Botanical Research
Vol.
11
No.
03
(
2022
), Article ID:
51544
,
9
pages
10.12677/BR.2022.113034
方山柿杂交种子不同父本来源花粉的萌发及 胚胎发育的观察
王阳1,吕旭东2,李军萍3,朱德俊3,俞敏娜1,高海力4,应尚蛟3,吕洪飞1*,严玲玲2*
1浙江理工大学生命科学与医药学院,浙江 杭州
2浙江省耕地质量与肥料管理总站,浙江 杭州
3浙江省永康市自然资源和规划局,永康林业技术和种苗推广站,浙江 金华
4浙江省公益林和国有林场管理总站,浙江 杭州
收稿日期:2022年4月7日;录用日期:2022年5月9日;发布日期:2022年5月23日

摘要
方山柿为优良的孤雌生殖无种子品种,为探究其杂交种子的形成,对太秋甜柿、野柿花粉的离体萌发率、方山柿与太秋甜柿、野柿杂交授粉亲和性以及方山柿的胚胎发育过程进行了研究。结果表明,太秋甜柿及野柿花粉萌发的最佳离体培养基配方为10%蔗糖、100 mg/L氯化钙、0.2%琼脂,并用硼酸调节pH至5.5;花粉离体培养6 h后,苯胺蓝染色观察:太秋甜柿花粉的萌发率为23.5% ± 0.2%,野柿的萌发率为25.6% ± 0.4%;太秋甜柿和野柿的花粉授粉与方山柿均具亲和性;正常杂交的胚在授粉后37、45、54、82 d依次发育至球形期、心形期、鱼雷形期和子叶形期,成熟种子纵轴长度大于2 cm;早期胚败育型表现为杂交胚退化消失,晚期胚败育型为杂交胚发育滞后,胚的着生位置偏斜。可得出结论,太秋甜柿及野柿均能成为方山柿杂交种子的父本来源,其中野柿的花粉活力更强,方山柿杂交后的胚胎发育的过程遵循柿属植物胚胎的发育规律。
关键词
方山柿,太秋甜柿,野柿,花粉萌发,花粉管生长,胚胎发育,胚败育

Observation on Pollen Germination and Embryonic Development of Different Paternal Parents of Hybrid Seeds of Diospyros kaki Thunb. cv. Fangshan
Yang Wang1, Xudong Lv2, Junping Li3, Dejun Zhu3, Minna Yu1, Haili Gao4, Shangjiao Ying3, Hongfei Lv1*, Lingling Yan2*
1College of Life Sciences and Medicine, Zhejiang Sci-Tech University, Hangzhou Zhejiang
2Zhejiang General Station of Cultivated Land Quality and Fertilizer Management, Hangzhou Zhejiang
3Department of Natural Resources and Planning of Yongkang, Yongkang Forestry Technology and Seedling Promotion Station, Jinhua Zhejiang
4Zhejiang Public Welfare Forest and State-Owned Forest Farm Management Station, Hangzhou Zhejiang
Received: Apr. 7th, 2022; accepted: May 9th, 2022; published: May 23rd, 2022

ABSTRACT
Diospyros kaki Thunb. cv. Fangshan is an excellent seedless parthenogenetic variety. To explore the formation of its hybrid seeds, the germination rate of pollen of D. kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino in vitro, the cross-pollination compatibility of D. kaki Thunb. cv. Fangshan with pollen of D. kaki Thunb. cv. “Taishuu” or D. kaki Thunb. var. sylvestris Makino, and the embryonic development process of D. kaki Thunb. cv. Fangshan hybrid seeds were studied. The results showed that the best in vitro medium is 10% sucrose, 100 mg/L calcium chloride, 0.2% agar and adjust the pH of boric acid to 5.5. The pollen germination rate of D. kaki Thunb. cv. “Taishuu” was 23.5% and that of D. kaki Thunb. var. sylvestris Makino was 25.6%. The cross pollination of D. kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino with D. kaki Thunb. cv. Fangshan had affinity. Normal hybrid embryo developed to globular embryo, heart-shaped stage, torpedo shaped stage and early cotyledon stage at 37, 45, 54 and 82 days after pollination, and the longitudinal axis length of seeds exceeded 2 cm after full maturity. The early embryo abortion type showed that the hybrid embryo degenerated and disappeared. The late embryo abortion type is hybrid embryo development lag, and the position of embryo implantation was skewed. It can be concluded that D. kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino could become the male parent source of D. kaki Thunb. cv. Fangshan hybrid seeds, D. kaki Thunb. var. sylvestris Makino has stronger pollen vitality. The embryonic development process of D. kaki Thunb. cv. Fangshan hybrid seeds followed the Diospyros developmental regularity.
Keywords:Diospyros kaki Thunb. cv. Fangshan, D. kaki Thunb. cv. “Taishuu”, D. kaki Thunb. var. sylvestris Makino, Pollen Germination, Pollen Tube Growth, Embryo Development, Embryo Abortion

Copyright © 2022 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
在植物学和园艺学上,单性结实是指植物的雌花不经传粉、受精,子房直接发育形成果实,这种果实不含种子 [1]。单性结实存在两种情况,天然单性结实和刺激单性结实 [2],如香蕉、菠萝、无核蜜橘等属于天然单性结实,刺激单性结实是指在生长发育的过程中受到外界的物理和化学刺激,诱导形成的无籽果实,例如盛花期喷施赤霉素可以诱导梨的单性结实 [3]。
《中国植物志》记载柿科(Ebenaceae)有3属共500余种,我国有1属,约57种,《中国果树志–柿卷》记载的柿品种有963个,无核及少核柿品种158个。方山柿(Diospyros kaki Thunb. cv. Fangshan)是浙江永康的地理标志农产品,属于天然单性结实,在我国有着悠久栽培历史 [4]。随着方山柿生产标准的全面实施,以及病虫害防治等栽培技术的落实 [5],目前方山柿的主要研究方向集中于经济价值的提升和其市场销售优势的扩大,包括防止褐变技术、脱涩技术的开发 [6];设施栽培下方山柿的最佳采收期、采后最佳储藏方式的探究 [7];方山柿的控时催熟以及品质控制 [8]。近年来,方山柿栽培区太秋甜柿的大量引种,外源花粉污染导致大量方山柿产生杂交种子,致使方山柿失去了原有的品质和口感,降低了方山柿的口碑及市场竞争力,不仅给农户造成了巨大的经济损失,也不利于该种质资源的保护。方山柿栽培区域内不仅存在引种的太秋甜柿,还有零散分布的野柿。在方山柿的盛花期,野柿和太秋甜柿的花粉均可通过虫媒对方山柿授粉。
日本学者在单性结实品种“平核无”的生殖研究过程中发现,“平核无”的幼果时期均含有发育不完全的种子,幼胚的胚轴及子叶发育畸形或未完全分化。柿果成熟后,种子会完全退化,导致无核 [9] [10]。国内学者对单性结实品种磨盘柿进行人工杂交,结果发现,磨盘柿杂种败育的比例极高,早期败育型杂种的初生胚乳核分裂出现异常,胚只能发育至球形期早期;晚期败育型杂种的胚乳没有明显的异常现象,但胚发育滞后 [11]。本文对野柿及太秋甜柿花粉离体培养基配方进行优化,并测定太秋甜柿、野柿花粉的离体萌发率及太秋甜柿、野柿与方山柿杂交授粉的亲和性,揭示方山柿杂交后的胚胎发育过程。旨在为方山柿杂交种子的形成及父本来源的进一步研究提供参考,为方山柿资源的综合开发和保护提供理论基础和技术指导。
2. 材料与方法
2.1. 实验材料
实验材料:太秋甜柿(Diospyros kaki Thunb. cv. “Taishuu”)、野柿(D. kaki Thunb. var. sylvestris Makino.)、方山柿(D. kaki Thunb. cv. Fangshan)采自浙江省永康市凌宅村。
2.2. 花粉采集
2021年5月初,采集太秋甜柿和野柿即将散粉的雄花。于室温(23℃左右)下阴干24小时后,用毛笔收集干燥后的花粉于培养皿中,再将培养皿放入装有干燥硅胶的密封袋中,4℃贮藏备用 [12]。
2.3. 杂交授粉
2021年5月初,于永康市凌宅村方山柿基地,用毛笔采集甜柿和野柿的花粉,选取即将开放的方山柿雌花,剥离其花瓣后对其柱头进行授粉,标记后套袋,10天后去除硫酸纸袋 [13]。
2.4. 离体萌发测定花粉活力
采用不同蔗糖浓度的半固体培养基 [14] [15] (表1),取适量灭菌后的培养基滴于凹玻片的凹槽中心,避免培养基超过槽口,在培养基冷却凝固后,用毛笔轻蘸花粉,将其均匀抖落于培养基上,后放入垫有湿滤纸的培养皿中,保持环境湿度。于室温(23℃)下暗培养6 h,0.1%水溶性苯胺蓝溶液进行染色后,在Olympus IX71型荧光显微镜下观察并拍照记录 [16] [17]。

Table 1. In vitro medium formula
表1. 离体培养基配方
2.5. 花粉活体萌发检测方山柿杂交授粉亲和性
用太秋甜柿或野柿花粉人工授粉方山柿24 h后,取授粉后的方山柿雌蕊用FAA固定液固定24 h,然后转入70%乙醇中;用刀片截取花柱及柱头,置于5 mol/L NaOH中软化脱色处理5 h左右,清水漂洗;用镊子轻取花柱及柱头于载玻片上,0.1%水溶性苯胺蓝溶液进行染色后进行整体压片,在Olympus IX71型荧光显微镜下观察并拍照记录 [18] [19]。
2.6. 胚发育过程观察
用标签标记不同授粉时间的方山柿雌花。自授粉后20 d起,每隔7~10 d取样,从子房中剥取出种子用FAA固定液保存。在Olympus BX51型荧光显微镜下解剖观察并拍照记录 [20] [21]。
3. 实验结果与分析
3.1. 太秋甜柿及野柿花粉最佳离体培养基的探究
柿花粉离体培养48 h后,如图1(A)所示,在1#培养基(无蔗糖)中,花粉粒细胞内部渗透压较高,花粉粒胀大饱满,并未观察到花粉粒萌发;如图1(B)所示,在2#培养基(5%蔗糖)中,有观察到花粉管的萌发,同时菌类繁殖较为旺盛,严重影响花粉管的观察;如图1(C)所示,在3#培养基(10%蔗糖)中,有观察到花粉管萌发,菌类繁殖较少,便于花粉管观察;如图1(D)所示,在4#培养基(20%蔗糖)中,花粉粒细胞内部渗透压较低,观察到花粉粒脱水干瘪,并未观察到花粉粒萌发。

Figure 1. Microscopic observation of pollen of Diospyros kaki Thunb. cv. “Taishuu” and Diospyros kaki Thunb. var. sylvestris Makino cultured in vitro with different sucrose concentration medium for 48 h, bar = 200 μm. A: 1#Medium (0% sucrose); B: 2#Medium (5% sucrose); C: 3#Medium (10% sucrose); D: 4#Medium (20% sucrose)
图1. 太秋甜柿及野柿花粉不同蔗糖浓度培养基离体培养48 h显微观察图,Bar = 200 μm。A:1#培养基(无蔗糖);B:2#培养基(5%蔗糖);C:3#培养基(10%蔗糖);D:4#培养基(20%蔗糖)
3.2. 太秋甜柿和野柿花粉离体萌发率的测定
如图2,室温(23℃)下暗培养6 h后,太秋甜柿和野柿的部分花粉已经萌发,花粉管开始生长。荧光显微镜下花粉粒呈蓝色,花粉管呈绿色。太秋甜柿的花粉萌发率为23.5% ± 0.2%,野柿的花粉萌发率25.6 ± 0.4%。野柿的花粉萌发率极显著高于太秋甜柿(图3)。
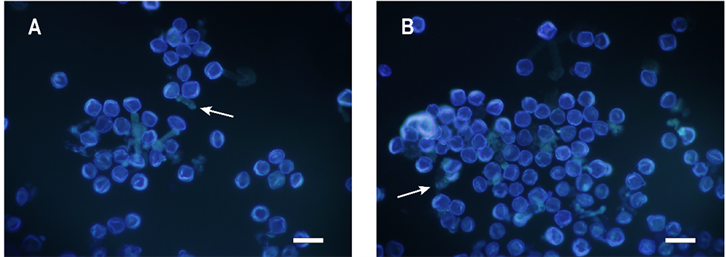
Figure 2. Observation on aniline blue staining of pollen of Diospyros kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino after cultured in the best medium for 6 hours, bar = 100 μm. A: D. kaki Thunb. cv. “Taishuu” pollen; B: D. kaki Thunb. var. sylvestris Makino pollen; the arrow shows pollen tube
图2. 太秋甜柿和野柿花粉最佳培养基离体培养6 h后苯胺蓝荧光染色观察,Bar = 100 μm。A:太秋甜柿花粉;B:野柿花粉;箭头示花粉管
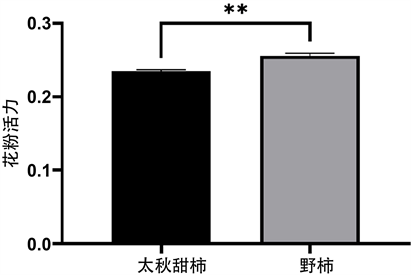
Figure 3. Comparison of pollen vitality between Diospyros kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino (p < 0.01)
图3. 太秋甜柿和野柿花粉活力(p < 0.01)
3.3. 太秋甜柿及野柿花粉在方山柿柱头萌发及其花柱中花粉管生长的观察
如图4所示,太秋甜柿和野柿花粉在方山柿柱头上都能正常萌发,且花粉管可在花柱中正常生长,荧光染色后可以清晰的观察到花柱中的花粉管。
3.4. 方山柿种子发育过程观察
杂交授粉后28 d,从方山柿果中剥离出的种子纵径长约4~5 mm,横径长约3 mm。种子解剖后,未观察到胚(图5(A))。授粉后37 d,剥离出的种子纵径达到7~8 mm,横径达到5 mm,解剖后可见球形胚,球形胚直径小于0.2 mm (图5(B-a))。授粉后45 d的杂交种子,纵径长9~10 mm,横径5~6 mm,幼胚的发育至心形期(图5(B-b)),胚体顶部两侧的细胞分裂较快形成两个突起。授粉后54 d后,杂交种子体积显著增大,约为发育45 d杂交种子体积的一倍,种子内可观察到早期的鱼雷形胚(图5(B-c)),心形胚时期形成的两个突起开始伸长。授粉后63 d的杂交种子同授粉后54 d的杂交种子在大小上无显著差异,胚发育至鱼雷形胚晚期(图5(B-d)),胚体顶部的突起发育成两片子叶。杂交授粉后82 d,种子大小又有显著增长,种子的胚乳开始硬化,种子内能观察到子叶形胚(图5(B-e))。授粉后157 d,方山柿果成熟(图5(C)),剥离出的种子被浅黄色肉质胎座包裹(图5(D)),胚发育成熟(图5(E)),胚呈乳白色,子叶二枚,子叶较为肥厚。
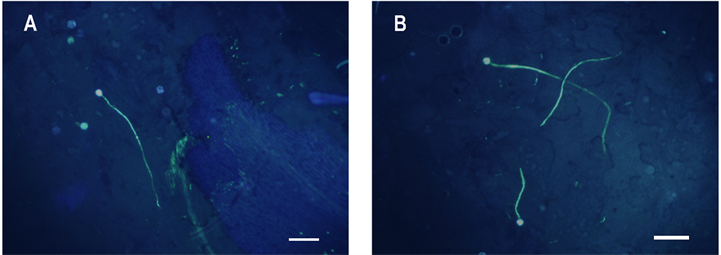
Figure 4. Fluorescence observation of pollen germination of Diospyros kaki Thunb. cv. “Taishuu” and D. kaki Thunb. var. sylvestris Makino on stigma and pollen tube grows in style of D. kaki Thunb. cv. Fangshan by aniline blue staining, bar = 200 μm. A: D. kaki Thunb. cv. “Taishuu”; B: D. kaki Thunb. var. sylvestris Makino
图4. 苯胺蓝染色荧光观察太秋甜柿和野柿花粉在方山柿柱头萌发及其花柱中花粉管生长,Bar = 200 μm。A:太秋甜柿;B:野柿


Figure 5. Hybrid seed morphology, embryonic development, and fruit of Diospyros kaki Thunb. var. sylvestris Makino pollinated D. kaki Thunb. cv. Fangshan at different developmental stages. A: Morphology of hybrid seeds at different developmental stages after artificial pollination. B: Morphological changes of embryos at different development times of hybrid seeds: (a) Globular embryo; (b) Heart-shaped embryo; (c) Early torpedo-shaped embryo; (d) Late torpedo-shaped embryo; (e) Early cotyledon embryo. C: D. kaki Thunb. cv. Fangshan. D: seeds wrapped in placenta. E: Mature embryo
图5. 野柿授粉方山柿不同发育时期的杂交种子形态及其胚胎发育过程和果实。A:人工授粉后不同发育时期杂交种子形态。B:杂交种子不同发育阶段胚的形态:(a) 球形胚;(b) 心形胚;(c) 早期鱼雷形胚;(d) 晚期鱼雷形胚;(e) 早子叶形胚。C:方山柿。D:胎座包裹的种子。E:成熟胚
3.5. 败育的杂交种子形态观察
在方山柿杂交种子的胚胎发育过程中,部分胚的子叶和胚轴呈畸形或未完全分化,出现胚败育的现象。与胚胎发育正常的种子相比较(图6(A)),早期败育型杂交种子(图6(B)、图6(C)),未见胚的结构,杂交种子中仅有半透明的胚乳,部分胚乳的中间留有空腔。晚期败育型杂交种子(图6(D)-(F)),能观察到乳白色胚,但胚的形态异常且发育滞后,着生位置偏斜,胚没有分化出胚芽、胚轴、胚根和子叶等结构。
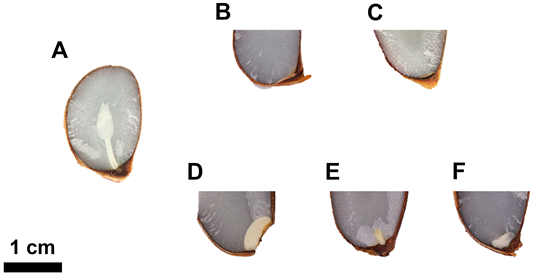
Figure 6. Morphological observation of normal embryo and abortion embryo of Diospyros kaki Thunb. cv. Fangshan hybrid seed. A: D. kaki Thunb. cv. Fangshan hybrid seeds containing mature embryos; B, C: Early abortion of D. kaki Thunb. cv. Fangshan hybrid seeds, no embryo was observed, D, E, F: Late abortion D. kaki Thunb. cv. Fangshan hybrid seeds, the position of embryo implantation is skewed, and impossible to distinguish the embryo bud, hypocotyl, radicle, cotyledon
图6. 方山柿杂交种子正常胚及败育胚和胚乳的形态。A:含正常胚的方山柿杂交种子;B,C:早期胚败育的方山柿杂交种子;D,E,F:晚期胚败育的方山柿杂交种子,示胚的着生位置偏斜
4. 讨论
在花粉的离体萌发实验中,离体培养基的组成、pH值及培养温度会影响花粉的萌发率,其中培养基的组成成分对花粉的萌发率有较大的影响。培养基中的蔗糖及其他矿物质能给花粉管的萌发提供必要的营养,但蔗糖及其他矿物质浓度过高会导致花粉粒脱水干瘪失去活性,抑制花粉萌发 [22] [23]。所以在花粉离体培养基中,合适的蔗糖浓度及其他矿物质浓度,能提高花粉离体培养的萌发率,更准确地反映其花粉活力。糖类物质是花粉萌发及花粉管生长所需的重要营养物质,周瑞金等人研究发现 [24],在不含糖的培养基中花粉的萌发率会显著低于10%蔗糖浓度的培养基。花粉离体培养基中适宜浓度的钙离子有利于花粉管的生长,硝酸钙浓度在0~250 mg/L时花粉的萌发率较高 [25],实验参考了詹术森等人研究结果,花粉离体培养基中硝酸钙浓度为100 mg/L时,柿花粉的萌发率最高 [26]。张绍铃等人发现花粉萌发的最佳pH范围在4~9,培养基过酸性或过碱性会降低花粉萌发率 [27]。实验结果表明,适合太秋甜柿及野柿花粉萌发的最佳培养基配方为10%蔗糖,0.2%琼脂,100 mg/L氯化钙,并用硼酸调节pH至5.5。太秋甜柿、野柿花粉的离体萌发观察结果显示,野柿的花粉粒活性显著高于太秋甜柿花粉粒,这与张志芳 [8] 的染色法花粉活力检测结果一致。
苯胺蓝通过结合花粉及花粉管壁的胼胝质产生荧光,顾翠花等人发现,花粉离体培养6 h后花粉管中形成大量的胼胝质 [28],离体培养时间超过12小时的花粉粒,苯胺蓝染色后荧光显微镜观察,花粉粒荧光显色过强,会干扰花粉管的观察 [29];离体培养超过48小时,播撒花粉时所携带的细菌会大幅度污染培养基,影响显微观察 [30] [31]。太秋甜柿和野柿花粉人工授粉6 h后方山柿柱头荧光染色,观察到野柿、太秋甜柿授粉方山柿均具有亲和性,说明引种的太秋甜柿和周围的野柿均有可能成为方山柿杂交种子的父本。
扈惠灵等 [32] 人发现磨盘柿在授粉后35、40、45、55 d,杂种胚依次发育至球形期、心形期、鱼雷期和子叶期;徐莉清等 [33] 人发现完全甜柿品种“次郎”在授粉后40 d幼胚发育至球形胚、50 d发育至心形胚或鱼雷形胚、60 d后发育至子叶形胚。方山柿在授粉后37 d、45 d、54 d、63 d、82 d,杂种胚依次发育至球形胚、心形胚、早期鱼雷形胚、晚期鱼雷形胚、子叶形胚,其有性生殖过程与其它柿属植物相似,且种子形成过程存在不同的败育现象。
综上所述,方山柿杂交种子的产生是因野柿、太秋甜柿花粉的授粉引起,其中野柿的花粉活力较强,其胚胎发育遵循柿属植物胚胎的发育规律,上述研究结果为进一步研究方山柿杂交种子父本来源提供参考。
5. 结论
本文通过对太秋甜柿、野柿花粉的离体萌发、方山柿雌花人工授粉及其花粉在柱头萌发和花粉管在花柱中生长的观察,以及方山柿胚胎发育过程的研究,获得以下主要结论:主要获得以下结论:1) 适合太秋甜柿及野柿花粉萌发的最佳培养基配方为10%蔗糖、100 mg/L氯化钙、0.2%琼脂,并用硼酸调节pH至5.5;2) 野柿的花粉粒活性显著高于太秋甜柿花粉粒;3) 野柿、太秋甜柿授粉方山柿均具有亲和性;4) 方山柿胚胎发育过程的与柿属的其它植物相似,种子形成过程存在不同的败育现象。
文章引用
王 阳,吕旭东,李军萍,朱德俊,俞敏娜,高海力,应尚蛟,吕洪飞,严玲玲. 方山柿杂交种子不同父本来源花粉的萌发及胚胎发育的观察
Observation on Pollen Germination and Embryonic Development of Different Paternal Parents of Hybrid Seeds of Diospyros kaki Thunb. cv. Fangshan[J]. 植物学研究, 2022, 11(03): 290-298. https://doi.org/10.12677/BR.2022.113034
参考文献
- 1. Murakami, S. (2020) Study of Parthenocarpy in Three Kiwifruit Commercial Cultivars. International Journal of Fruit Science, 20, S802-S811. https://doi.org/10.1080/15538362.2020.1769525
- 2. He, M., Song, S.W., Zhu, X.Y., et al. (2021) SITPL1 Silencing Induces Facultative Parthenocarpy in Tomato. Frontiers in Plant Science, 12, Article ID: 672232. https://doi.org/10.3389/fpls.2021.672232
- 3. 许楚楚. 利用植物生长物质诱导梨单性结实的研究[D]: [硕士学位论文]. 杭州: 浙江大学, 2017.
- 4. 贾美巧, 徐苏君. 方山柿品牌战略发展初探[J]. 柑桔与亚热带果树信息, 2005, 21(4): 15.
- 5. 董洪彪. 方山柿无公害栽培技术[J]. 浙江林业技, 2004, 24(5): 27-29.
- 6. 王书民, 张国春. 柿子的加工技术[J]. 商洛师范专科学校学报, 2004, 18(2): 43-45.
- 7. 卢婧玮. 设施栽培下方山柿最佳采收期及采后储藏的研究[D]: [硕士学位论文]. 金华: 浙江师范大学, 2016.
- 8. 张志芳. 方山柿控时催熟和品质控制研究[D]: [硕士学位论文]. 杭州: 浙江理工大学, 2019.
- 9. Sobajima, Y., Ishida, M. and Inaba, A. (1975) Studies on the Fruit Development of Japanese Persimmon (Diospyros kaki L.) II. Seed Abortion in Hiratanenashi. Journal of the Japanese Society for Horticultural Science, 44, 1-6.
- 10. Fukui, H., Mizuno, Y., Wakayama, Y., et al. (1993) Development of Abnormal Embryo Sac and Seed in Japanese Persimmon (Diospyros kaki) cv. Fuyu. Journal of the Japanese Society for Horticultural Science, 61, 773-778.
- 11. 扈惠灵, 曹永庆, 李壮, 等. 柿生殖生物学研究评述[J]. 中国农业科学, 2006, 39(12): 2557-2562.
- 12. Forbes, S.J., Mustiga, G., Romero, A., et al. (2019) Supplemental and Synchronized Pollination May Increase Yield in Cacao. HortScience, 54, 1718-1727. https://doi.org/10.21273/HORTSCI12852-18
- 13. Marafon, A., Herter, F., Hawerroth, F., et al. (2010) Occurrence Time and Intensity of Flower Bud Necrosis and Inflorescence Duplication in Pear Trees ‘Housui’ (Pyrus pyrifolia (Burm.) Nak.) during the Dormancy Period in Pelotas—RS. Brazil. Acta Horticulturae Sinica, 872, 97-100. https://doi.org/10.17660/ActaHortic.2010.872.10
- 14. Seif Aldin, D., Abdallah, F., Sizu, L., et al. (2019) Effects of Sucrose, Boric Acid, pH, and Incubation Time on in Vitro Germination of Pollen and Tube Growth of Chinese Fir (Cunnighamial lanceolata L.). Forests, 10, Article No. 102. https://doi.org/10.3390/f10020102
- 15. Du, G.C., Xu, J.G., Gao, C.R., et al. (2019) Effect of Low Storage Temperature on Pollen Viability of Fifteen Herbaceous Peonies. Biotechnology Reports, 21, e00309. https://doi.org/10.1016/j.btre.2019.e00309
- 16. Mondo, J.M., Agre, P.A., Asiedu, R., et al. (2021) Optimized Protocol for in Vitro Pollen Germination in Yam (Dioscorea spp.). Plants, 10, Article No. 795. https://doi.org/10.3390/plants10040795
- 17. Manchanda, P., Sharma, S., Marni, A., et al. (2022) Effects of Various Carbohydrates on the in Vitro Pollen Germination of Vinca rosea and Cucumis melo var. utilissimus. bioRxiv. https://doi.org/10.1101/2020.10.03.316216
- 18. Ma, Q.H., Wang, G.X., Chen, X., et al. (2013) Progress on Pollen-Stigma Compatibility in Corylus (Hazelnuts): A Review. Journal of Forestry Research, 24, 397-402. https://doi.org/10.1007/s11676-013-0372-7
- 19. Dhaliwal, A.S., Malik, C.P. and Singh, M.B. (1981) Overcoming Incompatibility in Brassica campestris L. by Carbon Dioxide and Dark Fixation of the Gas by Self- and Cross-Pollinated Pistils. Annals of Botany, 48, 227-234. https://doi.org/10.1093/oxfordjournals.aob.a086117
- 20. Tang, W.L., Zhao, Y.J., Zeng, J.J., et al. (2021) Integration of Small RNA and Transcriptome Sequencing Reveal the Roles of miR395 and ATP Sulfurylase in Developing Seeds of Chinese Kale. Frontiers in Plant Science, 12, Article ID: 778848. https://doi.org/10.3389/fpls.2021.778848
- 21. Zhao, Y., Chen, Z., Chen, J., et al. (2021) Comparative Tran-scriptomic Analyses of Glucosinolate Metabolic Genes during the Formation of Chinese Kale Seeds. BMC Plant Biology, 21, Article No. 394. https://doi.org/10.1186/s12870-021-03168-2
- 22. Barnabás, B., 陈周. 失水对玉米花粉萌发力的影响[J]. 国外农学–杂粮作物, 1985(4): 6-8.
- 23. 吴旺泽. 马铃薯花粉原生质体分离与培养[D]: [硕士学位论文]. 兰州: 甘肃农业大学, 2004.
- 24. 周瑞金, 扈惠灵, 李桂荣, 等. 甜柿花粉萌发及贮藏特性研究[J]. 中国农学通报, 2009, 25(2): 176-179.
- 25. 姜士博. 辽梅杏生殖生物学及繁殖技术研究[D]: [硕士学位论文]. 沈阳: 沈阳农业大学, 2020.
- 26. 詹术森, 陈思旭, 郭大勇, 等. 3种中国原产雄性柿种质花粉离体萌发条件优化及其活力快速检测[J]. 中国南方果树, 2020, 50(3): 107-110.
- 27. 张绍铃, 陈迪新, 康琅, 等. 培养基组分及pH值对梨花粉萌发和花粉管生长的影响[J]. 西北植物学报, 2005, 25(2): 225-230.
- 28. 顾翠花, 孙明哲, 王守先, 等. 3个紫薇品种花粉离体培养与萌发研究[J]. 安徽农业科学, 2011, 39(23): 14009-14011.
- 29. 秦晓君, 潘磊, 牛永志, 等. 烟草花粉活力检测方法筛选[J]. 云南大学学报, 2020, 42(3): 32-38.
- 30. 谷晓峰, 罗正荣. 甜柿巨大花粉萌发特征及辐射敏感性研究[J]. 武汉植物学研究, 2002(4): 280-282+328.
- 31. 朱悦凡. 天彭牡丹“红丹兰”不同外植体褐化防止及顶芽离体培养技术初步研究[D]: [硕士学位论文]. 雅安: 四川农业大学, 2012.
- 32. 扈惠灵, 李壮, 曹永庆, 等. 磨盘柿败育杂种胚胎发育的细胞学特性[J]. 果树学报, 2007, 24(5): 630-633.
- 33. 徐莉清, 张青林, 罗正荣. 幼胚发育阶段对次郎柿胚抢救效果的影响[J]. 果树学报, 2008, 25(1): 45-48.
NOTES
*通讯作者。