设为首页
加入收藏
期刊导航
网站地图
首页
期刊
数学与物理
地球与环境
信息通讯
经济与管理
生命科学
工程技术
医药卫生
人文社科
化学与材料
会议
合作
新闻
我们
招聘
千人智库
我要投搞
办刊
期刊菜单
●领域
●编委
●投稿须知
●最新文章
●检索
●投稿
文章导航
●Abstract
●Full-Text PDF
●Full-Text HTML
●Full-Text ePUB
●Linked References
●How to Cite this Article
Botanical Research
植物学研究
, 201
4
,
3
,
32
-
40
http://dx.doi.org/10.12677/br.2014.3100
7
Published Online
January
201
4 (
http://www.hanspub.org/journal/br
.html
)
Application of
n
rDNA
-
ITS Seque nces i n
Pl ant Phylogeny
and Evolution
Xihong Liu, Lei Zhang, Gang Li, Rui Qin, Hong Liu
*
Eng i neeri ng R esearch Centre for the Protection an d Utiliza t ion o f Bioresour ce in Ethnic Area of Southern Ch ina, Co l lege of Life Science,
South
-
Central University for Nationalities, Wuhan
Email:
*
liuhong@mail.scuec.edu.cn
Received: Dec
.
2
nd
, 2013; revised: Dec
.
24
th
, 2013;
accepted
: Jan.
2
nd
, 201
4
Copyrigh t © 201
4
Xihon g Liu
et al
. This is an open access article distributed under the Creative Commons Attribution License, which
permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In
accordance of the
Creative Commons Attribution License all Copyrights © 201
4
a re reserve d for Hans and th e owner of the int ellectu al propert y
Xihong
Liu
et al
. All Cop yright © 201
4
are guarded by l
a
w and by Hans as a guardian.
Abstract:
The application of
nuclear rDNA ITS sequences in plant phylogeny and evolution
analysis
was
discussed in this paper. Nuclear rDNA is an important nuclear gene fragment
,
which had been
wi dely
used
in
phylogenetic
studies.
The internal transcribed spacer of nuclear rDNA was d
ivided into ITS1 and ITS2 by
5.8S rDNA. The length of ITS1 ranges from 165
bp to 298
bp in an giosp er ms, a nd t he le ngt h o f IT S2 r anges
from 177
bp to 266
bp. IT S
sequence
is relatively longer
in
gymnosperms. It
much d epends o n the length o f
ITS1.
As
mutate
quickly, ITS sequences are capable of providing many variable sites and informative sites,
which has become an important molecular marker
in plant systematic development and classification r
e-
searches within low
-
order angiosperm categories.
AT the sa me ti me
, it
can prov
e
importa nt systematic in fo
r-
mation for exploring polyploidy reticulate evolution and allopo lyploid origins.
Keywords:
ITS
Sequence
; Phylogenetic Development; Angiosperm; Gymnospe rm;
Advanced S t ructure
n
rDNA
-
ITS
区序列在植物系统与进化研究中的应用
刘锡红,张
磊,李
刚,覃
瑞,刘
虹
∗
中南民族大学生命科学学院,南方少数民族地区生物资源保护与综合利用工程中心,武汉
Email:
*
liuhong@mail.scuec.edu.cn
收稿日期:
2013
年
12
月
2
日;修回日期:
2013
年
12
月
24
日;录用日期:
201
4
年
1
月
2
日
摘
要:
本文就核
rDNA
(
nrDNA
)
的
ITS
区序列在植物系统与进化研究中的应用进行了讨论。
nrDNA
是植物系统发育研究中广泛应用的重要核基因片段。
5.8S
rDNA
把
nrDNA
的内转录间隔区分为
ITS1
和
ITS2
两部分。被子植物中
ITS1
的长度为
165
~
298
bp
,
ITS2
的长度为
177
~
266
bp
;裸子植物中
ITS
片段较长,且其长度变化主要由
ITS1
的长度变异所致。由于
ITS
序列变异较快,能够提供较丰富的
变异位点和信息位点,已成为被子植物较低分类阶元的系统发育和分类研究中的重要分子标记,为探
讨多倍体复合体网状进化关系,异源多倍体的起源提供了重要的系统学信息。
关键词:
ITS
序列;系统发育;裸子植物;被子植物;高级结构
*
通讯作者。
OPEN ACCESS
32
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
1.
引言
植物系统进化一直是生物科学研究的重要内容,
同时也是植物种质资源鉴定研究的重要领域,但由于
基因组结构的复杂性,给研究带来了一定的困难。近
年来,
ITS
区在植物系统进化中得到越来越多的应用。
由于
ITS
序列变异较快,能够提供较丰富的变异位点
和信息位点,已成为被子植物较低分类阶元的系统发
育和分类研究中的重要分子标记,也为探讨多倍体复
合体网状进化关系,异源多倍体的起源提供了重要的
系统学信息
[1]
。本文主要针对
ITS
序列的特殊性及其
优点,对其基本结构、功能、应用、测序与二级结构
预测展开了讨论。
研究表明,
18S
、
rbcL
等编码基因及线粒体
DNA
(
mtDNA
)
一般适用于较高分类阶元甚至整个种子植物
谱系间的系统发育的探讨,而
ITS
及叶绿体
DNA
(
cpDNA
)
的非编码区序列等因进化速率较快多用于较
低分类阶元的
系统关系研究。作为核糖体
D
NA
(
rDNA
)
间隔序列的
ITS
序列在目前的研究中发现其在基因组
中保持高度的一致性,并呈现一定的保守性。
ITS
序
列与中高度重复序列和非编码序列相比较,保守性较
高;与编码基因序列相比,变异速度相对较快。根据
以上的研究特点,可以推断出它们在进化上的一些关
系。目前,这方面的研究在裸子植物和被子植物上得
到广泛的应用。
目前对植种亲缘关系的研究方法有很多,例如杂
交法。从整体上看,遗传多样性的研究方法从个体形
态学水平、细胞学水平、生理生化水平发展到了分子
水平,研究层次也随之深入
[1]
。
ITS
最初由
White
等
利用真菌的
nRNA
序列设计出通用的引物和扩增方法
而引入系统学。因为
ITS
所具有的一系列特征而使它
在系统学中被广泛应用
[2]
。首先,
ITS
和其它的核
rDNA
(
nrDNA
)
多基因家族成员一样,核染色体组中高
度重复,成千上百的
nrDNA
拷贝以串联重复的方式
排列在染色体的一个或多个位点上,便于扩增、克隆
与测序。其次,
nrDNA
基因家族有着快速的致同进化,
从而促进染色体组重复单元之间,甚至是非同源染色
体的
nrDNA
位点之间的一致性,这个特点对系统重
建尤为重要。第三,
ITS
片段的长度通常较短
(
在被子
植物中小于
700
bp)
,两个间隔区的侧翼都是高度保守
的序列,容易利用设计的通用引物对多数群体进行扩
增。
不同植物的基因组结构和功能存在差异,导致进
化速率不同。在植物系统发育和进化的研究中,应用
较为广泛的是核基因组和叶绿体基因组。其中核基因
组进化的最快,约为叶绿体基因组的
2
倍。由于
cpDNA
的进化速率远不及
nDNA
,从而限制其在较低
分类阶元
(
如属、亚属
)
中的应用。因此,越来越多的
研究者将注意力集中到
nDNA
中进化较快的
DNA
序
列上,
18S
~
26S
核核糖体
DNA
的内转录间隔区
ITS
正是符合要求的序列之一
[3]
。因此,分析和研究不同
植种的
ITS
序列,可以通过建立系统树的方法将不同
的物种区分开来。
2. nrDNA
的
ITS
序列
2.1. 18S~
26S nrDNA
结构特征
编码
rRNA
的
rDNA
成簇存在于染色体的核仁组
织区中,是基因组
DNA
中的中等重复序列,构成有
转录活性的基因家族。在植物基因组中,每个
rDNA
重复单位的长度约为
7.8
kb
到
18.5
kb
,单倍体中的拷
贝数约为
600
~
8500
个。每个
rDNA
重复序列由保守
的编码区和可变的间隔区
IGS
构成。
rDNA
基本结构如图
l
所示,由
l8SrDNA
、
5.8S
rDNA
、
26S
rDNA
和位
于三者之间的内转录间隔区
ITS
组成,其中
ITS
被
5.8S
分为
ITS1
和
ITS2
两部分。
ITS1
、
ITS2
分别位于
l8S
~
5.8S
、
5.8S
~
26S
rDNA
之间,
而
18S
、
5.8S
、
26S
rDNA
的序列非常保守,这样就可
以用与它们序列互补的通用引物对
ITS
区进行
PCR
扩增和测序。
PCR
扩增引物的设计非常重要,目前研
究中所用引物多是根据
white
等设计的或对已发表的
引物序列作适当调整
(
图
1)
。
真核生物
rDNA
的编码区包括
18S
、
5.8S
和
28S
rDNA
(
植物中为
26S
rDNA
)
,它们组成一个转录单元,
产生一个
45S
的前体
RNA
,
45S
的前体
RNA
迅速折
叠,形成茎环状二级结构,为加工前体转录本成为成
熟
rRNA
及降解
ITS2
和
ITS2
部分为核苷酸的大分子
复合物提供必要的识别、接触和作用的信号
[4-6]
。外 部
转录间隔区
(ETS)
存在于
18S
rDNA
基因的上游和
28S
rDNA
基因的下游。
ITS
和
ETS
中都包含有前体
rDNA
加工的信息,在
rRNA
成熟的过程中起着相当重要的
作用
[7]
。其中一个特异性的剪切就发生在
ITS2
序列内
OPEN ACCESS
33
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
Figure 1.
The stru ct ure o f 18S
-
26S
rDNA in plants
图
1.
植物
18S~26S
rDNA
的基本结构,箭头示
PCR
扩增和测序引物
部,导致成熟的
5.8S3’
端和大亚基
LSU5’
端的最终形
成。
ITS
区和
ETS
区的转录产物均在
rRNA
成熟过程
中被降解。它们的转录产物在加工过程中被剪切掉,
只是部分地对成熟起作用
[8]
。在两段
ETS
之间的即是
非转录区
(
NTS
)
,它们共同构成了
IGS
,使两个相邻的
重复单位被隔开。
IGS
含有多个重复序列,有的序列
可以为稳定的转录提供起始位点,有的可以作为结核
转录所需蛋白的启动子
[9]
。这些重复序列的长度及重
复次数都导致了
IGS
序列的长度不仅在种间、种内,
甚至在一个群体的不同个体间都是高度可变的。
高等植物中
nrDNA
是高度重复的串联序列单位,
l8S
、
5.8S
和
26S
连接在一起,共同作为一个转录单
位,又称顺反子。而
5S
则位于另外的位点上,与
nrDNA
在染色体上的分布彼此独。
l8S
~
26S
rDNA
在
植物上有一至数个位点,高度重复
(
几百至几千次
)
,
由于协同进化,绝大多数物种内这些重复单位问已发
生纯合或接近纯合。如果植物多倍化历史不长,那么
同步进化尚未使
rDNA
的重复单位发生一致化,其
ITS
区就会具有很高的异质性,这种现象在禾本科偃
麦草等植物中已被证实存在。在自然界的植物尤其是
多倍体植物进化中,协同进化的现象很明显
[10]
。多 倍
体植物
ITS
区的进化途径十分复杂,祖先的
ITS
序列
可能在后代中共存 ,也 可 能只向 一个方向进化
[
11
,12]
。
ITS
作为在分子生物学中的研究重点的同时,也在种
质资源鉴定中得到了大量的应用,主要源于两个因素:
首先,
ITS
在核基因组中中高度重复,并且通过基因
转换和不等交换,这些重复单位间已经发生了位点内
和位点间的同步进化,即不同
ITS
拷贝间的序列趋于
相近或完全一致
[13]
。即不同
ITS
拷贝间的序列趋于相
近或完全一致,这就为对
PCR
产物直接进行测序奠
定了理论基础
[14]
。其次,
ITS
序列包含被
5.8S rDNA
所分隔的
ITS1
和
ITS2
两个片段
。
DNA
测序的难易
程度与成本和
DNA
片段长度有着直接的关系。在裸
子植物中
,
ITS
序列长度变异较大
[15]
,
不利于扩增和
测序
。
在被子植物中
,
ITS
序列的长度稳定
,
而且序
列变异较快
,
可以提供丰富的信息位点,测序方便。
2.2.
ITS
序列在被子植物鉴定中的应用
在被子植物中,
ITS
长度
(
包括
ITS1
、
ITS2
和
5.8S
rDNA
)
通常为
565
~
700
bp
,变异幅度很窄;既具有核
苷酸序列的高度变异性又有长度上的保守性,说明这
些间隔区的序列很容易在近缘类群间排序,而且丰富
的变异可在较低的分类阶元上解决植物的分子鉴定
及系统发育问题;并且,只需用扩增时的一对引物即
可对被子植物的
ITS
进行全序列测定,这使得该序列
在探讨被子植物属下水平的系统分类与进化的研究
中得到广泛应用。
被子植物核
rDNA
的
ITS
区中,
5.8S
rDNA
的长
度非常保守,一般为
163
bp
或
164
bp
,仅在大豆属中
发现其长度为
168
bp
[16]
,序列保守,在有些类群中根
本没有变异
[17]
。因此,
5.8S
rDNA
提供的信息有限,
在研究中可不必测该片段的序列
[18]
。但近几年也有研
究表明,高度保守的
5.8S
rRNA
基因序列对揭示远缘
属间的系统发育关系能提供一定的信息量,在研究中
可将
5.8S
与
ITS
序列结合起来
[19]
。
ITS1
和
ITS2
的长度比较保守,被子植物
ITS1
的
长度为
187
~
298
bp
,
ITS2
的长度为
187
~
252
bp
。至 于
ITS1
与
ITS2
的相对长度,不同类群间变化较大
[20]
,
OPEN ACCESS
34
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
在菊科、十字花科等科中,
ITS1
比
ITS2
长;在葫芦
科、玄参科等科中,
ITS1
比
ITS2
短;在蔷薇科、蝶
形花科等科中,
ITS1
和
ITS2
的长短因属、种而异。
在大多数被子植物类群中这两个片段所提供的信息
量相近,虽然单独根据
ITS1
或
ITS2
均可得出重要的
系统学结论
[21]
,但考虑到这两个片段的长度有限,各
自的信息量并不十分充足,因此,大多数情况下都是
将这两个片段综合起来考虑。
2.2.1
.
在科、亚科、族水平的应用
ITS
序列分析已成为在分子水平上探讨科、亚科、
族内关系十分有效的手段,在种内,
ITS
序列对于揭
示异域分布或间断分布居群间的关系具有很大潜力
,
该片段特别适合于属、种级的系统发育和分类研究。
汪小全等通过测定苦苣科
4
族
5
个属的
nrDNA
的
ITS
区以及部分
5.8S
rRNA
基因的
3’
端序列,,对其中
5
种代表植物进行了系统学研究,发现尖舌苣苔族的代
表植物在
ITS1
和
ITS2
在长度、序列以及
GC
含量上
均与其它几个族有较大差异,由此推测尖舌苣苔族可
能很早就沿单独的一个分支演化
[22]
。
章群和施苏华等对金缕梅科部分植物的
nrDNA
的
ITS
(
包含
5.8S
区
)
进行了比较分析,深入探讨了金
缕梅科的系统发育
[23,24]
。形态和分子证据表明,五味
子科是被子植物最基部的分支之一。而在传统的分类
系统中,五味子科一直被认为是木兰亚纲中较特化的
类群。两种截然不同的观点导致五味子科的系统位置
和在被子植物起源演化上的意义也截然不同,使得该
类群成为被子植物系统学研究中的关键类群之一而
备受关注。刘忠、汪小全等分析了五味子科内
14
种
代表植物的
nrDNA
的
ITS
序列,为五味子科的系统
位置和演化趋势提供了分子证据
[25]
。
2.2.2
.
在属及属下分类阶元中的应用
由于
ITS
序列在关系密切的物种间的长度特别接
近,而且序列又存在一定程度的变异,因此
ITS
序列
非常适合在属及属下水平做关于分类鉴定和系统发
育的研究。
Alice
与
Campell
对蔷薇亚科的悬钩子属
(
Rubus
)
进行了
nrDNA
ITS
区序列分析与系统学研究,
分析结果显示,
ITS
序列在悬钩子属下亚属水平上可
提供较大的信息量,但同时发现,在关系较近的近缘
种之间,
ITS
序列的变异性较低
[26]
。
保曙琳,丁小余等对野生菱和栽培菱种
rDNA IT S
片段进行分析,探讨了该片段在两大群体中的系统学
及鉴别研究意义,通过测序得到了
rDNA ITS1
、
ITS2
和
5.8S
rDNA
的完整序列,发现菱属
(
Trapa
)
各居群间
rDNA ITS
的差异较小,但是根据
ITS
序列的碱基插
入、缺失等特征可以很好地鉴别野生菱和栽培菱。这
说明对于某些类群,即使变异性较低,仍然可以根据
ITS
的其它特征对物种进行鉴定
[27]
。
ITS
序列分析已成为近年来国际上植物多样性研
究的热点,并在药用植物种质资源研究中得到广泛的
应用。丁小余等利用
rDNA ITS
序列对国内外石斛属
民族药用植物的分类及生药学研究进行了探讨,并鉴
别出了易混淆的石斛品种
[28]
;马小军等研究了野山参
(
Panax
)
和栽培参
rDNA IT S
的遗传差异,利用得到的
ITS
序列鉴别出不同的品种
[29]
;赵志礼、周开亚等用
直接测序法对国产黑果山姜
(
Zingiber
)
以及水山姜的
rDNA
ITS
序列进行了测定,结果显示两者序列完全
一致,证实了《中国植物志》中记载的水山姜为黑果
山姜
[30]
。以上的研究成果都说明,
rDNA ITS
序列是一
种适宜于中药材鉴别分子标记。对稻属
(
Oryza
)
[31]
、苋
属
(
Amaranthus
)
[32]
等其它植物物种的分类和系统学研
究也说明,
ITS
将在物种鉴别中发挥越来越重要的作用。
2.2.3
.
种及种下水平的应用
rDNA
上的
5.8
、
18
和
28S rRNA
基因有极大保
守性,即存在广泛的异源同源性。而由于
ITS
序列不
参与成熟核糖体的组装,所以
ITS
序列在进化过程中
承受的自然选择压力非常小,因此能够容纳更多的变
异。
ITS1
和
ITS2
属于中度保守序列,其保守性基本
表现为种内相对一致,种间差异比较明显。这种特点
使
ITS
序列适合于植物的分子鉴定以及属内物种间
或种内差异较明显的物种间的系统发育关系分析。同
时由于
ITS
在位点内、位点间存在同步进化,导致在
许多物种内
ITS
不存在位点多态性
[
33
-35]
,但仍有不少
研究发现
ITS
存在种内多态性,即在不同亚种、居群
的同一位点有
2
或
2
个以上的核苷酸类型
[36
-38]
,有 些
种内多态性还可在种、亚种间关系的分析中得到应用,
如
Ainouche
等人发现在禾本科雀麦属植物中,虽然
大麦状雀麦
(
Bromus
hordeaceus
)(
4x
=
28
)
等
4
种植物
在不同亚种、居群间的
ITS
序列完全一致,但除此以
外,发现在有些物种的亚种、居群间仍存在位点多态
OPEN ACCESS
35
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
性,多态位点总数为
27
个,主要集中在四倍体物种
B.
lanceolatus
,
B.
secalinus
,
B.
grossus
和
B.
br
o-
moideus
之中
[39]
。同 时
Ainouche
等人在雀麦属的几个
二倍体物种中,如
B.
brizif ormis
,
B.
japonicus
,
B.
intermedius
也发现了
1~4
个多态性位点
[39]
,
PCR
产
物直接测序的方法限制了这种种内位点多态性,因此
可以先通过克隆
PCR
产物中不同的重复单位,然后
再进行测序来确定每一种重复单位的序列及所占比
例。
2.3.
ITS
序列在裸子植物鉴定中的应用
与被子植物相比,裸子植物中的
ITS
序列长度
更长,变异幅度更大,尤其是长度变异尤为显著,
变异幅度为
975
~
3125
bp
,其中变异较大的部分是
ITS1
片段,而
5.8S
rDNA
+
ITS2
的长度变化范围约
在
375
~
450
bp
之间,因此,一般认为
ITS
不适用于
裸子植物的鉴定和系统发育研究
[
40]
。裸子植物较长
的
ITS
片段给扩增、测序等都增加了难度,但同时
变异较大也表明其较大的信息含量。近年来利用
ITS
片段对裸子植物某些类群的研究分析工作有了新的
进展
[
41]
。
通过
PCR
扩增并测序并研究裸子植物
rRN A
基因
间转录间隔区的结构,屈良鹄等
[42]
发现裸子植物的
ITS
区较其它植物长几百到上千个核苷酸,如苔藓、
蕨类和被子植物。通过对
28
种冷杉属
(
Abies
)
植物的
ITS
序列进行研究,向巧萍等
[41]
发现
ITS
长度的变化
主要是由于
ITS1
的长度变异所致,同时,冷杉属属
下阶元植物的
ITS
片段长度变异虽然很大,但较长的
ITS
的加长部分可以被看作一个插入序列,其他部分
序列则有较高同源性,并不影响采用
ITS
序列进行系
统发育分析。但较大的长度变异常给排序带来太大的
困难,削弱了
ITS
在分子鉴定和系统发育研究中的应
用效果。由于人们对
ITS
序列长度变异的机理和原因
知之甚少,对
ITS
序列同源性的推测又很困难,因此,
不能直接采用
ITS
长度变异这一性状来进行系统发育
的分析,但长度的变异可以作为一个性状与其他性状
(
如形态学性状
)
一起用于系统学研究
[43]
。
2.4.
ITS
区测序方法的争议
在被子植物系统发育分析中,是测定
ITS
单一
PCR
克隆,还是直接测定
PCR
全部产物的序列更好,
尚处于争论之中
[8]
。客观的说,两种测定方法各有
其利弊。如果
rDNA
重复序列间的重合程度很高,
那么
PCR
产物直接测序就可以反映出
ITS
区的序列,
但是在
PCR
扩增中存在一定的随机错误
[
44]
。相反,
如果
rDNA
重复序列间的纯和程度较低,各重复单
位间序列差异较大,特别是当重复序列间存在插入
/
缺失时,对
PCR
产物直接测序将导致无法读出其序
列,必须对
PCR
产物进行常规克隆后再测序,如杉
科密叶杉属植物
Athrotaxis laxifolia
和
A.
cupre
s-
soides
的
ITS1
序列,如果
PCR
产物中不同重复序列
的浓度比较平均,直接测序可直接确定重复序列间
的变异,这在其他类群中也发现类似的情况
[
11
,
45
]
,
因此通过直接测定
PCR
产物的序列可以反映
rDNA
重复序列间的纯和度。另一方面,如果
PCR
产物中
不同重复序列的浓度比较平均的话,直接测序后根
据测序
-
放射自显影带纹可直接确定重复序列间的
变异情况,这些发生变异的核苷酸会在同一位点显
示出来
[
11
,
46
]
。
3.
ITS
区测序的技术流程及系统学分析
目前对
ITS
序列分析的具体流程大体相同,主要
包括总
DNA
提取、
PCR
扩增、
(
克隆
)
、测序、排序
等步骤。
3.1.
总
DNA
的提取
采用液氮冷冻研磨,
CTAB
法提取
DNA
。
3.2.
PCR
扩增、产物纯化及测序
选择合适的引物对
ITS
区进行扩增,得到双链的
PCR
产物。然后经琼脂糖电泳检测、回收、纯化。对
PCR
产物进行纯化是直接测序的关键,可用离心透析
法、透析袋法、
Gene clean
方法等,也可先对
PCR
产
物进行克隆然后测序。
3.3.
排序
用计算机软件如
CLUSTAL V
、
CLUSTAL X
、
CLUSTAL W
、
DNASTAR
程序等对所测得的序列进
行排序,然后根据空位
(
gap
)
情况对排序结果进行适当
调整。
ITS1
和
ITS2
的边界可根据已发表的植物
18S
、
5.8S
和
26S
rDNA
序列以及近缘类群的
IT S1
、
ITS2
OPEN ACCESS
36
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
序列确定,从目前发表的文章看大多数数据集中在被
子植物上。
3.4.
系统学分析
确定变异位点和信息位点的数目,然后进行系统
学分析,只有信息位点才能用于系统学分支分析。常
用的分析软件有
PAUP
分析软件、
PHYIIP3
软件包、
MEGA
等。对转换和颠换进行加权和对于将
gap
作为
缺失或新性状得到的简约树树形大致相同,但自展数
值不同。最常用的分支分析法有简约法、邻接法、最
大似然法等。
4.
ITS2
二级结构预测
ITS2
是真核生物
rDNA
基因的重要组成部分,在
rDNA
基因加工成熟过程中起到重要的作用。
ITS2
保
守的结构域和二级结构为广范围遗传距离排序提供
了重要的技术保障。
通过比较了
7
条热门条形码序列并对
6000
余个
样本的分析
[47]
,再依据各条形码评价指标,姚辉等
[48]
确定了序列
ITS2
是药用植物鉴定最优的序列,且物
种水平的鉴定成功率高达
92.7%
。此外,多位学者已
提出
ITS2
具有很多优点
[47,49]
,其序列长度较短,易于
扩增,甚至适用于已降解的药材
[50]
;作为
ITS
的一部
分,
ITS2
一直受到广大系统进化专家的青睐并被广泛
应用于系统进化的研究
[49,51]
。
ITS2
的二级结构研究也
具有一定的系统分类价值
[51,52]
。
测序峰图采用序列拼接软件
CodonCode Aligner
V 3.5
(
CodonCode.,
USA
)
校对拼接,去除低质量序列
及引物区,进行多序列比对并人工校验。对于
ITS2
序列,拼接得到一致序列后使用基于隐马尔可夫模型
的
HMMer
注释方法去除两端
5.8S
和
26S
区段获得
ITS2
间隔区序列
[53]
。去掉测序质量差、不能确定具体
物种的
(
比如含“
sp.
”的序列
)
及不完整序列。通过
HMM
真菌模型
(
Florida State University, USA
)
计算
K2P
距离。采用平均种内距离、最大种内距离及
theta
值来描述种内变异,相应的采用平均种间距离、最小
种间距离及
theta
值来描述种间变异
[54,55]
。用计算机
Perl
语言统计种间、种内遗传距离的分布频度,评价
Barcoding
Gap
[56]
。通过
Wilcoxon
两个样本的统计检
验
[57]
,进一步比较种间、种内变异程度的大小。最后
采用相似性搜索法和最小距离法考察
ITS2
的鉴定成
功率
[58]
。
姚辉等
[48]
采用植物
DNA
条形码候选序列来讨论
ITS2
片段在黄芪属药用植物中的鉴定能力。结果显示,
ITS2
基因区通用性强,序列在黄芪种属间的差异较大,
具有明显的
Barcoding Gap
,能够正确的鉴定
41
个黄
芪属植物物种,仅余
6
个物种不能鉴定,其研究表明
ITS2
能够作为黄芪属药用植物鉴定的
DNA
条形码序
列,并具有重要的应用价值。
郑言等
[
1]
对栽培稻、广陆矮四号、药用野生稻、
宽叶野生稻和高杆野生稻这几种植物的
ITS2
的二
级结构进行了预测,发现广陆矮四号稻和药用野生
稻的二级结构在一定程度上具有一定的相似性,具
有四个臂和一定的结构环,宽叶野生稻和高杆野生
稻也具有相似的特点。这些可以认为是稻属的保守
性在
ITS2
序列中的具体体现。但是也可以发现具
体结构仍然存在较大的差异。这些差异可能是由
PCR
引物的选择和种属自身的特异性这两个原因
引起的。
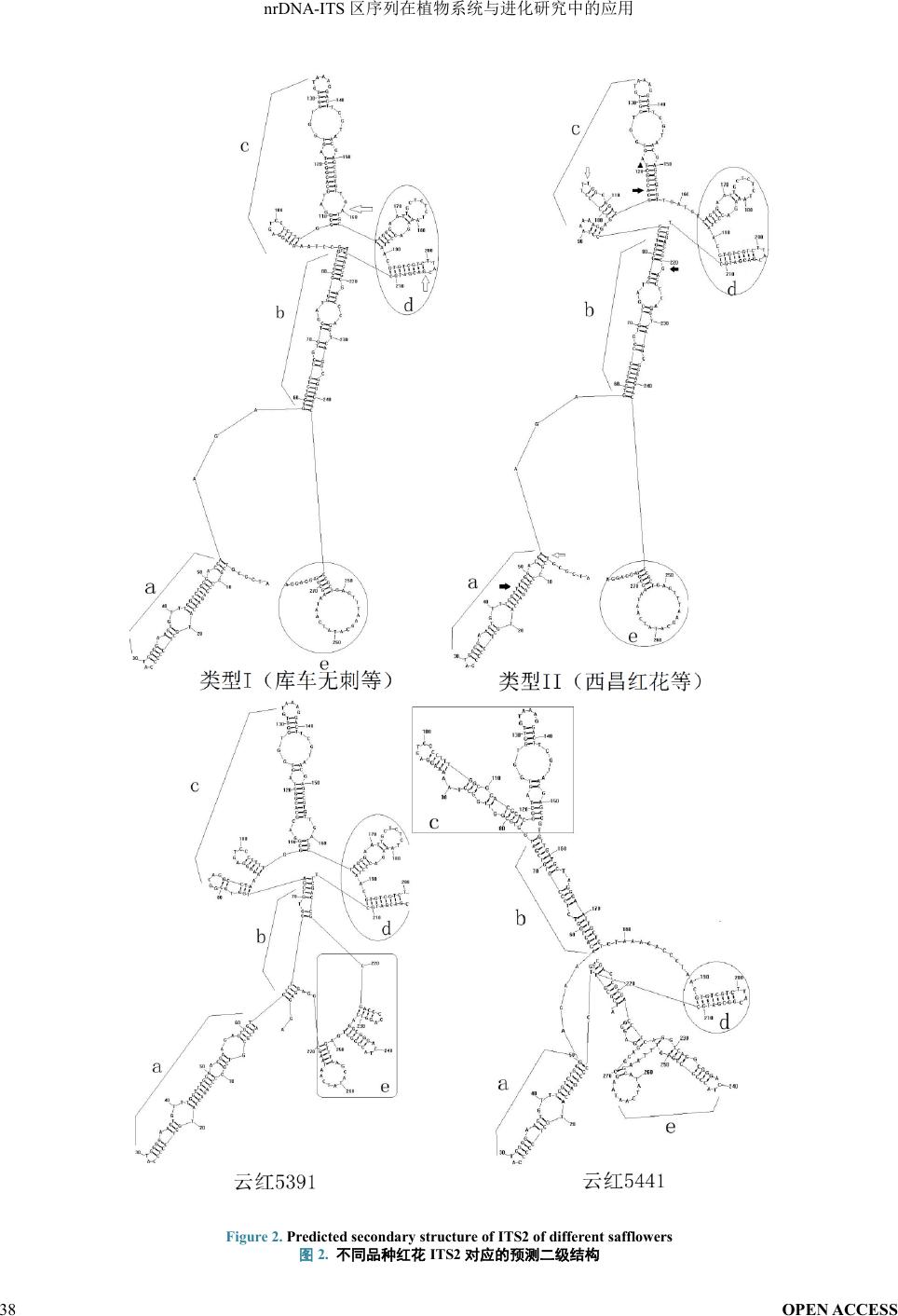
本实验室在不同品种红花
ITS
序列分析中,发现
33
个红花群体中有
15
个位点发生变异。在
ITS2
区有
三个位点发生小群体一致性突变。结果显示正常情况
下
(
如库车无刺
)
二级结构大致可分为
a
、
b
、
c
、
d
和
e
五个区
(
图
2)
。大部分红花品种可分为以库车无刺为代
表的
I
型结构和以西昌红花为代表的
II
型结构,同时
也有因为序列一级结构变异导致二级结构变化幅度
较大的云红
539
1
和云红
5441
(
图
2)
。
I
型结构:库车
无刺、南溪红花、乌鲁木齐
HH3
、吐 鲁 番
HH5
、云红
4581
和云红
5551
。其余品种除云红
5391
与云红
5441
之外,均为
II
型结构。
5.
结论
综上所述,
ITS
区序列分析已经在植物物种鉴定
以及分子系统学研究中发挥了重要的作用,为解决一
些长期存在的分类争议提供了令人信服的证据。裸子
植物和被子植物的
ITS
序列的差异很可能来自于长期
的环境剧烈的变化。相信随着分子生物学技术日新月
异的发展,人们将对
ITS
序列有更深入的了解,并探
索出更好的方法用于我国植物种质资源的鉴定和系
统学研究。
OPEN ACCESS
37
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
Fig
ure
2.
Predicted secondary
struc ture of ITS2 of different
safflower
s
图
2.
不同品种红花
ITS2
对应的预测二级结构
OPEN ACCESS
38
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
项目基金
武汉市青年晨光计划项目
(201270131395)
。
参考文献
(References)
[1]
郑言
,
覃瑞
(
2008
)
水稻与药用野生稻、宽叶野生稻、高杆野
生稻的
ITS
序列分析
.
生命科学仪器
,
6
,
30
-
40.
[2]
White
, T.J.
, Bruns
, T.D.
, Lee
, S
. and
Taylor
,
J.
(
1990)
Analysis
of phylogenetic relationships by amplification and direct se
-
quenc ing of ribosomal RN A genes
.
InnisMA.
PCR
Protoc ols: A
guide to methods a n d appl icat io ns .
Academic,
New
York
,
15
-
22.
[3]
赵玥
,
赵文军
(
2005)
核
rDNA ITS
序列在植物种质资源鉴定
中的应用
.
辽宁农业科学
,
5
,
26
-
28.
[4]
Venema,
J
. an d
Tollervey
, D. (
1999
)
Ribosome synthesis in
Sac
-
charom y ces cere vis ia e
.
Annual Review of Genetics
,
33
,
261
-
311.
[5]
Ha dji olova, A
.
Norm ann , J
.,
Cavaillé, et al.
(
1994
)
Processi n g of
truncated mouse or human rRNA transcribed from
ribosomal
minigenes transfected into mouse cells
.
Molecular and Cellular
Biology
,
14
,
4044
-
4056.
[6]
Lalev
, A.I
. and
Nazar
, R.
N.
(
1999)
Structural equivalence in the
transcribed spacer of pre
-
rRNA transcripts in
Schizosaccharo
-
myces pom
be
.
N ucleic A cids Research
,
27
,
3071
-
3078.
[7]
Joseph,
N.
,
Krauskopf,
E.
, Vera1
,
M.I.
,
et al.
(
1999
)
Ribosomal
internal transcribed spacer 2
(
ITS2) exhibits a common core of
secondary structure in vertebrates and yeast
.
Nucleic Acids R
e-
search
,
27
,
4533
-
4540.
[8]
Baldwin,
B.
G.
,
Sanderson
, M.
J.
, Porter
,
J.
M.
, et al.
(
199
5
)
The
its
region of nuclear ribosomal DNA: A valuable source of evi
-
dence on angiosperm phylogeny
.
Annals of the Missouri Botan
-
ical Garden
,
82
,
247
-
277.
[9]
Suzuki
, A.
, Ta nifuji
, S
.,
Komed a Y
.
, et al.
(
1996)
Structu ral and
functional characterization of the intergenic spacer region of the
rDNA in
Daucus carota
.
Plant and Cell Physiology
,
37
,
233
-
238.
[10]
Wang
, J.B
. and
Zhang
, W.
J.
(
2000
)
Concerted evolut ion of nuc
-
lear rDNA in allopolyploid plants.
Hereditas
,
22
, 54
-
56.
[11]
Sang,
T
.,
Crawford
, D.J.
, Stu essy
, T.F.
, et al.
(
1995)
Documenta
-
tion of reticulate evolution in peonies
(
Paeonia) using internal
transcribed spacer sequences of nuclear ribosomal DNA:
Impli
-
cations
for biogeography and concerted evolution
.
Proceedings
of the National Academy of Sciences of the United States of
Americ
a
,
92
,
6813
-
6817.
[12]
Wendel
, J.F.
, Sc hnab el
, A
. and
Seelanan
,
T.
(
1995) Bidirectional
interlocus concerted evolution following allopolyploid specia
-
tion in cotton
(
Gossypium)
.
Proceedings of the National Acad
-
emy of Scien ces of the Unit ed Sta tes of Am erica
,
92
,
280
-
284.
[13]
Elder
,
J.R
. and
Turner
, B.
J.
(
1995)
Concerted evolut ion of repe
-
ti tive DN A sequen ce in eu kar yotes
.
The Quarterly Review of Bi
-
ology
,
70
,
297
-
319.
[14]
Ainouche
, M.L
. and
Bayer
,
R.
J.
(
1997)
On the origins of the
tetraploid Bromus species
(
section Bromus, Poaceae):
Insights
from internal transcribed spacer sequences of nuclear ribosomal
DNA
.
Genome
,
40
,
730
-
743.
[15]
汪 小全
,
洪德元
(
1997
)
植物分子系统学近五 年的研究 进展
概况
.
植物分类学报
,
35
,
465
-
480.
[16]
Kollipara
, K.P.
, Singh
, R.J
. and
Hymowitz
,
T.
(
1997
)
Phyloge
-
netic and genomic relationships in the genus Glycine Willd Based
on sequences from the ITS region of nuclear rDNA
.
Genome
,
40
,
57
-
68.
[17]
Sang
, T.
, Crawford
, D.J
. and
Stuessy
, T.
F.
(
1995
) Documentation
of reticulae evolution in peonies
(
Paeonia) using internal tran
-
scribed spacer sequences of nuclear ribosomal DNA:
Implica
-
tions
forbiogeography and concerted evolution
.
Proceedings of
the National Academy of Sciences of the United States o
f America
,
92
,
6813
-
6817.
[18]
张文驹
(
1998
)
应用
rDNA
的
ITS
区探讨多倍体小麦的基因
组起源
.
武汉大学
,
武汉
.
[19]
陈之端
,
汪小全
,
孙海英等
(
1998
)
马尾树科的系统位置:来
自
rbcL
基因核苷酸序列的证据
.
植物分类学报
,
1
,
407
-
416.
[20]
王建波
,
张文驹
(
1999
)
核
rDNA
的
ITS
序列在被子植物系统
与进化研究中的应用
.
植物分类学报
,
4
,
407
-
416.
[21]
周毅
,
邹喻苹
,
洪德元等
(
1996)
中国野生稻及栽培 稻核糖
体
DNA
第一转录间隔区序列分析及其系统学意义
.
植物学报
,
38
,
785
-
791.
[22]
汪小全
,
李振宇
(
1998)
rDNA
片段的序列分析在苦苣苔亚科
系统学研究中的应用
.
植物分类学报
,
2
,
97
-
105.
[23]
章群
,
施苏华
,
黄椰林等
(
1999
)
金缕梅亚科
ITS
序列分析及
其系统发育初探
.
中山大学学报
(
自然科学版
),
1
,
107
-
110.
[24]
章群
,
施苏华
,
黄椰林等
(
2000
)
金缕梅族
ITS
序列分析及其
系统学意义
.
中山大学学报
(
自然科学版
),
1
,
72
-
76.
[25]
刘忠
,
汪小全
,
黄椰林等
(
2000
)
五味子科的系统发育:
核糖
体
DNA ITS
区序列证据
.
植物学报
,
7
,
758
-
761.
[26]
Allice
, L.A
. and
Campbe ll
, C.
S.
(
1999)
Phylogeny
of
r
ubus
(
Ro
-
saceae) based on nuclear ribosomal DNA internal transcribed
spacer region sequences
.
American Journal of Botany
,
1
,
81
-
97.
[27]
保曙琳
,
丁小余
,
常俊等
(
2004
)
长江中下游地区菱属植物的
DNA
分子鉴别
.
中草药
,
8
,
926
-
930.
[28]
Ding
, X.Y.
, Wang
, Z.T.
, Xu
, H.
, et al.
(
2000)
Database estab
-
lishment of the whole rDNA ITS region of Dendrobium species
of
“
Fengdou
”
and authentication by analysis of their sequences
.
Acta P h ar m aco l ogic a Sinic a
,
7
,
567
-
573.
[29]
Ma
,
X.J.
, Wa n g
, X.Q.
, Xiao
, P.G.
, et al.
(
2000
)
Comparison of
sequences between wild ginseng DNA and garden ginseing
DNA
.
China
Journal of Chinese
Ma ter
ial
Med
ical
,
4
,
206
-
209.
[30]
赵志礼
,
周开亚
,
董辉等
(
2001
)
国产
“
水山姜
”
的分类学研
究
——
来自核糖体
DNAITS
区序列的证据
.
云南植物研究
,
4
,
439
-
443.
[31]
Bao
, Y
. and
Ge
,
S.
(
2003
)
Identification of
o
ryza species with
the CD genome based on RFLP analysis of nuclear ribosomal
ITS sequences
.
Acta Pharm acologi ca Sini ca
,
7
,
762
-
765.
[32]
宋葆华
,
陈之端
,
汪小全等
(
2000
)
中国苋属
nrDNA
的
ITS
序列分析及其系统学意义
.
植物学报
,
11
,
1184
-
1189.
[33]
Francisco
, O.
J, Santos
, G.
A, Hines
, A.
, et al.
(
1997
)
Molecular
evidence for a Mediterranean origin of the Macaronesian en
-
demic genus Argyranthemum
(
Asteraceae)
.
American J ournal of
Botany
,
84
,
1595
-
1613.
[34]
Hsiao
, C.
, Chatterton
, N.J
. an d
Assay
, K.
H.
(
1994
)
Phylogenetic
relati onship s of 10 grass sp ecies:
An
a s ses sm en t o f ph ylo gen et ic
utility of the internal transcribed spacer region in nuclear ribo
-
somal DNA in monocots
.
Genome
,
37
,
112
-
120.
[35]
Wojciechowski
, M.F.
, Sanderson
, M.J.
, Baldwin
, B.G.
, et al.
(
1993
)
M on oph yly o f
aneuploid astragalus
(f
abaceae): Evidence
fr
om nuclear ribosomal dna internal transcribed spacer se
-
quences
.
American Journal of Botany
,
80
,
711
-
722.
[36]
Downie
, S., Ramanath
, S.
, Katz
-
Downie
, D. and Llanas
, E (1
996
)
A
molecular phylogeny of apiaceae subfamily apioideae:
Evi
-
dence from
nuclear ribosomal dna internal transcribed spacer
sequences
.
American Journal of Botany
,
83
,
234
-
251.
[37]
Buckler
, E.S
. and
Holtsford
, T.P. (
1996
)
Zea systematics:
Ribo
-
somal
ITS eviden ce
.
Molecular Biology and Evolution
,
13
,
612
-
622.
[38]
Suh
, W.C.
, Ross
, W
. and
Record,
Jr.
, M.T. (
1993)
Two open
complexes and a requirement for Mg2
+
to open the lambda PR
transcription start site.
Science
,
259
,
358
-
361.
[39]
Ainouche
, M.L
. and
Bayer
, R.
J.
(
1997)
On the origins of the
tetraploid
Bro
mus
species
(
section
Bromus, Poaceae):
Insights
from in ternal t ranscrib ed spac er sequences of n uclear ri bosoma l
.
DNA Genome
,
40
,
730
-
743.
[40]
Karvonen
,
P.
(
1995)
Genetic variation and structure of ribosom al
DNA
(
rDNA)
in
Scots pine and Norway spruce
.
Acta Univer sity
Ovlu ensis Se r ie s A Scien ti ae Rer um Nat urali um
,
10
, 1-
68.
[41]
向巧萍
,
向秋云
(
2000)
ITS
(
nrDNA)
片段在冷杉属植物中的
OPEN ACCESS
39
n
rDNA-
ITS
区序列在植物系统与进化研究中的应用
长度多态性及其在松科的系统与演化研究中的应用
.
植物学
报
,
9
,
946
-
951.
[42]
Qu, L.H.
, X
i, B.Q.
, S
hi
, S.H.
, et al.
(
1993)
A characteristic size
of the
ITS in gymnosperm rDNA
.
15
th
International Botanical
Congress
,
Tokyo
, 28 Aug ust
-
3 September 1993
, 201.
[43]
Gernandt
, D.S
. an d
Liston
, A. (
1999)
Internal transcribed spacer
region evo lution
in Lanx and P seudotsuga
(
Pinaceae)
.
American
Journal of Botany
,
86
,
711
-
723.
[44]
Gething
, M.J
. and
Sambrook
,
J.
(
1992
)
Protein folding in the
cell
.
Nature
,
355
, 33
-
45.
[45]
Gernandt
, D.S
. an d
Liston
,
A.
(
1999)
Internal transcribed spacer
region evo luti on in Larix and Pseudots uga
(
Pinaceae)
.
American
Journal of Botany
,
5
,
711
-
723.46.
[46]
张文驹
,
瞿礼嘉
(
1998
)
普通小麦基因组最可能的
4
个供体
的
ITS1
和
ITS2
序列及其亲缘关系
.
植物学报
:
英文版
,
11
,
994
-
1000
.
[47]
Chen
, S.L.
, Yao
, H
.,
Han
, J.P.
, et al.
(
2010
)
Validation of the
ITS2
region as a novel
DNA
barcode
for
identifying medicinal
pla n t species
.
PLoS O
ne
,
5
,
370
-
375.
[48]
高婷
,
姚辉
,
马新业等
(
2010)
中国黄芪属药用植物
DNA
条
形码
(
ITS2)
鉴定
.
世界科学技术
(
中医药现代化
),
2
,
222
-
227.
[49]
Coleman
, A.W. (
2003)
ITS2 is a doble
-
edged tool for eukayote
evolut ionary comparis ons
.
Trends in Geneti cs
,
19
,
370
-
375.
[50]
Chiou
, S.J.,
Yen
, J.H.,
Fang
, C.L., et al.
(
2007
)
Authentication of
medicinal herbs using PCR
-
Amplified ITS2 with specific pri
-
mers
.
Plant Medica
,
73
,
1421
-
1426.
[51]
Coleman
, A.W. (
2007
)
Pan
-
eukaryote ITS2 h omologies r ev ealed
by RNA secondary structure
.
Nu cleic A cids Research
,
35
,
3322
-
3329.
[52]
Prasad
, P.K.
, Tandon,
V
.,
Biswa l
, D.K.
, et al.
(
2009
)
Phyloge
-
netic reconstrucion using secondary structures and sequence
motif s of ITS2 rDN A of Paragoni mu s westerma ni (Kerbert,1
878)
Braun,1899
(
Dig enea : Para goni mida e) and rel ated spec ies
.
BMC
Genomics
,
10
,
S25.
[53]
Wolf
, M.
, Achtziger
,
M.
, Schultz
, J.,
et al.
(
2005) Homology
modeling revealed more than 20000 rRNA internal transcribed
spacer 2
(
ITS2) secondary structures
.
RNA
,
11
,
1616
-
1623.
[54]
Meier
, C.P
. and
Paulay
,
G.
(
2005)
DNA barcoding:
Error
rates
based on comprehensive sampling
.
PLoS
B iol
ogy
,
3
, Article
ID:
e422.
[55]
Meier
, R.
, Zhang
, G
. and
Ali
,
F.
(
2008)
Th e us e of mea n in st ead
of smallest interspecific distances exaggerates the size of the
“Barcoding Gap” and leads to misidentification
.
Systematic Bi
-
ology
,
57
,
809
-
813.
[56]
Lahaye
, R.
, Va n
der Bank
, M.
, Bogarin
, D.,
et al.
(
2008) DNA
barc oding the flora s of biod iversit y hotspot s
.
Proceedings of the
National Academy of Sciences of the United States of America
,
105
,
2923
-
2928.
[57]
Kress
, W.J
. and
Erickson
, D.
L.
(
2007
)
A tw o
-
locu s global DN A
barcode for land plants: The coding rbcL gene complements the
non
-
coding trnH
-
psbA spacer region
.
PLoS O
ne
,
2
, Article
ID:
e508.
[58]
Ros s
, H.A.
, Murgan
, S.
and
Li
, W.
L.
(
2008
)
Testing the reliab
i
lity
of genetic methods of species identification via simulation
.
Sys
-
tema tic Bio logy
,
57
,
216
-
230.
OPEN ACCESS
40