏价值高、种下多样性丰富、历史悠久,秋海棠是一种不可多得的优良育种材料。在国外,由于对秋海棠属育种和观赏应用高度重视,经过一百多年努力,已培育出品种过万。据相关文献[89] [90] 、美国秋海棠协会数据库(ABS Database)[91] 及英国皇家园艺学会数据库(RHS Database)[92] 等资料记载统计,秋海棠相关的品种至少就有23个(表1)。为了培育耐寒性品种,Speese[93] 在美国佛吉尼亚的Charlottesville将产自中国的樟木秋海棠(B. picta Smith)同秋海棠杂交,但杂交后代并不能在此地过冬。
在我国,仅有中科院昆明植物所从上世纪90年代后期开始秋海棠属育种,已培育出20多个品种[94] -[98] 。李景秀等[99] 利用秋海棠与变色秋海棠(B. versicolor,要求空气湿度80%以上)进行杂交,F1植株能够在较低空气湿度(65%左右)环境条件下正常生长,具备一定的适应能力。但是,无论通过直接选育或杂交手段,国内至今无一个源于秋海棠原种及其杂交后代的品种,栽培的该种均来自其野生植物。因此,秋海棠在我国的育种及观赏利用还有待加强。
8. 总结与讨论
我们的研究结果表明:秋海棠的馆藏标本数量大,但鉴定较混乱,同一地点的相同或相近居群标本常被鉴定为不同亚种,显然同实际不符;《中国植物志》和FOC对其种下单元分类处理并不科学,即使利用有叶、花、果等全部重要形态特征的植株来对照鉴定,也无法确定到亚种,说明现有的种下分类依据标准不适用,在实践中无法操作,有待进一步深入研究。
尽管本文提出了一个目前最完整、准确的秋海棠分布图,但由于还有少量标本的采集信息不全或缺失,如南昌大学、江西中医药大学、北京林业大学等一些标本馆所藏该种标本的相当一部分无采集地点记录,因此无法考证具体分布地点;中科院昆明植物所标本馆(KUN)中一份条形码为0533127的复印件标本,数据库中记录为采集号41094,C. Wang采自东沙群岛,采集地点实为抄录错误。因为经我们考证,中科院广西植物所标本馆(IBK)一份条形码为IBK00117401的标本(采集号:41094,采集人:黄志C. Wang)为同号复份标本,采集地点为广西南丹县,但被错误鉴定为裂叶秋海棠(B. palmata D. Don);再者,凭现有分布范围、海拔和气候条件等推断,南沙群岛不会有秋海棠分布。此外,未调查的甘肃、东北三省、西藏等国内和国外部分标本馆可能有少量秋海棠腊叶标本,还在收集中。因此,县级单位及以下分布点有待进一步统计核实和精确化。
秋海棠仅原产中国,但很多国外文献记载该种也产马来西亚,实为错误。鉴于秋海棠在我国分布范围广、种下变异大,要更好地把握其种下分类单位界限及关键区别性状,除了对该种馆藏标本尽可能全面收集和考证外,还要结合大量野外考察和引种栽培观察,才能获取更全面、准确的形态及生理学信息。同时,必须借助细胞学、分子生物学等综合手段开展深入研究,才可望解决其种下分类、多样性及其演化规律难题。当前,作者课题组正在对该种的多样性及种下分类开展系统全面的研究,已对秋海棠全国分布范围内的的20个省(市、自治区)44个居群进行取样,采集了大量标本和分子材料,并进行引种栽培。计划在形态学调查基础上,调查染色体数目和核型特征,并利用流式细胞仪分析其基因组大小和利用分子生物学方法研究居群多样性,综合分析探讨秋海棠种下亲缘关系和遗传多样性,为种下单位的正确划分提供科学依据(结果待发表)。
从全国范围看,秋海棠是一个广布种,在大多数自然保护区、森林公园、风景名胜区等多有分布、保护良好,保护区和风景区外一些地方也可见。这表明,相对同属其它种类,该种适应性强、抗逆境能力高,能适应一定程度的生态环境改变。因此,较小的生态环境破坏并不一定导致其生存危机[23] 。但该种对所栖息地的小生境要求很高,即对水、相对湿度、光(或郁闭度)和低温均有一定的范围要求,所以通常呈现片段化零星分布,有时一座山仅有一两小处生长,且个体数量少,长期以来居群没有扩大,如分布在浙江平阳山门满田森林公园的白岩寺庙旁的秋海棠成株还不过3株。极端分布点的类群性状往往出现明显分化,如产云南丽江的全柱秋海棠亚种,居群柱头不分支、不扭曲、呈头状;分布于麻栗坡中越边界的秋海棠表现为个体小、雄蕊数少等特征;辽宁凌源和西藏察隅的居群处于极端分布点,地理位具有特殊性,个体小,抗寒性最高;此外,一些特殊居群,如产湘西永顺的白斑叶类群,性状十分稳定等,比较合适作为一个新变种或变型,从遗传多样性保护角度,这些类群该优先重点保护。
有关秋海棠的科学问题,还有许多值得进一步研究回答。例如:该种是否有必要再下分亚种、变种,如需要,如何细分,依据和关键区别性状是什么?种内多样性规律及特点、分布中心位置、如何分化和迁移扩散的?日本自然化居群的多样性程度及其可能来源(同中国哪些分布类群亲缘关系最接近)?一些多样性关键性状由哪些基因决定和调控,表型如何受环境影响?是否能利用其作为亲本培育出抗寒能力强的商业化品种?秋海棠的药用和食用成分(如有机酸、色素)的产品开发等等。
致 谢
感谢中科院北京植物所、昆明植物所、华南植物园、西双版纳植物园、武汉植物园、广西植物园、庐山植物园、西北农林大学、四川大学、武汉大学、广西中医药研究院、贵州科学院生物研究所、香港嘉道理农场暨植物园、台湾中央研究院以及美国密苏里植物园、德国自然博物馆等国内外近百个标本馆提供标本查阅或在线标本考证机会;感谢中科院上海辰山植物科研中心其他课题组帮忙收集部分馆藏标本和野外材料;感谢中国数字植物标本馆(CVH)、台湾生物多样性咨询入口网等提供在线标本信息,中国自然标本馆(CFH)、百度、谷歌等网站提供该种野生照片的摄影者;感谢所考察中国各级保护区的工作人员协助野外调查。
项目基金
上海市绿化和市容管理局攻关项目(F122416)。
参考文献 (References)
- Nakata, M., Ueno, T., Li, J.X., et al. (2012) Chromosome number and pollen fertility of Begonia grandis (Begoniaceae) from Japan and China. Bulletin of the Botanic Gardens of Toyama, 17, 23-29.
- Catterall, E. (1991) Begonias: The Complete Guide. Crowood Press, Marlborough.
- Streich, A. and Steinegger, D. (1984) G84-698 selecting a ground cover (revised July 1999). Historical Materials from University of Nebraska-Lincoln Extension, Lincoln, 1052.
- 李丹平 (2011) 红黑二丸中甾醇类物质提取, 鉴定及生理活性的研究. 博士论文, 华中农业大学, 武汉.
- Gao, L., Huang, X.J. and Ling, J.Y. (2012) Study on ultrasonic extraction technology of sterol from the rhizome of Begonia grandis Dry. subsp. sinensis (A. DC.) Irmsch. Medicinal Plant, 3, 39-43.
- 赵学敏(清) (1983) 本草纲目拾遗. 人民卫生出版社(第二版), 北京, 261-262.
- Guan, K.Y., Yamaguchi, H., Li, J.X., et al. (2007) Traditional uses of Begonias (Begoniaceae) in China. Acta Botanica Yunnanica, 29, 58-66.
- Sands, M.J.S. (2001) Begoniaceae in the Flora Malesiana Region. In: Saw, L.G., Chua, L.S.L. and Khoo, K.C., Eds., Taxonomy: The Cornerstone of Biodiversity, Proceedings of the 4th International Flora Malesiana Symposium, Forest Research Institute Malaysia, Kuala Lumpur, 161-168.
- Hoover, W.S., Karegeannes, C., Wiriadinata, H., et al. (2004) Notes on the geography of South-East Asian Begonia and species diversity in montane forests. Telopea, 10, 749-764.
- Aitawade, M.M. and Yadav, S.R. (2012) Taxonomic status of Begonia aliciae (Begoniaceae). Rheedea, 22, 111-115.
- Tian, D.K., Li, C., Yan, Y.H., et al. (2014) Begonia intermedia, a new species of Begoniaceae from Hainan China. Phytotaxa, 166, 114-122.
- Doorenbos, J., Sosef, M.S.M. and de Wilde, J. (1998) The Sections of Begonia. Landbouw universiteit Wageningen (Wageningen Agricultural University), Wageningen.
- Linnaeus, C. (1753) Begonia. Species plantarum. Vol. 2, Stockholm.
- 周汝昌 (2005) 周汝昌梦解红楼. 漓江出版社, 广西.
- 姜楠南, 汤庚国, 沈永宝 (2008)《红楼梦》海棠花文化考. 南京林业大学学报: 人文社会科学版, 1, 79-84.
- Dryander, J.C. (1791) On the genus of Begonia. Transactions of the Linnean Society of London, Botany, 1, 163-164.
- Otto, C.F. (1836) Allgemeine gartenzeitung, Berlin, 4, 349.
- De Candolle, A.L.P.P. (1681) Begonia grandis. In: Flora Brasiliensis, 4, 369.
- 谷粹芝, 李振宇, 黄蜀琼, 等 (1999) 中国植物志. 科学出版社, 北京, 126-269.
- Gu, C.Z., Peng, C.I. and Turland, N.J. (2007) Begoniaceae. In: Wu, Z.Y., Raven, P.H. and Hong, D.Y., Eds., Flora of China, Vol. 13, Science Press, Beijing; Missouri Botanical Garden Press, St Louis, 153-207.
- 中国科学院昆明植物研究所 (2006) 云南植物志 (第12卷). 科学出版社, 北京, 143-247.
- Shui, Y.M., Peng, C.I. and Wu, C.Y. (2002) Synopsis of the Chinese species of Begonia (Begoniaceae), with a reappraisal of sectional delimitation. Botanical Bulletin of Academia Sinica, 43, 313-327.
- 王印政 (1999) 中国植物采集简史I——1949年之前外国人在华采集. 谷粹芝, 李振宇, 黄蜀琼, 等, 编. 中国植物志(第1卷). 科学出版社, 北京, 658-704.
- 赵宝林, 陈春玲, 刘东焕等 (2013) 中华秋海棠的引种栽培和保护生物学研究. 安徽农业科学, 5, 2023-2028.
- 马宏 (2008) 中国秋海棠属秋海棠组系统学与保护生物学研究. 博士论文, 中国科学院昆明植物研究所, 昆明.
- 管开云, 李景秀, 李宏哲 (2005) 云南秋海棠属植物资源调查研究. 园艺学报, 1, 74-80.
- Kiew, R. (2005) Begonias of peninsular Malaysia. Natural History Publications, Borneo, 1-308.
- Dewitte, A., Twyford, A.D., Thomas, D.C., et al. (2011) The origin of diversity in Begonia: Genome dynamism, population processes and phylogenetic patterns. The Dynamical Processes of Biodiversity-Case Studies of Evolution and Spatial Distribution, 27-52.
- 国家中医药管理局《中华本草》编委会 (1999) 中华本草 (第14卷). 上海科学技术出版社, 上海, 490-502.
- Guan, K.Y., Yamaguchi, H., Li, J.X., et al. (2007) Traditional uses of Begonias (Begoniaceae) in China. Acta Botanica Yunnanica, 29, 58-66.
- 杨亮, 何顺志 (2008) 谈中国秋海棠属的研究进展. 时珍国医国药, 12, 2979-2981.
- 段志芳, 章炜中, 黄丽华 (2007) 紫背天葵多糖提取与含量测定. 中成药, 2, 274-276.
- 陕西省革命委员会卫生局商业局 (1971) 陕西中草药. 科学出版社, 北京, 1009-1016.
- 杨济秋, 杨济中 (1978) 贵州民间方药集. 贵州人民出版社, 贵州, 113, 183.
- 赵运林, 喻勋林, 傅晓华等 (2009) 湖南药用植物资源. 湖南科学技术出版社, 湖南, 395-396.
- 李世全 (1987) 秦岭巴山天然药物志. 陕西科学技术出版社, 陕西.
- 贵州省中医研究所 (1970)贵州草药. 贵州人民出版社, 贵州, 714.
- 江西药科学校革命委员会 (1970) 草药手册. 人民卫生出版社, 北京, 960-961.
- Forrest, L.L. and Hollingsworth, P.M. (2003) A recircumscription of Begonia based on nuclear ribosomal sequences. Plant Systematics and Evolution, 241, 193-211.
- Plana, V., Gascoigne, A., Forrest, L., et al. (2004) Pleistocene and pre-pleistocene Begonia speciation in Africa. Molecular Phylogenetics and Evolution, 31, 449-461.
- Dewitte, A., Eeckhaut, T., Van Huylenbroeck, J., et al. (2006), Flow cytometric detection of unreduced pollen in Begonia. In: XXII International Eucarpia Symposium, Section Ornamentals, Breeding for Beauty, 714, 107-112.
- Goodall-Copestake, W.P., Perez-Espona, S., David, J.H., et al. (2010) The early evolution of the mega-diverse genus Begonia (Begoniaceae) inferred from organelle DNA phylogenies. Biological Journal of the Linnean Society, 101, 243-250.
- Thomas, D. (2010) Phylogenetics and historical biogeography of Southeast Asian Begonia L. (Begoniaceae). University of Glasgow, Glasgow.
- Matolweni, L.O., Balkwill, K. and McLellan, T. (2000) Genetic diversity and gene flow in the morphologically variable, rare endemics Begonia dregei and Begonia homonyma (Begoniaceae). American Journal of Botany, 87, 431-439.
- Matsuura, H. and Okuno, S. (1936) Cytogenetical studies on Begonia. The Japanese Journal of Genetics, 12, 42-43.
- Okuno, S. and Nagai, S. (1953) Cytological studies on Begonia Evansiana Andr. with special reference to its meiotic chromosomes. The Japanese Journal of Genetics, 28, 132-136.
- 高丽 (2010) 中华秋海棠甾醇类化合物的提取纯化及微胶囊化研究. 硕士论文, 华中农业大学, 武汉.
- 李丹平, 朱乐, 余卓尔等 (2012) 红黑二丸中甾醇类物质的抑菌活性. 食品科学, 11, 70-74.
- 高丽, 黄行健, 凌洁玉 (2012) 中华秋海棠根茎中甾醇的超声提取工艺研究. 安徽农业科学, 6, 3274-3277.
- 叶丛进, 孙颖桢, 陈科力 (2006) 中华秋海棠的显微鉴别. 中药材, 5, 435-436.
- 税玉民, 李启任, 黄素华 (1999) 云南秋海棠属叶表皮及毛被的扫描电镜观察. 云南植物研究, 3, 309-316.
- Matsuura, H. and Okuno, S. (1943) Cytogenetical studies on Begonia (Preliminary survey). Cytologia, 13, 1-18.
- White, O.E., Taylor, J.H. and Speese, B.M. (1946) Begonia species hybrids. Journal of Heredity, 37, 66-70.
- Zeilinga, A.E. (1962) Cytological investigation of hybrid varieties of Begonia semperflorens. Euphytica, 11, 126-136.
- Legro, R.A.H. and Doorenbos, J. (1969) Chromosome numbers in Begonia. Netherlands Journal of Agricultural Science, 17, 189-202.
- Legro, R.A.H. and Haegeman, J.F.V. (1971) Chromosome numbers of hybrid tuberous begonias. Euphytica, 20, 1-13.
- 田代科, 管开云, 周其兴等 (2002) 云南八种秋海棠属植物的染色体数目. 云南植物研究, 2, 245-249.
- Oginuma, K. and Peng, C.I. (2002) Karyomorphology of Taiwanese Begonia (Begoniaceae): Taxonomic implications. Journal of Plant Research, 115, 225-235.
- Nakata, M., Guan, K.Y., Godo, T., et al. (2003) Cytological studies on Chinese Begonia (Begoniaceae) I. Chromosome numbers of 17 taxa of Begonia collected in 2001 field studies in Yunnan. Bulletin of the Botanic Gardens of Toyama, 8, 1-16.
- 李宏哲, 管开云, 马宏 (2005) 五种中国秋海棠植物的染色体数目. 云南植物研究, 1, 92-94.
- Nakata, M., Guan, K.Y., Li, J.X., et al. (2007) Cytotaxonomy of Begonia rubropunctata and B. purpureofolia (Begoniaceae). Botanical Journal of the Linnean Society, 155, 513-517.
- Bowden, W.M. (1945) A list of chromosome numbers in higher plants. I. Acanthaceae to Myrtaceae. American Journal of Botany, 32, 81-92.
- Dewitte, A., Leus, L., Eeckhaut, T., et al. (2009) Genome size variation in Begonia. Genome, 52, 829-838.
- 蔡红, 王明奎 (1998) 蕺叶秋海棠化学成分的研究. 天然产物研究与开发, 1, 48-50.
- 李玉环, 王锋, 刘群山 (2002) 皂化法提取番茄红素的研究. 食品科学, 8, 146-150.
- Fizet, C. (1996) Process for obtaining tocopherols and sterols from natural sources. U. S. Patent No. 5487817.
- Morin, D. (1999) Usefulness of methylated-β-cyclodextrin-based buffers for the separation of highly hydrophobic solutes in non-aqueous capillary electrophoresis. Journal of Chromatogaphyr A, 837, 281-287.
- 曹莹, 谷克仁, 孟冬等 (2006) 植物甾醇提取方法研究进展. 粮油食品科技, 5, 25-28.
- 管开云, Fershalova, T.D., Tsybulya, N.V., 等 (2005) 云南秋海棠挥发物抗微生物活性的研究. 云南植物研究, 4, 437-442.
- 管开云 (2007) 中国秋海棠属植物的多样性及保护研究. 蓝天园林, 5, 11-13.
- Nagao, M., Esashi, Y., Tanaka, T., et al. (1959) Effects of photoperiod and gibberellin on the germination of seeds of Begonia evansiana Andr. Plant and Cell Physiology, 1, 39-47.
- Esashi, Y. (1962) Studies on the formation and sprouting of aerial tubers in Begonia evansiana VII. Photo-sprouting of tuberizing buds. Plant and Cell Physiology, 3, 67-82.
- Esashi, Y. (1964) Studies on the formation and sprouting of aerial tubers in Begonia evansiana Andr. X. Tuberization under long-days and in darkness. Plant and Cell Physiology, 5, 101-117.
- Esashi, Y., Ogata, K. and Nagao, M. (1964) Studies on the formation and sprouting of aerial tubers in Begonia evansiana Andr. IX. Effects of temperature on tuber initiation. Plant and Cell Physiology, 5, 1-10.
- Esashi, Y. (1969) The relation between light and temperature effects in the induction and release of dormancy in the aerial tuber of Begonia evansiana. Plant and Cell Physiology, 10, 583-595.
- Esashi, Y., Nagao, M. (1973) Effects of oxygen and respiratory inhibitors on induction and release of dormancy in aerial tubers of Begonia evansiana. Plant and Cell Physiology, 51, 504-507.
- Esashi, Y. (1969) Different actions of red and blue or far-red lights in the photoperiodic tuberization of Begonia evansiana. Plant and Cell Physiology, 10, 349-356.
- Esashi, Y. (1963) Studies on the formation and sprouting of aerial tubers in Begonia evansiana Andr. VIII. Dual effect on tuberization of near infrared light given in dark periods. Plant and Cell Physiology, 4, 135-143.
- Esashi, Y. (1961) Studies on the formation and sprouting of aerial tubers in Begonia evansiana Andr. V. Antagonistic action of long-days to short-day response. Plant and Cell Physiology, 2, 117-127.
- Esashi, Y. (1966) The relationship between red and blue or far-red lights in the night-interruption of the photoperiodic tuberization inBegonia evansiana. Plant and Cell Physiology, 7, 405-414.
- Hashimoto, T. and Tamura, S. (1969) Effects of abscisic acid on the sprouting of aerial tubers of Begonia evansiana and Dioscorea batatas. The Botanical Magazine Tokyo, 82, 69-75.
- Okagami, N. and Nagao, M. (1973) Gibberellin-induced dormancy in Begonia aerial tubers. Increase of the growth inhibitor content by gibberellin treatment. Plant and Cell Physiology, 14, 1063-1072.
- Okagami, N. and Esashi, Y. (1972) Dormancy regulation by morphactin in aerial tubers of Begonia evansiana. Planta, 104, 195-200.
- Esashi, Y., Eguchi, T. and Nagao, M. (1964) The role of auxin in the photoperiodic tuberization in Begonia evansiana Andr. Plant and Cell Physiology, 5, 413-427.
- Sano, H. and Nagao, M. (1970) Change in the indole-3-acetic acid oxidase level in leaves of Begonia evansiana cuttings at the time of aerial tuber formation under short-day conditions. Plant and Cell Physiology, 11, 849-856.
- Okagami, N., Esashi, Y. and Nagao, M. (1977) Gibberellin-induced inhibition and promotion of sprouting in aerial tubers of Begonia evansiana Andr. in relation to photoperiodic treatment and tuber stage. Planta, 136, 1-6.
- Okagami, N. and Esashi, Y. (1972) Dormancy regulation by morphactin in aerial tubers of Begonia evansiana. Planta, 104, 195-200.
- Ahn, Y.J. and Park, J.M. (2007) Pre-adaptation to Cu during plant tissue culture enhances Cu tolerance and accumulation in Begonia (Begonia evansiana Andr.). Journal of Ecology and Field Biology, 30, 271-276.
- Thompson, M.L. (1981) Begonias: The complete reference guide. Times Books Press, New York, 1-356.
- De Wilde, J.J.F.E. (2011) Begoniaceae. Springer, Berlin, Heidelberg.
- American Begonia Society. http://wwwbegoniasorg/
- The Royal Horticultural Society Horticultural Database. http://www.rhs.org.uk/
- Speese, B.M. (1946) Begonia species hybrids. Journal of Heredity, 37, 66-70.
- 田代科, 管开云, 李景秀等 (2002) 秋海棠新品种‘大白’、‘健绿’、‘美女’和‘中大’. 园艺学报, 1, 90-91.
- 田代科, 管开云, 李景秀等 (2001) 秋海棠新品种‘大白’、‘健绿’、‘美女’和‘中大’. 园艺学报, 3, 281-282.
- 田代科, 李景秀, 管开云 (2001) 秋海棠新品种‘昆明鸟’、‘康儿’和‘白雪’. 园艺学报, 2, 186-187.
- 管开云, 李景秀, 李宏哲等 (2006) 秋海棠新品种‘紫叶’、‘紫柄’和‘大裂’. 园艺学报, 4, 933.
- 柏斌 (2011) 中科院昆明植物所7个秋海棠新品种通过专家鉴定. 中国花卉园艺, 18, 14.
- 李景秀, 管开云, 李志坚等 (2001) 秋海棠抗性育种初探. 云南植物研究, 4, 509-514.
- Missouri Botanical Garden. http://www.missouribotanicalgarden.org/
- National Capital Area Chapter. The Gesneriad Society. http://www.nationalcapitalgesneriads.org/
- Heronswood Nursery. http://heronswood.com/

NOTES
*通讯作者。