DPrP21 Preamylase 1 (X77318.1) 2 L. vannamei 南美白虾 251 8.00E-32
DPrP24 ribosomal protein 4 (AY829788.1) 1 Lonom i a o blique 天蚕蛾幼虫 254 4.00E-42
DPrP27 ribosomal protein L8 (DQ316258.1) 1 L. vannamei 南美白虾 236 7.00E-64
DPrP31 S2 ribosomal protein (AY292479.1) 1 Eleginus gracilis 红花鳕鱼125 5.00E-30
The number in the third column indicates functional groups. 1: Ribosomal, 2: Metabolism, 3: Immune response, 4: Respiration, 5: Vision, 6: Signaling, 7: Structure, 8:
Translation, 9: Chromosome.
2.7. 统计分析
由于米虾非纯品系(outbred)动物,个体之间表现
型背景的差异性大,所以每一相对数值至少进行 5重
复,每一重复至少使用 5只虾子。以 one-way ANOVA
和Duncan进行不同浓度处理组之间的比较,分析后
有显着差异者(P < 0.05)。
3. 结果
3.1. 差异表现片段的扣减基因库(subtracted
cDNA library)的建立
为了得到 DPrP 处理前后具有差异异性表现之基
因,本研究由实验组及控制组的虾子,利用抑制性扣
减杂交法后,针对所有转型株,利用巢式 PCR 确认后,
筛选获得431 个转型株。进一步确认插入之基因片段
是否相同,以三种限制酶EcoRI、Bam HI 和HimdⅢ进
行切割,以分析各 cDNA 的限制图谱后,共挑选得 226
个转型株,即得到一个扣减后cDNA 基因库。
3.2. 基因片段的比对
基因库所有基因片段定经序后,以 DNAStar 软件
将不同片段序列上重复的部分进行前后顺序的排列
及重组,完成后得到共获得71 个独特的表达序列标
签(expressed sequence tages; ESTs)。接着,利用 NCBI
以短暂暴露于邻苯二甲酸二丙酯(Dipropyl Phthalate)之淡水多齿新米虾
(Neocaridina denticulata)建立生态毒理基因组生物标志物
Table 2. The specific primers of ESTs used in this study
表2. 使用在本研究之表达序列标签的专一性引子序列
EST ID Sequence for primers (5’3’) Size (bp)
Re9 L:CAGCCCAAATGTCCGTTACT
R:CCTTGAAGCTTGGCACATC 600
Re17 L:CGTGAGCACGTCTTGCTTT
R:TGAGCAGTGTGGCAGTCAA 701
Re18 L:GGCTCGTGTGTCAACATCC
R:CCCCCATTAGCAAGAGGAA 522
R20 L:CTTCAACCCAGCCAATGTG
R:GGTAGCAATGCCGCAGTTA 337
DPrP9 L:GCC TTGTTACGGTCGTGAGT
R:GGGAAGAAAGAAGGCTGACG 330
DPrP10 L:CTCATGTCCCAGAAACCAA
R:CCCAGGTAAAGAAGGAGTCA 248
DPrP11 L:CACACCTCGCGGGTAGTAGT
R:ACCAAGCTGCTGCTTCAAA 510
DPrP15 L:CCTCCAGGTCTTCTGGTGAG
R:GAAGCGCTTGAGTGTCACAG 375
uDPrP4 L:ATGTCGAGTGCCAGCTCAG
R:GTGTTGGCAGTGACGACTCA 315
uDPrP12 L:CTTCAGCAACTGCAAGTCCA
R:GCATGACTGTGAGGGCTTTT 356
uDPrP13 L:CACTAGGCGAGTTAAGGATG
R:GTTTCCTTTATGGTGCGAGT 254
uDPrP14 L:GCCGATGTCACGGATAAGTC
R:GGAGTCTGT GCCCAAGTCAT 304
EST, expressed sequence tag; uDPrP denotes ESTs had not be identified in the
public databases and are regarded as function unknown genes.
的BLASTN 和BLASTX分别进行核酸序列及胺基酸
序列的比对,将其中核酸序列比对所得之E值低于(或
等于)10−25 且比对到的物种为无脊椎动物之基因片
段,定义为已知生理功能;比对后得到 23 段ESTs 对
应为已知功能基因(表1)及48段ESTs为未知功能。
已知功能ESTs 大致可分与九类生理功能相关,包括:
核糖体组成的核糖体蛋白(ribosomal proteins;DPrP24,
DPrP27 及DPrP31);代谢相关基因的胰蛋白酶(trypsin;
Re4 及Re5)、胰凝乳蛋白酶(chymotrypsin; Re6)、淀 粉
脢原(preamylase; DPrP21)、溶脢体组织蛋白脢
(cathepsin-L-like cysteine peptidase; Re20)和磷酸烯醇
式丙酮酸羧激酶(Phosphoenolpyruvate carboxykinase;
DPrP10);防御相关的血青素(hemocyanin; Re9)、QM
蛋白(QM protein; DPrP9);呼吸相关的粒线体基因
(DPrP11)、粒线体16S rRNA (Re1及Re2)、血青素
(Re9)、细胞氧化酵素COI (cytochrome oxidase subunit
I; Re18)及COII (cytochrome oxidase subunit II;
DPrP16);视觉相关的视紫蛋白(opsin; Re12);讯号 传
递相关的携钙蛋白(sarcoplasmic calcium-binding pro-
tein; Re13);细胞和组织结构相关 的胶 原溶 解丝 氨酸
蛋白酶(Collagenolytic serine protease; Re19)和肌球蛋
白轻链(Myosin light chain; DPrP15);转译相关的延长
因子(elongation factor 1-alpha; Re17)与染色体相关基
因(Chromosome 4; DPrP1)等(表1)。
3.3. 不同浓度 DPrP 对目标基因的影响
确认上述得到的 ESTs 是否因 DPrP 处理而表现确
有差异,实验中选取不同已知功能的ESTs 和四个未
知功能 ESTs,以 RT-PCR半定量法,对暴露于不同浓
度DPrP 的米虾,进行这些目标基因之 mRNA 合成量
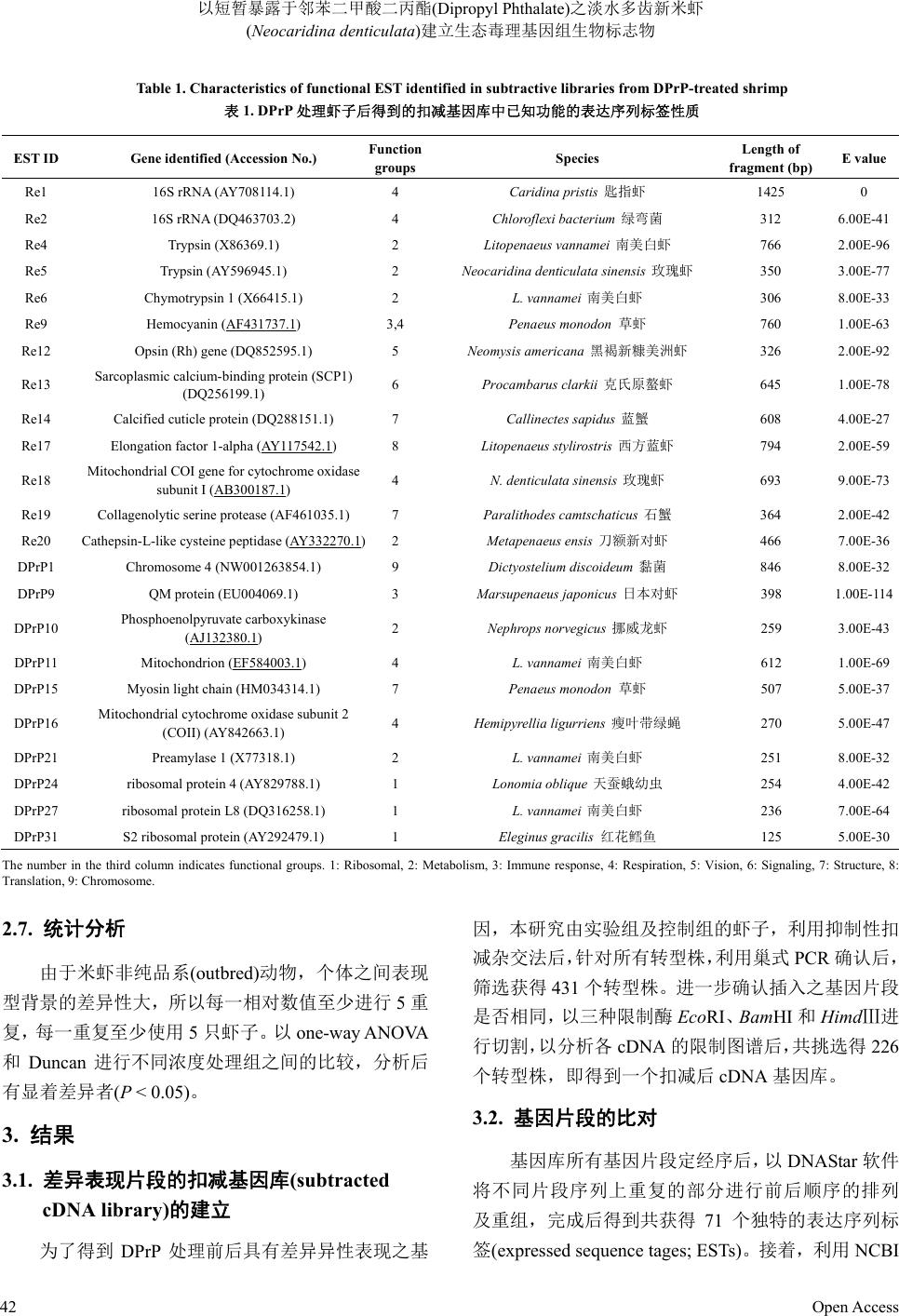
的分析。DPrP 影响免疫相关基因的结果如图 1所示,
浓度为 0.5~1.0 mg/L会明显增进血青素基因(hc)
mRNA 量,对MQ 基因(mq)则无明显影响;DPrP浓
度提高至50 mg/L 明显抑制两个基因的转录。对于代
谢相关基因,DPrP 于0.5 和1.0 mg/L 时会促进溶脢体
组织蛋白脢基因catl 的表现,浓度为50 mg/L 的DPrP
对此基因无影响;然而,基因磷酸烯醇式丙酮酸羧激
Re9
Hemocyanin
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
*
*
*
DPrP7
QM protein
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
*
0.51.0 50.0
Concentration (mg/L)
Figure 1. Messenger RNA expression of immunity-related genes of
Neocaridina denticulate treated with various concentrations of
dipropyl phthalate (DPrP) for one day
图1. 米虾曝露于不同浓度之邻苯二甲酸二丙酯一天后免疫相关基
因的讯息 RNA 表现量
Open Access 43
以短暂暴露于邻苯二甲酸二丙酯(Dipropyl Phthalate)之淡水多齿新米虾
(Neocaridina denticulata)建立生态毒理基因组生物标志物
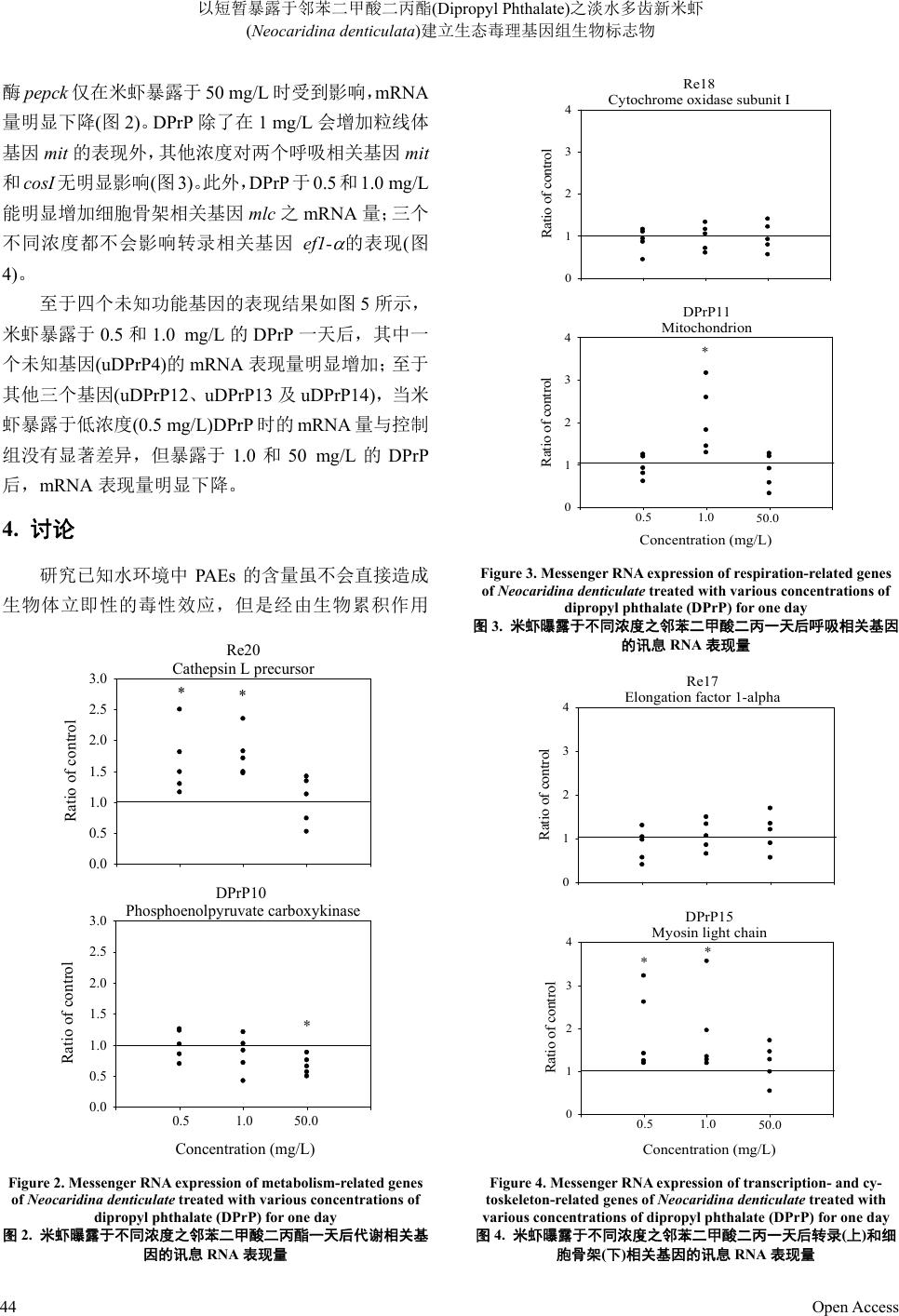
酶pepck 仅在米虾暴露于 50 mg/L时受到影响,mRN A
量明显下降(图2)。DPrP 除了在 1 mg/L 会增加粒线体
基因 mit 的表现外,其他浓度对两个呼吸相关基因mit
和cosI无明显影响(图3)。此外,DPrP 于0.5 和1.0 mg/L
能明显增加细胞骨架相关基因mlc 之mRNA 量;三个
不同浓度都不会影响转录相关基因 ef1-
的表现(图
4)。
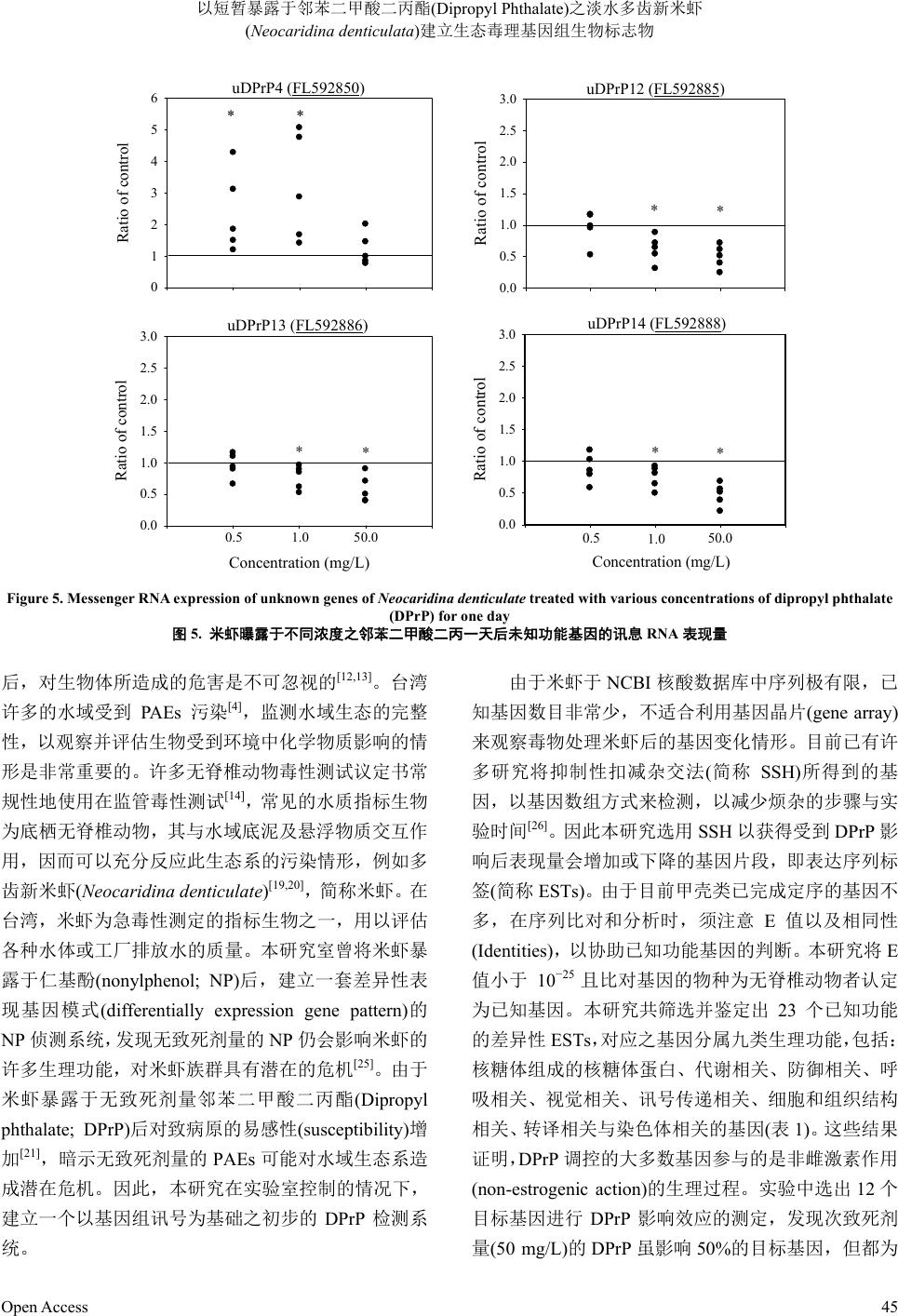
至于四个未知功能基因的表现结果如图 5所示,
米虾暴露于 0.5 和1.0 mg/L的DPrP 一天后,其中一
个未知基因(uDPrP4)的mRNA 表现量明显增加;至于
其他三个基因(uDPrP12、uDPrP13 及uDPrP14),当 米
虾暴露于低浓度(0.5 mg/L)DPrP时的 mRNA 量与控制
组没有显著差异,但暴露于1.0和50 mg/L的DPrP
后,mRNA 表现量明显下降。
4. 讨论
研究已知水环境中 PAEs的含量虽不会直接造成
生物体立即性的毒性效应,但是经由生物累积作用
DPrP10
Phosphoenolpyruvate carboxykinase
Concentration (mg/L)
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Re20
Cathepsin L precursor
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
**
*
0.51.0 50.0
Figure 2. Messenger RNA expression of metabolism-related genes
of Neocaridina denticulate tr eated with various concentrations of
dipropyl phthalate (DPrP) for one day
图2. 米虾曝露于不同浓度之邻苯二甲酸二丙酯一天后代谢相关基
因的讯息 RNA 表现量
Re18
Cytochrome oxidase subunit I
Ratio of control
0
1
2
3
4
DPrP11
Mitochondrion
Ratio of control
0
1
2
3
4
*
Concentration (mg/L)
0.51.050.0
Figure 3. Messenger RNA expression of respiration-related genes
of Neocaridina denticulate treated with various concentrations of
dipropyl phthalate (DPrP) for one day
图3. 米虾曝露于不同浓度之邻苯二甲酸二丙一天后呼吸相关基因
的讯息 RNA 表现量
Re17
Elongation factor 1-alpha
Ratio of control
0
1
2
3
4
DPrP15
Myosin light chain
0
1
2
3
4
*
Concentration (mg/L)
*
0.51.0 50.0
Ratio of control
Figure 4. Messenger RNA expression of transcription- and cy-
toskeleton-related genes of Neocaridina denticulate treated with
various concentrations of dipropyl phthalate (DPrP) for one day
图4. 米虾曝露于不同浓度之邻苯二甲酸二丙一天后转录(上)和细
胞骨架(下)相关基因的讯息 RNA表现量
Open Access
44
以短暂暴露于邻苯二甲酸二丙酯(Dipropyl Phthalate)之淡水多齿新米虾
(Neocaridina denticulata)建立生态毒理基因组生物标志物
Open Access 45
uDPrP4 (FL592850)
Ratio of control
0
1
2
3
4
5
6
**
uDPrP13 (FL592886)
Concentration (mg/L)
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0.5 1.0 50.0
*
*
uDPrP12 (FL592885)
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
*
*
uDPrP14 (FL592888)
Concentration (mg/L)
Ratio of control
0.0
0.5
1.0
1.5
2.0
2.5
3.0
**
0.5 1.0 50.0
Figure 5. Messenger RNA expression of unknown genes of Neocaridina denticulate treated with various concentrations of dipropyl phthalate
(DPrP) for one day
图5. 米虾曝露于不同浓度之邻苯二甲酸二丙一天后未知功能基因的讯息RNA表现量
后,对生物体所造成的危害是不可忽视的[12,13]。台湾
许多的水域受到 PAEs 污染[4],监测水域生态的完整
性,以观察并评估生物受到环境中化学物质影响的情
形是非常重要的。许多无脊椎动物毒性测试议定书常
规性地使用在监管毒性测试[14],常见的水质指标生物
为底栖无脊椎动物,其与水域底泥及悬浮物质交互作
用,因而可以充分反应此生态系的污染情形,例如多
齿新米虾(Neocaridina denticulate)[19,20],简 称 米 虾 。在
台湾,米虾为急毒性测定的指标生物之一,用以评估
各种水体或工厂排放水的质量。本研究室曾将米虾暴
露于仁基酚(nonylphenol; NP)后,建立一套差异性表
现基因模式(differentially expression gene pattern)的
NP 侦测系统,发现无致死剂量的 NP 仍会影响米虾的
许多生理功能,对米虾族群具有潜在的危机[25]。由于
米虾暴露于无致死剂量邻苯二甲酸二丙酯(Dipropyl
phthalate; DPrP)后对致病原的易感性(susceptibility)增
加[21],暗示无致死剂量的PAEs可能对水域生态系造
成潜在危机。因此,本研究在实验室控制的情况下,
建立一个以基因组讯号为基础之初步的 DPrP 检测系
统。
由于米虾于 NCBI 核酸数据库中序列极有限,已
知基因数目非常少,不适合利用基因晶片(gene array)
来观察毒物处理米虾后的基因变化情形。目前已有许
多研究将抑制性扣减杂交法(简称 SSH)所得到的基
因,以基因数组方式来检测,以减少烦杂的步骤与实
验时间[26]。因此本研究选用SSH 以获得受到 DPrP 影
响后表现量会增加或下降的基因片段,即表达序列标
签(简称 ESTs)。由于目前甲壳类已完成定序的基因不
多,在序列比对和分析时,须注意E值以及相同性
(Identities),以协助已知功能基因的判断。本研究将E
值小于 10−25 且比对基因的物种为无脊椎动物者认定
为已知基因。本研究共筛选并鉴定出 23个已知功能
的差异性ESTs,对应之基因分属九类生理功能,包括:
核糖体组成的核糖体蛋白、代谢相关、防御相关、呼
吸相关、视觉相关、讯号传递相关、细胞和组织结构
相关、转译相关与染色体相关的基因(表1)。这 些结 果
证明,DPrP 调控的大多数基因参与的是非雌激素作用
(non-estrogenic action)的生理过程。实验中选出 12 个
目标基因进行 DPrP 影响效应的测定,发现次致死剂
量(50 mg/L)的DPrP 虽影响50%的目标基因,但都为
以短暂暴露于邻苯二甲酸二丙酯(Dipropyl Phthalate)之淡水多齿新米虾
(Neocaridina denticulata)建立生态毒理基因组生物标志物
向下调节效应(down-regulation effect),显示对米虾生
理作用产生相当的压迫。当暴露剂量降低至无致死剂
量的 1 mg/L时,67%目标基因受到 DPrP 的上调或下
调,显示影响米虾的生理层面广;若暴露于更低浓度
0.5 mg/L 时,仅33%目标基因受到影响,且均为上调
效应(up-regulation effect)。这些无致死剂量 DPrP 对米
虾基因的影响结果无法判断是否对米虾的生理功能
具有压迫性,需进一步探讨。此外,米虾暴露在本实
验使用的三种次致死或无致死浓度之DPrP 后,在 12
个选定的目标基因中,讯号传递相关的 ef1-
和呼吸
相关的 cosI两个基因完全不受到影响;但其他主要参
与免疫(hc、qm和catl)和代谢(catl 和pepck)相关的基
因则对不同浓度DPrP 有不同反应(表3)。已知米虾暴
露于无致死剂量邻 DPrP 后的易感性(susceptibility)增
加[21],结合本研究结果推测,水体出现无致死剂量的
PAEs 可能主要经由防御或代谢路径影响米虾族群的
存活。
对大多数的甲壳类而言,污染物造成的环境压力
似乎是引起免疫力下降及患病率增加的重要因子之
一[27,28]。先前的研究显示,PAEs会降低米虾和淡水长
脚大虾的免疫活性和增加易感性[21,23];本研究证明 hc
及qm的表现确实会受 DPrP 调节,尤其高浓度 DPrP
会降低两个基因的表现。在节肢动物,血清素
(hemocyanin; Hc)是在肝胰脏合成[29,30],且为六聚合体
(hexamer)的氧气载体(oxygen carrier)。除了携带氧气
的功能外,许多研究也证明 Hc具有防御功能,例如
蜘蛛(Eurypelma californicum)[31-34]和鲎(Tachypleus-
tridentatus)[35]的Hc 经特定蛋白质水解后具有酚氧化
酵素(phenoloxidase)活性,且甲壳类的 Hc 经蛋白酵素
分解后,产生具抗微生物活性的抗微生物胜肽
(antimicrobial peptides)[36,37]。另一个免疫相关基因qm
的产物 QM 蛋白质,其在哺乳类和酵母菌细胞是一种
核糖体蛋白质[38,39];更多证据发现,QM 类蛋白
(QM-like protein)在细胞生长和分化等时期具有核糖
体蛋白质以外的功能[40,41]。Xu 等人以 SSH 方法分析
得知,白点病毒感染之日本对虾(Penaeus japonicas)
体内的 QM表现大量增加,且能够与Hc和肌球蛋白
(myosin)形成复合物而参与原酚氧化酵素系统(简称
proPO系统)和调节 Hc的酚氧化酵素活性,显示 QM
在节肢动物免疫系统具有特殊功能[42]。本研究室先前
Table 3.The summary of the effect of dipropyl phthalate (DPrP) on
the EST expression of Neocaridina denticulata
表3. 不同浓度邻苯二甲酸二丙酯基对米虾表达序列标签表现的影
响效应之摘要
DPrP (mg/L)
EST ID Target genes
0.5 1.0 50
Immunity
Re9 Hemocyanin (hc) E E D
DPrP9 QM protein (qm) N N D
Metabolism
R20 Cathepsin-L-like cysteine
peptidase (catl) E E N
DPrP10 Phosphoenolpyruvate
carboxykinase (pepck) N N D
Translation
Re17 Elongation factor 1-alpha (ef1-
) N N N
Respiration
Re18 Mitochondrial cytochrome
oxidase subunit I (cos I) N N N
DPrP11 Mitochondrion (mit) N E N
Cytoskeleton
DPrP15 Myosin light chain (mlc) E E N
Unknown function
uDPrP4 E E N
uDPrP12 N D D
uDPrP13 N D D
uDPrP14 N D D
Abbreviations are as follows: E = an enhancement effect of NP on gene expres-
sion; D = a reduction effect of NP gene expression; and N = no difference of gene
expression in DPrP-treated individuals from that in corresponding control.
研究指出,DPrP会影响米虾的超氧岐化酵素(super-
oxide dismutase; SOD)和原酚氧化酵素(prophenoloxi-
dase)等免疫酵素活性[21]。本研究结果推测,DPrP 也
可能透过影响hc和qm 的表现而而参与 proPO 系统以
调控米虾的免疫力。
细胞自溶酵素(cathepsin L; CatL)属于硫醇型蛋白
质分解酵素(cysteine protease)家族,多存在于胞内溶
体(lysosome)中,广泛的分布于正常组织,在发育过
程扮演重要角色,主要参与分化、原肠型成、脱皮及
卵黄利用等作用[43-49]。在无脊椎动物,硫醇型蛋白质
分解酵素与细胞内蛋白质的分解有关[50]。在甲壳类,
Open Access
46
以短暂暴露于邻苯二甲酸二丙酯(Dipropyl Phthalate)之淡水多齿新米虾
(Neocaridina denticulata)建立生态毒理基因组生物标志物
CatL 主要存在于肝胰脏,参与食物的分解,例如南美
白对虾(Litopenaeus vannamei)[51]和刀额新对虾 (Meta-
penaeus ensi s)[52]。近年研究证明,CatL 参与无脊椎动
物的先天性免疫力,包括南美白对虾[53]、中国对虾
(Fenneropenaeus chinensis)[54]、中华绒螯蟹(Eriocheir
sinensis)[55]和合浦珠母贝(Pinctada fucata)[56]等。根据
这些研究推测,DPrP 对米虾的防御反应或代谢活性具
有多重调节效应(multi-modulating effect)。相同的现象
也发生在NP 对米虾的影响,但是调节效应相反[25]。
磷酸烯醇式丙酮酸羧激酶(Phosphoenolpyruvate
carboxykinase)是醣质新生(gluconeogenesis)过程的重
要酵素,负责将草醋酸(oxaloacetate)转换成磷酸烯醇
丙酮酸(phosphoenolpyruv ate)和二氧化碳,以供后续转
换为葡萄糖。在某些特定的组织或细胞中相当重要,
例如大脑、肾脏髓质等皆以葡萄糖作为唯一能量来
源,当葡萄糖量不足时,就必须以醣质新生合成葡萄
糖来维持正常生理。本研究发现此酵素基因 pepck的
表现仅在高浓度时会受到抑制,推测次致死剂量可能
影响葡萄糖的恒定而对米虾造成一定程度的压迫。
综合以上暴露于 DPrP 后差异性表现基因的表现
結果证明,DPrP 主要经由多重生理功能的改变对米虾
产生外生毒力(ectotoxigenicity),推测无致死剂量 PAE
的污染仍可能危害到米虾族群的生存。由于台湾的环
境保护署使用米虾以监测水体的急毒性,结合本研究
的结果建议,米虾确实可作为水域指针系统(aquatic
indicator system)以评估水体污染对水生环境或生物的
风险,且发展自米虾的差异性表现ESTs未来应具有
作为基因组生物标志(genomic biomarkers)的潜力。由
于本研究获得的有效 ESTs 种类有限,因此未来仍须
搜集和鉴定更多污染相关的专一性基因,并整合更多
针对不同毒物的差异反应信息,以建立一套更具灵敏
度及可信度的监测模式组。
5. 致谢
本研究感谢台湾的国家科学委员会之计划经费
支持(NSC96-2745-B-031-003-URD)。
参考文献 (References)
[1]Holmes, P., Harrison, P., Bergman, A., Brouwer, I., Brouwer, B.,
Keiding, N., Randall, G., Sharpe, R., Skakkebaek, N., Ashby, J.,
Barlow, S., Dickerson, R., Humfrey, C. and Smith, L.M. (1997)
European workshop on the impact of endocrine disrupters on
human and wildlife. Proceedings of a Workshop, Weybridge, UK,
Report NO. EUR 17549.
[2]Staples, C.A., Peterson, D.R., Parkerton, T.F. and Adams, W.J.
(1997) The environmental fate of phthalate esters: A literature
review. Chemosphere, 35, 667-749.
[3]Yin, M.C. and Su, K.H. (1996) Investigation on risk of phthalate
ester in drinking water and marketed foods. Journal of Food and
Drug A n al ys is , 4, 313-318.
[4]Liu, C., Wang, S.K. and Lu, Y.B. (2000) Chemical characteriza-
tion of Tamshui River sediment in northern Taiwan. Journal of
Food and Drug Analysis, 76, 205-218.
[5]Chang, B.V., Liao, C.S., Huang, B.B. and Lee, C.C. (2004)
Occurrence of phthalate esters in Taiwan. Proceeding of the
Third Conference on Environmental Hormones and POPs, Tai-
pei, 12 November 2004, 63-66.
[6]Eckardt, R.E. (1973) Recent developments in industrial car-
cinogens. Journal of Occupational Medicine, 15, 904-907.
[7]Tavares, I.A. and Vine, N.D. (1985) Phthalic acid esters inhibit
arachidonate metabolism by rat peritoneal leucocytes. Journal of