Advances in Microbiology
Vol.
09
No.
02
(
2020
), Article ID:
36244
,
8
pages
10.12677/AMB.2020.92011
Study on the Soil Persistence of Trichoderma harzianum and Bacillus velezensis
Zhendong Xu, Jianfeng Hu, Jiaqi Shen, Ying Wang, Changsheng Sun*
Zhejiang BioAsia Pharmaceutical Co., Ltd., Pinghu Zhejiang

Received: May 29th, 2020; accepted: Jun. 17th, 2020; published: Jun. 24th, 2020

ABSTRACT
Wheat head blight is caused by some Fusariums of Fusarium genus. It is a serious disease that widely exists in wheat planting areas all over the world. Our previous studies found that Trichoderma harzianum and Bacillus velezensis can significantly reduce the incidence rate of wheat head blight, but the duration of its control is unknown. Therefore, through this experiment, we explored the survival and persistence dynamics of these two biocontrol bacteria in soil, and found that Trichoderma harzianum and Bacillus velezensis can exist in soil for a long time. The survival rates of the Trichoderma harzianum and Bacillus velezensis were 29% and 52% respectively in the Nylon mesh cage experiment, which were significantly higher than 1.3% (p = 0.0007) and 3% (p = 1.18 × 10−7) of the field experiment, indicating that repeated application of pesticides was needed every year in order to obtain good control effect. After accumulating a certain amount of microorganism, the dosage and the times of control can be reduced properly.
Keywords:Trichoderma harzianum, Bacillus velezensis, Persistence

哈茨木霉及贝莱斯芽孢杆菌土壤宿存 探究
徐振栋,胡建峰,沈佳奇,王 莹,孙长胜*
浙江泛亚生物医药股份有限公司,浙江 平湖

收稿日期:2020年5月29日;录用日期:2020年6月17日;发布日期:2020年6月24日

摘 要
小麦赤霉病是由镰孢属多种镰刀菌侵染引起的,广泛存在于全世界小麦种植区的一种严重病害。我们此前的研究发现,哈茨木霉和贝莱斯芽孢杆菌在田间能显著减少小麦赤霉病的发病率,但对其防效的持续时间还未可知,因此,通过本次实验,对这两种生防菌在土壤中的生存情况和宿存动态进行探究,发现哈茨木霉和贝莱斯芽孢杆菌能在土壤中长期存在,尼龙纱网笼实验中的两种菌的宿存率分别为29%和52%,均显著高于大田实验的1.3% (p = 0.0007)和3% (p = 1.18 × 10−7),说明为了获得良好的防治效果初始年份每年都需要重复施药,待积累一定带菌量后,可适当减少用药量和防治次数。
关键词 :哈茨木霉,贝莱斯芽孢杆菌,宿存

Copyright © 2020 by author(s) and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/


1. 引言
小麦赤霉病是由镰孢属多种镰刀菌侵染引起的,广泛存在于全世界的小麦种植区的一种毁灭性病害 [1]。其在我国华南和长江中下游地区冬麦区以及北方春麦区普遍存在 [2]。我国近代曾多次发生大流行,给我国小麦产业造成了极大的损失,导致了我国江南部分地区减少甚至停止小麦的种植 [3]。目前我们主要使用化学农药对小麦赤霉病进行防治,而化学农药具有容易造成环境污染,病菌会产生抗药性等缺点,因此,越来越多人将目光聚焦到了生物防治上面。
目前,国内外对小麦赤霉病的微生物防治研究已经取得了很大的进展,发现了许多种细菌、真菌均具有小麦赤霉病防治的功能,主要有芽孢杆菌和木霉等 [4]。微生物防治应用的最大优点就是生防菌可以在土壤中定殖、宿存,当侵染源达到一定数量时就可获得自然控制病原微生物的能力。我们此前的研究发现哈茨木霉和贝莱斯芽孢杆菌在田间能显著减少小麦赤霉病的发病率,但我们对其防效的持续时间还未可知,因此我们通过本次实验,对这两种生防菌在土壤中的生存情况和宿存动态进行探究,定时取样,检查生防菌存活情况及动态,根据生防菌的动态,确定防治策略。
2. 材料与方法
2.1. 材料
2.1.1. 供试材料
哈茨木霉(Trichoderma harzianum) BACC0957菌株:由浙江泛亚生物医药股份有限公司提供;哈茨木霉孢子粉(250亿CFU/g):浙江泛亚生物医药股份有限公司实验室生产;贝莱斯芽孢杆菌(Bacillus velezensis) BACC01558菌株:由浙江泛亚生物医药股份有限公司提供;贝莱斯芽孢杆菌菌粉(100亿CFU/g):浙江泛亚生物医药股份有限公司实验室生产;实验土壤:于浙江泛亚医药股份有限公司绿化处采集。
2.1.2. 仪器与试剂
ZHJH-C1214B超净工作台:上海智城分析仪器制造有限公司;ZWYR-211D卧式脚踏开门型真彩触摸屏摇床:上海智城分析仪器制造有限公司;ME204电子天平:梅特勒–托利多国际贸易有限公司;HPX-300BSH-3恒温恒湿培养箱:上海新苗医疗器械制造有限公司。
LB固体培养基:葡萄糖10.0 g,酵母提取物10.0 g,胰蛋白胨10.0 g,氯化钠5.0 g,琼脂20 g,加蒸馏水1000 ml,121℃高压蒸汽灭菌20分钟;孟加拉红固体培养基:蛋白胨5.0 g,葡萄糖10.0 g,磷酸二氢钾1.0 g,硫酸镁(MgSO4∙7H2O) 0.5 g,琼脂20.0 g,孟加拉红0.033 g,氯霉素0.1 g,加蒸馏水1000 ml,121℃高压蒸汽灭菌20分钟。
2.2. 方法
2.2.1. 哈茨木霉及贝莱斯芽孢杆菌土壤存活实验
孟加拉红固体培养基、LB固体培养基、载玻片121℃高压蒸汽灭菌20分钟,待冷却至50℃~60℃,以无菌载玻片蘸取培养基,使载玻片两面涂满培养基,放置冷却,待培养基凝固后去除一面的培养基,仅保留单面培养基,制成孟加拉红玻片和LB玻片,各做16片。在孟加拉红玻片上接种哈茨木霉,在LB玻片上接种贝莱斯芽孢杆菌,分别于26℃和30℃下保湿培养3 d。
将一片尼龙纱网覆盖在载玻片上,两端用胶带粘上,然后将载玻片放入饭盒中,覆盖上菜园壤土中(土壤湿度以手捏成团,一拍即散为准),每种菌重复16个,保湿放置,并定期(第10天,第20天,1个月,2个月,4个月,6个月,8个月,10个月)抽出2片,将载玻片菌面置于相应培养基的平板上,哈茨木霉载玻片培养7天,贝莱斯芽孢杆菌培养2天,检查接种物的存活情况。
2.2.2. 混菌法探究哈茨木霉及贝莱斯芽孢杆菌在土壤中的动态变化情况
将接种物哈茨木霉、贝莱斯芽孢杆菌,接入菜园壤土(土壤自然晾干过100目筛,湿度以手捏成团,一拍即散为准),接种量为每100 g土样中加入2 g菌粉,混匀。混匀后装入20 cm × 10 cm × 10 cm的尼龙纱网笼中,埋入菜园土中,笼的上方与地表齐平。每隔一个月取样一次进行检测,每次取土5 g,烘干后打碎混匀,再从中取土1 g,溶于100 ml水中,摇床25℃,180 rpm震荡15分钟,随后取1 ml悬浊液加入9 ml蒸馏水试管中进行梯度稀释,稀释至10−8,各梯度稀释液分别吸取100 ul在相应的培养基上涂平板,各自培养合适时间,观察菌落数量,以菌落总数在10~30个为宜,确定一个合适的稀释浓度。首次确定合适稀释浓度后,后面一个月可根据上一个月的稀释浓度上下浮动1个梯度进行确认。选择合适的稀释浓度,稀释土壤,取100 ul稀释液涂平板(4~6个),各自培养合适的时间后计数哈茨木霉和贝莱斯芽孢杆菌的成菌落单位(CFU),取平均数。按下列公式计算单位土壤的菌落数:
单位土壤(g)中的菌落数(CFU/g) = 各平板菌落平均数 × 稀释倍数 × 10
2.2.3. 大田试验后土壤中哈茨木霉及贝莱斯芽孢杆菌动态变化
每隔一个月取大田土样一次进行检测,五点取样法每次取土50 g,烘干后打碎混匀,再从中取土1 g,溶于100 ml水中,摇床25℃,180 rpm震荡15分钟,随后取1 ml悬浊液加入9 ml蒸馏水试管中进行梯度稀释,稀释至10−7,各梯度稀释液分别吸取100 ul在相应的培养基上涂平板,各自培养合适时间,观察菌落数量,以菌落总数在10~30个为宜,确定一个合适的稀释浓度。首次确定合适稀释浓度后,后面一个月可根据上一个月的稀释浓度上下浮动1个梯度进行确认。选择合适的稀释浓度,稀释土壤,取100 ul稀释液涂平板(4~6个),各自培养合适的时间后计数哈茨木霉和贝莱斯芽孢杆菌的成菌落单位(CFU),取平均数。按下列公式计算单位土壤的菌落数:
单位土壤(g)中的菌落数(CFU/g) = 各平板菌落平均数 × 稀释倍数 × 10
2.2.4. 数据统计分析
实验数据使用Excel软件进行数据处理并制作菌落数曲线图。
3. 结果与分析
3.1. 哈茨木霉及贝莱斯芽孢杆菌土壤存活情况
在载玻片上接种哈茨木霉及贝莱斯芽孢杆菌,培养3 d后插入土壤中,定期检查存活情况,具体结果如表1所示。

Table 1. Survival of Trichoderma harzianum and Bacillus velezensis in soil
表1. 哈茨木霉及贝莱斯芽孢杆菌土壤存活情况
玻片实验结果显示,哈茨木霉和贝莱斯芽孢杆菌能在土壤中长期存在,而它们又能抑制镰刀菌生长,减少镰刀菌在土壤中的菌量,因此能对赤霉病有一个长期的抑制效果。
3.2. 哈茨木霉及贝莱斯芽孢杆菌在混菌土壤中的动态变化情况
采用混菌法进行实验,每100 g土壤中混入2 g菌粉,装入尼龙纱网笼中,埋入土中,每月检测结果如下表2和图1,图2所示。
Table 2. Changes of colony forming units of Trichoderma harzianum and Bacillus velezensis (mixed method)
表2. 哈茨木霉及贝莱斯芽孢杆菌菌量变化情况(混菌法)
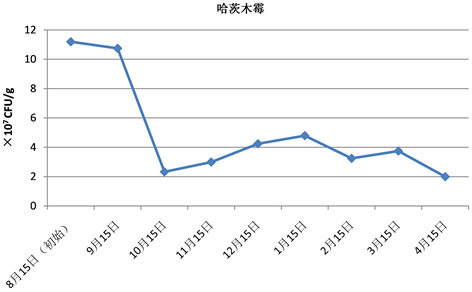
Figure 1. Changes of colony forming units of Trichoderma harzianum (mixed method)
图1. 哈茨木霉菌量变化情况(混菌法)
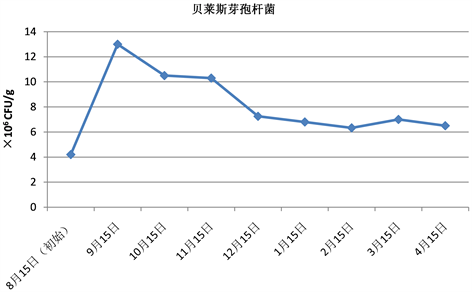
Figure 2. Changes of colony forming units of Bacillus velezensis (mixed method)
图2. 贝莱斯芽孢杆菌菌量变化情况(混菌法)
由表2和图2可知,混在土中的贝莱斯芽孢杆菌成菌落总数第一个月呈显著上升,上升约3倍,这可能是由于第一次检测时贝莱斯芽孢杆菌菌粉刚从冰箱取出,还未活化,导致首次检测成菌落单位数量偏少,埋于土中一个月后,贝莱斯芽孢杆菌活化,此时温、湿度均对该菌增殖有利,在充分利用土壤养分基础上增殖,成菌落总量出现显著上升。从第二个月开始,土壤养分逐步显得不足,而且此时江淮一带气温开始降低,该菌繁殖生长速率下降,土中贝莱斯芽孢杆菌的成菌落单位数逐渐下降,从12月开始贝莱斯芽孢杆菌养分消耗与枯枝落叶产生的养分相适应,成菌落单位稳定在6.8 × 106 CFU/g左右,约为最高菌量的52%。
由表2和图1可知,混在土中的哈茨木霉成菌落总数第一个月没有明显变化,第二个月出现大幅度下降,随后维持在3.3 × 107左右,约为初始菌落的29%。这可能是由于第一个月时哈茨木霉利用土壤养分增殖,增长与死亡平衡,菌量变化不大,而第二个月土壤养分消耗,哈茨木霉没有足够营养,所以菌体开始大量死亡,成菌落单位出现大幅下降,随后哈茨木霉在土壤中生长和繁殖消耗的养分和枯枝落叶逐渐产生的养分相适应,第3~第5个月时成菌落单位有缓慢增加,随后保持平稳波动。
贝莱斯芽孢杆菌最后的宿存率为50%,哈茨木霉最后的宿存率约为17.9%,表明混菌法中贝莱斯芽孢杆菌的宿存能力强于哈茨木霉。
3.3. 哈茨木霉及贝莱斯芽孢杆菌在大田土壤中的动态变化情况
Table 3. Changes of colony forming units of Trichoderma harzianum and Bacillus velezensis (field)
表3. 哈茨木霉及贝莱斯芽孢杆菌菌量变化情况(大田)
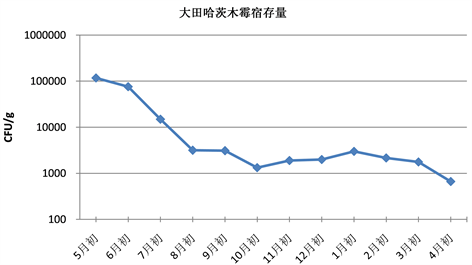
Figure 3. Changes of colony forming units of Trichoderma harzianum (field)
图3. 哈茨木霉菌量变化情况(大田)
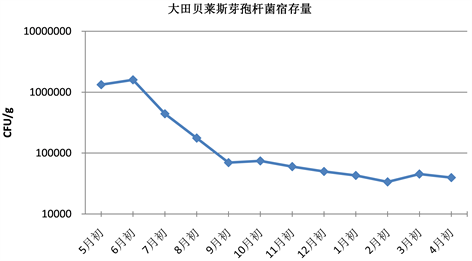
Figure 4. Changes of colony forming units of Bacillus velezensis (field)
图4. 贝莱斯芽孢杆菌菌量变化情况(大田)
如表3和图3所示,大田中哈茨木霉宿存量在施药后的第一个月略有下降,随后两个月有明显的下降,可能是由于第一个月消耗土壤积攒的养分进行增殖,养分大量消耗,同时哈茨木霉在高温下适应性差,最佳繁殖温度为25度,7~8月份气温较高,不利于哈茨木霉繁殖,增殖速度小于死亡速度,菌落数量下降。在之后的几个月里,哈茨木霉保持平稳波动状态,成菌落总数维持在1.5 × 103 CFU/g左右,约为初始检测量的1.3%。从10月份开始,哈茨木霉整体呈上升趋势,这可能是因为此时气温逐渐降低,以及秋季杂草枯萎腐烂,土壤有机质增加,为哈茨木霉生长提高了有利条件。到了3月份,哈茨木霉有较大的下降趋势,可能是由于春季植物复苏,大量消耗土壤营养水分,哈茨木霉可利用的营养变少,使得菌量减少。如表3及图4所示,大田中贝莱斯芽孢杆菌宿存量第一个月有所上升,之后三个月大幅下降,与土壤养分消耗有关。随后10月初至2月初总体呈缓慢下降的趋势,变化幅度不大,成菌落单位大致维持在4×104 CFU/g左右,约为初始检测量的3%。结果说明哈茨木霉和贝莱斯芽孢杆菌的菌落总数在下降到一定程度后,与大田的环境相适宜,增殖和死亡呈动态平衡。
贝莱斯芽孢杆菌一年后的宿存率为3%左右,哈茨木霉一年后的宿存率约为0.6%,说明贝莱斯芽孢杆菌在大田的生态适应性较强,宿存效果比哈茨木霉要强一些。但从最后结果来看都比初始菌量小很多,表明此时哈茨木霉及贝莱斯芽孢杆菌对小麦赤霉病的防治效果大幅度下降,需要再次施药以保证防效。
4. 结论与讨论
近几年来,木霉和芽孢杆菌作为拮抗菌,具有良好的生物防治功能的报道已有很多。而哈茨木霉和贝莱斯芽孢杆菌对镰刀菌的抑制作用也有报道 [5] [6]。
Hasan等 [7] 筛选了一种哈茨木霉(Trichoderma harzianum),用其进行了小麦赤霉病的大田防治试验,他们的试验结果表明,哈茨木霉有效地抑制了禾谷镰刀菌对小麦的侵染和发病,甚至在抑制镰刀菌引起苗腐率的同时促进了小麦的出苗率,提高了小麦31.58%的产量。哈茨木霉的抑菌机制可能是通过其寄生作用产生的,它首先分泌出了一些可以裂解细胞壁的水解酶,如几丁质酶和纤维素酶,通过这些酶裂解了镰刀菌的细胞壁,随后其侵入镰刀菌的细胞中吸收营养从而导致致病菌的死亡 [8]。
宗英等 [9] 筛选了一株贝莱斯芽孢杆菌JS25R,实验发现其对禾谷镰刀菌有抑制作用,并且在温室中进行了小麦赤霉病防治实验,显著降低了赤霉病发病率、发病程度和病情指数,结果表明其在温室中有较好的防治效果。贝莱斯芽孢杆菌的抑制机理可能是来自于自身的代谢产物,如一些降解酶类、抗菌蛋白、脂肽类抑菌物质及成分较为复杂的挥发性物质 [10] [11]。其能产生36种挥发性物质,其中2-十一烷酮、二甲基二硫醚、2-壬酮和2-壬醇能完全抑制禾谷镰刀菌的生长。
我们此前的研究发现,通过哈茨木霉和贝莱斯芽孢杆菌的联用,从多个方面抑制小麦赤霉病病原菌生长,能达到比单一使用更好的防治效果。
大田实验和尼龙纱网笼实验的哈茨木霉和贝莱斯芽孢杆菌宿存结果不一致,其中哈茨木霉在大田的宿存率约为1.3%,而在尼龙纱网笼的宿存率约为29%;贝莱斯芽孢杆菌在大田的宿存率约为3%,在尼龙纱网笼的宿存率约为52%,结果发现尼龙纱网笼实验的两种菌的宿存率均显著高于大田实验(p < 0.01),贝莱斯芽孢杆菌宿存率高于哈茨木霉。产生这种差异的原因可能是:1、尼龙纱网笼实验中每单位土壤的哈茨木霉和贝莱斯芽孢杆菌的含菌量比大田土壤高了很多,并且其使用的是混菌法,土壤与菌混合均匀,受环境影响小,营养利用充分;而大田是采用喷洒的方法,菌主要集中在表层,受环境(紫外线,降水等)的影响较大;2、大田土壤杂菌比尼龙网内土壤杂菌多很多,受各种杂菌不同方面的影响,导致大田中两种菌的宿存率显著低于尼龙网内;3、菜园土营养远大于大田土壤营养,生防菌的循环产孢能力强于大田;4、贝莱氏芽孢杆菌的生态适应能力高于哈茨木霉,所以土壤宿存力高于哈茨木霉。结果表明小麦赤霉病防治所用菌剂,初始每年均需重复施药,以保证较好的防治效果。待积累一定带菌量后,可适当减少用药量和防治次数。
文章引用
徐振栋,胡建峰,沈佳奇,王 莹,孙长胜. 哈茨木霉及贝莱斯芽孢杆菌土壤宿存探究
Study on the Soil Persistence of Trichoderma harzianum and Bacillus velezensis[J]. 微生物前沿, 2020, 09(02): 66-73. https://doi.org/10.12677/AMB.2020.92011
参考文献
- 1. Bal, G. (1994) Scab of Wheat: Prospects for Control. Plant Disease, 78, 760-766. https://doi.org/10.1094/PD-78-0760
- 2. 陈然, 李俊凯, 李黎, 等. 小麦赤霉病生物防治研究进展[J]. 河南农业科学, 2014, 43(12): 1-5.
- 3. 侯明生, 黄俊斌. 农业植物病理学[M]. 北京: 科学出版社, 2006: 68-73.
- 4. 陈文华, 殷宪超, 武德亮, 等. 小麦赤霉病生物防治研究进展[J]. 江苏农业科学. 2020, 48(4): 12-18.
- 5. 宋磊, 李雅乾, 陈捷, 等. 混合发酵木霉菌对4种病原真菌的拮抗作用[J]. 沈阳农业大学学报, 2012(3): 331-335.
- 6. 王纳贤. 尖孢镰刀菌拮抗细菌的筛选与抗菌蛋白的分离[D]: [硕士学位论文]. 保定: 河北大学, 2010.
- 7. Hasan, M.M., Rahman, S.M.E., Kim, G.H., et al. (2012) Antagonistic Potentiality of Trichoderma harzianum towards Seed-Borne Fungal Pathogens of Winter Wheat cv. Protiva in Vitro and in Vivo. Journal of Microbiology and Biotechnology, 22, 585-591. https://doi.org/10.4014/jmb.1107.07063
- 8. Mukherjee, M., Mukherjee, P.K., Horwitz, B.A., et al. (2012) Trichoderma-Plant-Pathogen Interactions: Advances in Genetics of Biological Control. Indian Journal of Microbiology, 52, 522-529. https://doi.org/10.1007/s12088-012-0308-5
- 9. 宗英, 赵月菊, 刘阳, 等. 一株贝莱斯芽孢杆菌抑制禾谷镰刀菌的研究[J]. 核农学报, 2018, 32(2): 310-317.
- 10. 孙冰冰, 李伟, 魏军, 等. 生防芽孢杆菌的研究进展[J]. 天津农业科学, 2015(12): 102-107.
- 11. Joshi, R. and Gardener, M.S. (2006) Identification and Characterization of Novel Genetic Markers Associated with Biological Control Activities in Bacillus subtilis. Phytopathology, 96, 145-154. https://doi.org/10.1094/PHYTO-96-0145
NOTES
*通讯作者。