Botanical Research
Vol.07 No.02(2018), Article ID:24104,8
pages
10.12677/BR.2018.72020
Characteristics of Seed Germination and Seedling Growth of Myricaria laxiflora under Different Groundwater Table Treatments
Yanran Ma1,2, Shoupeng Guan1,2, Fangqing Chen1,2*, Kun Lv1,2
1Hubei International Scientific and Technological Cooperation Center of Ecological Protection and Management in the Three Gorges Area, China Three Gorges University, Yichang Hubei
2Engineering Research Center of Eco-Environment in the Three Gorges Reservoir Region, Ministry of Education, China Three Gorges University, Yichang Hubei

Received: Feb. 22nd, 2018; accepted: Mar. 8th, 2018; published: Mar. 21st, 2018

ABSTRACT
Objective: Soil water table and soil water content are the key factors influencing regeneration of riparian plant populations. Myricaria laxiflora has lost all the habitat in the reservoir and become an endangered species due to the construction of the Three Gorges Project. Method: This article simulates different soil water table and test the responses of M. laxiflora’ seed germination and seedling growth to soil water table to reveal the regeneration mechanism of survival populations. Result: The results show that soil water table has a significant effect on seed germination. Seed germination rate increases with the decrease of soil water table. The highest seed germination rate occurred at −10 cm soil water level treatment increasing by 200.2% contrast to the 0 cm soil water level treatment. Soil water table also influenced significantly the survival and growth of seedlings. The emergence rate, plant fresh weight and seedling length all increased significantly with the decrease of groundwater level. The emergence rate, plant fresh weight and seedling length of −10 cm soil water level treatment were higher than 0 cm soil water level by 108.5%, 158.5% and 140.7%, respectively. Conclusion: Soil water table has a significant effect on seed germination and seedling growth of M. laxiflora, which further affects population regeneration.
Keywords:Soil Water Level, Myricaria laxiflora, Seed Germination, Seedling Growth, Endangered Species
不同地下水位处理下的疏花水柏枝种子萌发 和幼苗生长特征
马嫣然1,2,官守鹏1,2,陈芳清1,2*,吕坤1,2
1湖北省三峡地区生态保护与治理国际联合研究中心,三峡大学,湖北 宜昌
2三峡库区生态环境教育部工程研究中心,三峡大学,湖北 宜昌

收稿日期:2018年2月22日;录用日期:2018年3月8日;发布日期:2018年3月21日

摘 要
目的:土壤地下水位与土壤含水量是影响河岸带植物种群更新的关键因素。疏花水柏枝因三峡工程修建失去了库区所有的生境而成为了一种濒危物种。方法:本文通过模拟不同土壤地下水位,测定了疏花水柏枝种子萌发和幼苗生长对土壤地下水位的响应,以揭示疏花水柏枝残留种群的更新机制。结果:结果表明,土壤地下水位对种子的萌发有着显著影响。种子萌发率随着土壤地下水位的下降而不断增加,−10 cm土壤水位的种子萌发率比0 cm土壤水位高201.8%。土壤地下水位还对幼苗的存活与生长有着显著影响。出苗率、植株鲜重和幼苗长度都随着地下水位的降低而显著增加,−10 cm土壤水位的出苗率、植株鲜重和幼苗长度分别比0 cm土壤水位高108.5%,158.5%和140.7%。结论:综上所述,土壤地下水位对疏花水柏枝的种子萌发和幼苗生长都有着显著影响,进而对种群的更新产生影响。
关键词 :土壤地下水位,疏花水柏枝,种子萌发,幼苗生长,濒危物种

Copyright © 2018 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
种群自然更新是指植物从其种子成熟后进入土壤及随后的种子萌发、幼苗生长,最后长成个体的连续过程 [1] 。一个种群的自然更新需要有活力充足的种子、适宜于种子萌发和幼苗生长的生境 [2] 。很多物种濒危的机制多是种群更新受阻,而造成这种结果的原因主要是物种繁殖器官的形成、种子传播、种子萌发和幼苗生长过程遇到了障碍 [3] 。种子萌发和幼苗生长是种群自然更新的关键之一阶段 [4] ,该阶段植物对外界环境胁迫的抵抗力最弱,任何不利因素都会影响到植物种子的萌发和幼苗的生长,使得种群新个体的产生和补充受阻,进而影响到种群的更新 [5] 。对于在湿地生活的植物来说,土壤地下水位的变化在其生境中是频繁出现的,地下水位的变化直接影响了植物的生长发育及分布 [6] ,最终会显著影响种群的自然更新。
疏花水柏枝(Myricaria laxiflora)是柽柳科水柏枝属的一种多年生灌木,主要分布于长江中上游干流的河滩地 [7] 。三峡工程竣工后,三峡库区的水位被抬升到145 m以上,并在145~175 m的范围内形成新的消落带。疏花水柏枝因此失去了库区所有的生境,成为了濒危物种 [8] 。濒危物种种群年龄结构往往呈现倒金字塔型的结构特征,种群中老龄个体数量多、幼龄个体数量少,种群自我更新能力弱 [9] [10] 。鲍大川等对疏花水柏枝残存种群的调查显示,该物种种群结构也呈倒金字塔型,具有幼苗数量少、种群更新困难的特征 [11] 。该问题的出现表明残存种群的种子萌发和幼苗生长两个关键阶段有可能受限。我们前期的研究显示土壤含水量对疏花水柏枝种子萌发有着显著影响,疏花水柏枝种子萌发的最低含水量为10%,在土壤含水量饱和的情况下种子萌发率最高 [12] 。由于影响河岸带土壤含水量的直接因素是水位的消涨,而关于疏花水柏枝种子萌发和幼苗生长对于地下水位之间的响应目前未见报道。三峡大坝和葛洲坝修建显著改变了下游疏花水柏枝残存种群生境的地下水位消涨节律,夏季洪水期变短,秋季水位消退进程加快,且枯水期土壤地下水位因河道下切而大幅下降 [13] 。据此我们提出了一个科学假设:疏花水柏枝残存种群受阻的原因是因为生境地下水位的显著变化而导致的。基于这个假设,我们在实验室模拟了不同土壤地下水位,通过测定各处理水平下种子萌发与幼苗生长的变化,揭示土壤地下水位对种子萌发与幼苗生长的影响。并结合疏花水柏枝残存种群生境土壤地下水位的变化规律,探讨残存种群更新受阻的机制,为疏花水柏枝的保护提供科学建议。
2. 材料与方法
2.1. 实验材料与实验设计
2017年10月29日从长江干流宜昌段江心岛胭脂坝(111˚19'34''E, 30˚38'52''N)疏花水柏枝残存种群的植株上采集果序,将所采集的果序放入自封袋中带回实验室,然后从中选择饱满的种子进行种子萌发与幼苗生长实验。
从长江干流宜昌段河滩上采集河沙,将河沙风干后过40目土壤筛,烘干混匀后分别装入长15 cm、宽15 cm和高17 cm的培养钵中,每个培养钵中装满经过处理的细沙。从上述采集回来的种子中随机选取50粒种子均匀放入一个培养皿作为一个实验单位。整个实验包含6个处理水平,分别是地下水位0 cm(表面略微积水)和经历饱和水后依此降至地下水位−1 cm (水位低于地表1 cm)、−3 cm (水位低于地表3 cm)、−5 cm (水位低于地表5 cm)、−7 cm (水位低于地表7 cm)、−10 cm (水位低于地表10 cm)。将各实验单位分别放入设置了不同水深的水箱中(长、宽、高分别为100 cm、80 cm、70 cm),培养钵下部垫有木板,通过调节木板的厚度使得培养钵基质的地下水位符合实验要求,并在培养钵低于地表处理水位处开若干小孔,保证培养钵中多余的水分迅速的流出,水箱每天换水一次。每个处理水平10个重复。
2.2. 萌发指标的测定与计算
每天统计发芽的种子数,萌发标准为明显胚根“露白” [14] ,当种子连续5 d不萌发时则认为萌发停止,计算疏花水柏枝种子在不同地下水位处理的发芽率、发芽指数、发芽势。播种后11 d统计存活的幼苗,计算疏花水柏枝在不同地下水位处理的11 d幼苗出苗率 [15] ,后面出苗率均为11 d出苗率,之后每钵留下生长状况良好的幼苗一株,待45 d后每个地下水位处理随机取出完整的幼苗5株,在室内用清水洗净后测量幼苗生长指标参数(幼苗鲜重、幼苗总长度)。同时取不同地下水位处理表层1 cm的细沙用烘干法测量含水量,每个处理3个重复。幼苗鲜重测量方法:用1/10000的电子秤测量,测量后取其平均值。幼苗总长度和幼苗表面积测量方法:用WinRHIZO根系分析系统(加拿大)测量,测量后取其平均值。
发芽率G = 发芽种子数/供试种子数 × 100%
发芽指数GI = ∑Gt/Dt = 当天的发芽数/发芽日数 + 当天的发芽数/发芽日数 + ……,Gt为t日下的发芽数目,Dt为相应的发芽天数
发芽势 = 发芽高峰期的发芽的种子数/试供种子数 × 100%
出苗率 = 存活幼苗数/试供种子数 × 100%
2.3. 数据分析
将发芽率、发芽势、发芽指数、幼苗出苗率以及幼苗鲜重、幼苗总长度等指标为因变量,将地下水位为自变量,进行单因素方差分析(One-way ANOVO),检验各个指标在处理间的差异显著性。当因素处理效应达到显著水平时,再进行多重比较,揭示各处理之间的差异水平。所有数据均采用SPSS(19.0)进行处理分析。
3. 结果与分析
3.1. 土壤地下水位对疏花水柏枝种子萌发的影响
随着地下水位的降低,不同处理的表层含水量发生了变化,−10 cm、−7 cm、−5 cm、−3 cm处理的含水量显著低于−1 cm和0 cm,但−10 cm、−7 cm、−5 cm、−3 cm之间没有显著性差异,0 cm、−1 cm之间也没有显著性差异。
土壤地下水位对疏花水柏枝的萌发有显著的影响(P < 0.05)。发芽率、发芽指数和发芽势随着地下水位的下将而呈现显著上升的趋势。其中−10 cm处理的发芽率、发芽指数和发芽势显著高于−7 cm、−5 cm、−3 cm、−1 cm和0 cm等各处理水平,为所有处理中发芽率最高,而0 cm处理水平的各项指数均为最低。0 cm、发芽率在各处理之间都存在显著差异,而发芽指数和发芽势在0 cm、−1 cm、−3 cm处理间没有显著性差异,而在−10 cm、−7 cm、−5 cm处理间有显著性差异(表1)。−10 cm处理的发芽率、发芽指数和发芽势分别比0 cm处理水平高201.8%、198.7%和212.5%。
3.2. 土壤地下水位对幼苗存活和生长的影响
土壤地下水位对幼苗出苗率有着显著影响(P < 0.05)。随着地下水位的降低,疏花水柏枝幼苗出苗率呈先增加后降低的趋势。地下水位0 cm、−1 cm、−3 cm、和−5 cm时,幼苗出苗率分别为11.8%、16.6%、18.2%和16.0%,无显著性变化;当地下水位为−7 cm时,幼苗出苗率显著上升,达到所有处理中最高值34.6%。其后随着土壤地下水位降低,幼苗出苗率有所减少。−7 cm处理的最高出苗率与0 cm处理的最低出苗率相比增加了193.2% (图1)。
幼苗的生长也受到了地下水位的显著影响,幼苗的鲜重和长度随着地下水位的降低而呈增加的趋势(P < 0.05)。地下水位为0 cm和−1 cm时,幼苗鲜重和幼苗长度分别为0.53 mg和0.67 mg与2.7 mm和3.2 mm,两个处理间没有显著差异;地下水位降低到−3 cm、−5 cm、−7 cm和−10 cm时,幼苗鲜重和幼苗长度比0 cm和−1 cm处理显著增加。−10 cm处理的幼苗鲜重和幼苗长度与0 cm处理相比分别增加了158.5%和140.7%。
3.3. 种子萌发率及幼苗存活与地下水位的相关性分析
疏花水柏枝新鲜成熟种子的萌发率与幼苗出苗率与地下水位存在显著的负相关关系(表2),也即随

Table 1. Characteristics of seed germination under different groundwater level treatments
表1. 不同土壤地下水位的种子萌发特征
注:同列不同字母表示差异显著(P < 0.05)。
注:不同字母表示不同处理间差异显著水平(P < 0.05)
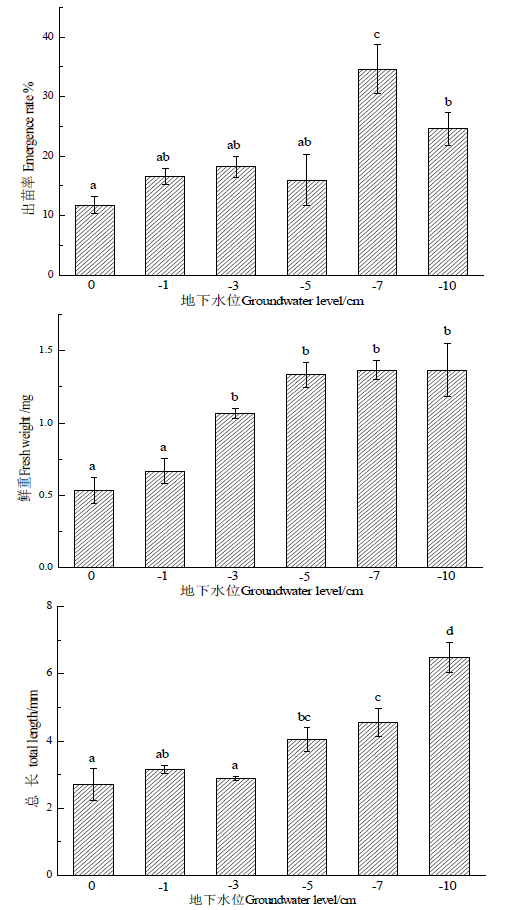
Figure 1. Effect of different soil groundwater levels on seedling survival and growth
图1. 不同土壤地下水位对幼苗存活和生长的影响
Table 2. Correlation coefficient between seed germination, seedling emergence and soil groundwater level
表2. 幼苗出苗率、种子萌发参数与地下水位之间的相关系数
注:**表示在0.01水平(双侧)上显著相关。
着地下水位的下降,种子萌发率和幼苗出苗率都会显著的上升,其中种子萌发率与地下水位的相关系数的绝对值大于幼苗出苗率与地下水位的相关系数的绝对值。种子萌发率与幼苗出苗率存在着显著正相关关系。
4. 讨论
土壤地下水位的变化是影响湿地植物种群演替的关键因素之一 [16] ,合适的土壤地下水位有利于种群的更新 [17] ,土壤地下水位的变化不仅会影响到种子萌发与幼苗生长的水位需求,同时还会影响到土壤中氧气状况 [18] ,并由此影响到种子萌发与幼苗生长进程。Keddy等研究了种子大小、土壤粒径和地下水位对植物种子萌发的影响,最终结果显示对种子萌发起决定作用的是土壤地下水位 [19] 。种子萌发阶段,水分进入种子后,胚逐渐增大活化,进而诱导赤霉素和各种酶的产生,促进种子的萌发 [20] 。水分过多或过少都不利于种子的萌发,水分过多会导致种子萌发时缺氧,不仅会降低种子的萌发率,还会造成幼苗形态发生异常,水分过少,又会导致干旱,使种子无法吸收足够的水分进行萌发 [21] 。不同种类的植物种子的萌发和幼苗生长对地下水位的响应也不同。徐玲玲等研究了枫杨种子和幼苗对地下水位及埋藏深度的响应,研究结果表明枫杨种子萌发的最适水位为0~−4 cm;幼苗定植与生长的最适水位为-6 cm。随埋藏深度增加,枫杨出苗率下降 [22] 。黄振英等研究了白沙蒿种子萌发与幼苗生长与土壤地下水位的关系,发现受土壤地下水位所影响的土表水分在1.7%~14.7%的范围内,种子萌发率随水分升高而增加,但是当土表水分超过19.4%,种子萌发与幼苗生长均受到抑制 [23] 。本研究结果显示疏花水柏枝种子萌发的最适地下水位为−10 cm,幼苗存活的最适地下水位为−7 cm,幼苗生长的最适地下水位为−10 cm。说明适当低的土壤地下水位有利于疏花水柏枝的种子萌发与幼苗生长。高的土壤地下水位对疏花水柏枝种子萌发以及幼苗存活与生长有抑制作用,可能是高水位使土壤氧气缺少,处于还原状态,影响到种子和幼苗根系的呼吸,并进一步影响种子萌发及幼苗的存活与生长 [24] 。
三峡大坝下游残存的疏花水柏枝种群目前报道的有三处,全部位于长江干流宜昌段的江心岛 [25] 。鲍大川等2008与2009年调查宜都关洲疏花水柏枝残存种群结构动态的结果显示该残存种群幼苗数量少,种群更新存在障碍 [11] 。疏花水柏枝一般在秋季水位消退后开始开花结果,种子成熟后直接经风传播直接落入地面。如果地下水位引起的地表水分适宜,种子一般就能渗入沙土中就地萌发 [25] 。但是三峡大坝和葛洲坝的修建显著改变了下游三个疏花水柏枝残存种群生境的地下水位消涨节律,使得夏季洪水期变短,秋季水位消退进程加快,且枯水期土壤地下水位因河道下切而大幅下降 [13] 。疏花水柏枝种子萌发和幼苗生长既需要充足的水分又需要比较充沛的氧气,因此适宜的地下水位有利于种子的萌发与幼苗生长。本研究结果表明−10 cm左右的地下水位比较适于疏花水柏枝种子萌发和幼苗生长。大坝修建之前,长江干流河岸带的水位是自然消退,其进程一般比较缓慢,经风直接传播的种子落入地表后或直接萌发或通过水流二次传播后另找安全地点进行萌发与生长,实现自然更新 [25] 。大坝修建之后,由于秋季长江流域的雨量偏少,同时水库又要蓄水发电,下游三个疏花水柏枝残存种群生境地的土壤水位下降速度较快 [13] ,减少了种子经水流传播的途径,同时也难以满足于疏花水柏枝种子萌发和幼苗生长对地下水位的需求,种群的更新受到严重影响。因此要保证种群的自然更新,就必须在疏花水柏枝种子萌发和幼苗生长阶段,降低库区的蓄水速率,减少土壤地下水位的下降幅度与速率,以确保疏花水柏枝残存种群更新的生态需水。
基金项目
国家自然科学基金项目(31770564)和三峡库区生态环境教育部工程研究中心开放基金(KF2016-01)。
文章引用
马嫣然,官守鹏,陈芳清,吕 坤. 不同地下水位处理下的疏花水柏枝种子萌发和幼苗生长特征
Characteristics of Seed Germination and Seedling Growth of Myricaria laxiflora under Different Groundwater Table Treatments[J]. 植物学研究, 2018, 07(02): 150-157. https://doi.org/10.12677/BR.2018.72020
参考文献
- 1. 潘开文, 何静, 吴宁. 森林凋落物对林地微生境的影响[J]. 应用生态学报, 2004, 15(1): 153-158.
- 2. 曾德慧, 尤文忠, 范志平, 等. 樟子松人工固沙林天然更新特征[J]. 应用生态学报, 2002, 13(1): 1-5.
- 3. 陈发菊, 梁宏伟, 王旭, 等. 濒危植物巴东木莲种子休眠与萌发特性的研究[J]. 生物多样性, 2007, 15(5): 492-499.
- 4. Moles, A.T. and Westoby, M. (2004) What Do Seedlings Die from and What Are the Implications for Evolution of Seed Size? Oikos, 106, 193-199.
https://doi.org/10.1111/j.0030-1299.2004.13101.x - 5. Mills, M.H. and Schwartz, M.W. (2005) Rare Plants at the Extremes of Distribution: Broadly and Narrowly Distributed Rare Species. Biodiversity & Conservation, 14, 1401-1420.
https://doi.org/10.1007/s10531-004-9666-6 - 6. Stanturf, J.A., Conner, W.H., Gardiner, E.S., Schweitzer, C.J. and Ezell, A.W. (2004) Recognizing and Overcoming Difficult Site Conditions for Afforestation of Bottomland Hardwoods. Ecological Restoration, 22, 183-193.
https://doi.org/10.3368/er.22.3.183 - 7. 陈芳清, 谢宗强, 熊高明, 等. 三峡濒危植物疏花水柏枝的回归引种和种群重建[J]. 生态学报, 2005, 25(7): 1811-1817.
- 8. 陈芳清, 谢宗强. 濒危植物疏花水柏枝种群生态保护研究进展[C]//全国生物多样性保护与持续利用研讨会. 2004.
- 9. 刘海洋, 金晓玲, 沈守云, 等. 湖南珍稀濒危植物——珙桐种群数量动态[J]. 生态学报, 2012, 32(24): 7738-7746.
- 10. 张文辉, 许晓波, 周建云, 等. 濒危植物秦岭冷杉种群数量动态[J]. 应用生态学报, 2005, 16(10): 1799-1804.
- 11. 鲍大川, 卢志军, 江明喜, 等. 三峡大坝下游残存疏花水柏枝种群结构和动态[J]. 植物科学学报, 2010, 28(6): 711-717.
- 12. 袁万群, 詹海燕, 陈芳清, 等. 濒危植物疏花水柏枝种子萌发的生态学特性[J]. 生态环境学报, 2008, 17(6): 2341-2345.
- 13. 段唯鑫, 郭生练, 王俊. 长江上游大型水库群对宜昌站水文情势影响分析[J]. 长江流域资源与环境, 2016, 25(1): 120-130.
- 14. 秦洪文, 刘正学, 钟彦, 等. 濒危植物疏花水柏枝种子萌发力丧失及贮藏条件研究[J]. 河南农业科学, 2013, 42(2): 116-119.
- 15. 陈叶, 郝宏杰, 王治江, 等. 不同处理对甜叶菊种子出苗率和幼苗生长的影响研究[J]. 种子, 2012, 31(4): 102-104.
- 16. 宋长春. 湿地生态系统对气候变化的响应[J]. 湿地科学, 2003, 1(2): 122-127.
- 17. 李红丽, 智颖飙, 雷光春, 等. 不同水位梯度下克隆植物大米草的生长繁殖特性和生物量分配格局[J]. 生态学报, 2009, 29(7): 3525-3531.
- 18. 徐玲玲. 储藏方式、时间及环境因子对枫杨种子萌发及幼苗生长的影响[D]: [硕士学位论文]. 武汉: 华中师范大学, 2012.
- 19. Keddy, P.A. and Constabel, P. (1986) Germination of Ten Shoreline Plants in Relation to Seed Size, Soil Particle Size and Water Level: An Experimental Study. Journal of Ecology, 74, 133-141.
- 20. 林涛, 田有亮, 王燕, 等. 基质含水量、温度条件对柠条等5种沙生植物种子萌发的影响[J]. 内蒙古农业大学学报(自然科学版), 2008, 29(4).
- 21. 任坚毅, 林玥, 岳明. 太白山红桦种子的萌发特性[J]. 植物生态学报, 2008, 32(4): 883-890.
- 22. 徐玲玲, 雷耘, 汪正祥. 枫杨种子和幼苗对地下水位及埋藏深度的响应[J]. 生态与农村环境学报, 2013, 29(1): 49-52.
- 23. 黄振英, 胡正海, 张新时. 白沙蒿种子传播和萌发特性的生理生态学研究[C]//西部地区植物科学与开发学术讨论会. 2001.
- 24. 田应兵, 艾天成. 湿地土壤及其生态功能[J]. 生态学杂志, 2002, 21(6): 36-39.
- 25. 陈芳清, 王传华. 三峡珍稀濒危植物疏花水柏枝的生态保护[M]. 武汉: 科学出版社, 2015.