设为首页
加入收藏
期刊导航
网站地图
首页
期刊
数学与物理
地球与环境
信息通讯
经济与管理
生命科学
工程技术
医药卫生
人文社科
化学与材料
会议
合作
新闻
我们
招聘
千人智库
我要投搞
办刊
期刊菜单
●领域
●编委
●投稿须知
●最新文章
●检索
●投稿
文章导航
●Abstract
●Full-Text PDF
●Full-Text HTML
●Full-Text ePUB
●Linked References
●How to Cite this Article
Hans Journal of Computational Biology
计算生物学
, 2012, 2, 11-25
http://dx.doi.org/10.12677/hjcb.2012.22002
Published Online June 2012 (http
://www.hanspub.org/journal/hjcb)
Bioinformatics Analysis of Two-Component Singal
Transduction Systems of
Xanthomonas
Fen Hu, Xia Zou, Han Mei, Qing Tang, Jin He
*
State Key Laboratory of Agricultural Microbiology, College of Life Science and Technology,
Huazhong Agricultural University, Wuhan
Email:
*
hejin@mail.hzau.edu.cn
Received: Jun. 10th, 2012; revised: Jun. 24th, 2012; accepted: Jun. 27th, 2012
Abstract:
Two-component signal transduction systems (TCSs) represent the dominant sense-response mechanisms to
regulate a wide array of physiological pathways in prokaryo
tes. TCSs can regulate the majority of physiological proc-
esses, including bacterial growth, chemotaxis, osmoregulati
on, sporulation, biosynthesis of secondary metabolites,
virulence of pathogens, biofilm formation, etc. In this paper, we predicted all the TCS genes and comprehensively ana-
lyzed their biological functions in the whole genomes of 8
Xanthomonas
strains. We depicted a systematic classification
of these proteins, then analyzed their structures and putative biological functions by sequence alignment, multiple se-
quence alignment, phylogenetic tree analysis, Hidden Markov Model (HMM), secondary structure prediction etc, and
finally constructed the regulatory networks in which some
TCSs involved. Our research revealed the relationship be-
tween TCS genes and the pathogenicity of
Xanthomonas
, as well as the possible evolutionary relationship; furthermore,
our results could lay th e foundation for exploring n ew drug targets.
Keywords:
Xanthomonas
; Two-Component System (TCS); Regulatory Network; Pathogenicity; Bioinformatics
黄单胞菌双组分信号转导系统的生物信息学分析
胡
芬,邹
霞,梅
寒,唐
清,何
进
*
华中农业大学生命科学技术学院,农业微生物学国家重点实验室,武汉
Email:
*
hejin@mail.hzau.edu.cn
收稿日期:
2012
年
6
月
10
日;修回日期:
2012
年
6
月
24
日;录用日期:
2012
年
6
月
27
日
摘
要:
双组分信号转导系统是原核生物感知与响应刺激的重要代谢调节机制,广泛参与细菌的各种生理生化
反应。本文采用生物信息学手段,利用多序列对比、系统进化树分析、跨膜区分析、二级结构预测等,对已完
成全基因组测序的
8
株黄单胞菌中的双组分进行系统分类、结构分析和功能预测,初步构建了部分双组分信号
转导系统的调控网络关系图,揭示了黄单胞菌致病性与双组份之间的联系,并初步阐明了黄单胞菌种间的进化
关系,同时也为寻找新的药物靶标奠定了基础。
关键词:
黄单胞菌;双组分系统;调控网络;致病性;生物信息学
1.
引言
黄单胞菌属
(
Xanthomonas
)
属于变形菌门,黄单胞
菌科,革兰氏阴性菌。菌体呈短杆状,多单生,少双
生,单端极生鞭毛,专性好氧
[1]
。黄单胞菌属种类繁
多,《伯杰氏系统细菌学手册》
(
第二版
)
收录了
20
个
黄单胞菌种,
70
个分类地位已经确定的和
70
个分类
地位尚不确定的致病变种
[2]
。
黄单胞菌属大部分成员为致病菌,且致病性非常
多样,引起的植物病害遍布全世界,病害症状多为叶
*
通讯作者。
Copyright © 2012 Hanspub
11
黄单胞菌双组分信号转导系统的生物信息学分析
斑、叶枯、萎蔫、溃疡等。目前,由黄单胞菌引起的
水稻白叶枯病、禾谷黑径病、柑橘溃疡病、辣椒斑点
病、十字花科植物黑腐病和棉花角斑病等对农业生产
造成了巨大的危害。因此,深入了解其致病机理对黄
单胞菌的防治有深远的意义。
双组分信号转导系统
(two-component signal trans-
duction system
,
TCS)
是广泛存在于原核生物和真核生
物中的一种信号调节系统。在细菌中,该系统可以对
环境的变化做出相应的反应,直接或间接地接收且传
递生物信号,以调节相关基因的表达。典型的
TCS
由组氨酸蛋白激酶
(histidine protein kinase
,
HK)
和响
应调节蛋白
(response regulator protein
,
RR)
组成。
HK
的输入结构域感应外界条件刺激,使其传递结构域的
组氨酸
(His)
残基自磷酸化,随后,再将磷酸基团转移
到
RR
接受结构域的天冬氨酸
(Asp)
残基上,磷酸化的
RR
抑制或激活下游基因的转录
[3]
。原核生物中的
TCS
常以简单的“
HK-RR
”形式存在。但是,某些组氨酸
激酶除了含有输入结构域和传递结构域外,其
C
端还
融合了一个含有
Asp
残基的接受域,这种组氨酸激酶
被称为杂合型组氨酸激酶
(hybrid histidine kinase
,
HY)
,真核生物大多拥有
HY
参与的多步骤磷酸传递
的双组分信号系统
[4]
。在细菌中,
TCS
参与调节许多
的生理生化过程,包括细菌的趋化性、蛋白质合成、
营养物质同化、细胞运动、渗透压、群体感应、感受
性和致病性、生物膜和群体感应等
[5]
。
目前,全基因组序列被测通的
Xanthomonas
属菌株
共有
8
种,分别是水稻白叶枯病菌
(
Xanthomonas
oryzae
pv.
oryzae
,
Xoo
)MAFF311018
菌株
[6]
、
KACC10331
菌株
[7]
和
PXO99A
菌株
[8]
,
野油菜黄单胞菌野油菜致
病变种
(
Xanthomonas campestris
pv.
campestris
,
Xcc
)8004
菌株
[9]
、
B100
菌株
[10]
和
ATCC33913
菌株
[11]
,柑橘溃疡黄单胞菌
(
Xanthomonas axonopodis
pv.
citri
,
Xac
) 306
菌株
[11]
以及番茄疮痂病菌
(
Xanthomonas campes- tris
pv.
vesicatoria
,
Xcv
)85-10
菌株
[12]
。本文采用生物信息学手段,从这
8
株菌中
鉴定
HK
、
RR
和
HY
,对这些
TCS
组分进行系统分
类、结构分析和功能预测,并初步构建了部分
TCS
的调控网络关系图。该研究为进一步揭示
TCS
的作
用机制和开发药物靶标提供了重要的理论指导。
2.
材料与方法
2.1.
序列来源
Xoo
KACC10331(GenBank accession
NC006834.1)
、
Xoo
MAFF311018(GenBank accession
NC007705.1 )
、
Xoo
PXO99A(GenBank accession
NC010717.1 )
、
Xcc
8004(GenBank accession
NC007086.1 )
、
Xcc
ATCC33913(GenBank accession
NC003902.1 )
、
Xcc
B100(GenBank accession
NC010688.1 )
、
Xcv
85-10(GenBank accession
NC007508.1 )
和
Xac
306(GenBank accession
NC005240.1 )
全基因组序列来自
GenBank
(www.ncbi.nih.gov/genomes/Bacteria)
;其基因组注释
来自华中农业大学
DIGAP
(http://ibi.hzau.edu.cn/digap/phytopa thogens.php)
。
2.2. T CS
预测、多重序列比对及进化树构建
根据上述
8
株菌的基因组注释搜寻可能的
HK
、
RR
和
HY
,利用
Pfam
中
HK
和
RR
的保守结构域
HATPase_c(Pfam02518)
和
Response_reg(Pfam00072)
对结果进行筛选,最后通过
NCBI
的
BLASTp
程序确
认
TCS
的各组分;蛋白质序列比对采用
ClustalW
软
件进行;进化树利用同源基因非同义突变的相邻–连
接分析,
采用
ClustalW
和
Mega(Molecular Evolutionary
Genetics Analysis)Vision 4.1
软件进行构建,分析结果
进行
boot- strap
验证,重复次数设置为
1000
。
2.3.
序列相似性分析和功能域分析
利用
Pfam
和
BLASTp
进行保守结构域分析,用
ClustalW
分析序列的相似性,
TMHMM Server 2.0
(h ttp://www.cbs.dtu.dk/services/TMHMM-2.0 )
对
TCS
跨
膜结构域进行预测,结合文献报道,预测
TCS
各组分的
功能,
并在此基础上构建黄单胞菌
TCS
信号转导网络。
3.
结果
3.1.
黄单胞菌双组分系统各组分的鉴定
对黄单胞菌株全基因组序列进行生物信息学分
析,我们一共预测出
671
个编码
TCS
各组分
(
包括
HK
、
RR
和
HY)
的基因,分布特征如表
1
所示,详细信息
见附表
1~8
。可见,
TCS
基因均匀地分布于这
8
株
Xanthomonas
基因组中,
且在不同菌株中
TCS
的排列
顺序和相对位置也相似,如图
1
。
Cop
yright © 2012 Hanspub
12
黄单胞菌双组分信号转导系统的生物信息学分析
Table 1. General features of TCSs in the 8 genomes of
Xanthomonas
表
1.
黄单胞菌中双组分信号转导系统的特征
General feature
Gene family
No. of
genes
Average length
(bp)
Genome
(%)
No. of
HK/RR
HK 29 1304.4 0.76
RR 35 835.8 0.59
Hybrid 12 2536.5 0.62
Xoo
10331
Total 76 - 1.97
20
HK 21 1372.7 0.58
RR 33 888.2 0.59
Hybrid 11 2886.9 0.64
Xoo
311018
Total 66 - 1.81
21
HK 22 1221.2 0.51
RR 28 958.4 0.51
Hybrid 8 2585.4 0.34
Xoo
99A
Total 58 - 1.36
12
HK 31 1492.8 1.57
RR 41 779.1 0.63
Hybrid 17 2653.1 0.89
Xcc
33913
Total 89 - 3.09
27
HK 31 1404.1 0.85
RR 42 777 0.63
Hybrid 18 2677.7 0.94
Xcc
8004
Total 91 - 2.42
27
HK 31 1500.3 1.54
RR 36 838.8 0.59
Hybrid 18 2705.5 0.96
Xcc
B100
Total 85 - 3.09
23
HK 37 1509.8 1.08
RR 57 795.3 0.88
Hybrid 20 2894.6 1.12
Xcv
85-10
Total 114 - 3.02
31
HK 30 1361.9 0.82
RR 44 799.9 0.68
Hybrid 18 2584.2 0.89
Xac
306
Total 92 - 2.39
26
Figure 1. Distributions of TCSs in the chromosomes of the 8
Xan-
thomonas
strains
图
1.
双组分系统在八株黄单胞菌基因组中的分布
在这
8
株菌的
671
个
TCS
基因中,
每株菌
HK
的
数量在
21~37
之间、
RR
的数量在
28~57
之间、
HY
的
数量在
8~20
之间,可见各个菌株中的
TCS
组分的数
量波动范围并不大。另外一方面,成对的
HK/RR
的
数量也相差不大,平均为
23
对。虽然
TCS
基因在基
因组中所占的比例非常小,最高的仅为
3.09%
,但
TCS
在细菌的整个生理生化过程中却起着重要的作用,因
此,进一步深入了解
TCS
的调控机制是十分必要的。
3.2.
黄单胞菌属双组分系统各组分进化图
为了更好地了解
TCS
编码基因之间的相互联系,
我们分别构建了各组分
(HK
、
RR
、
HY)
在不同菌中的
进化树,如图
2~4
。根据氨基酸序列相似性程度,结
合功能预测
(
见表
2)
,我们将具有高相似性的一类蛋白
归于一个群
(cluster)
。分析结果显示黄单胞菌的
TCS
大致可以分为
16
个群,每一个群代表一对同源的
TCS
,它们分别是:
ExsG/ExsF
、
RpfC/RpfG
、
AlgZ/AlgR
、
ColS/ColR
、
PhoQ/PhoP
、
TctD/TctE
、
KdpD/KdpE
、
YgiY/YgiX
、
LytS/LytT
、
CreC/CreB
、
NtrB/NtrC
、
CvgS/CvgY
、
RegS/RegR
、
PilS/PilR
、
BaeS/BaeR
、
PhoR/PhoB
。
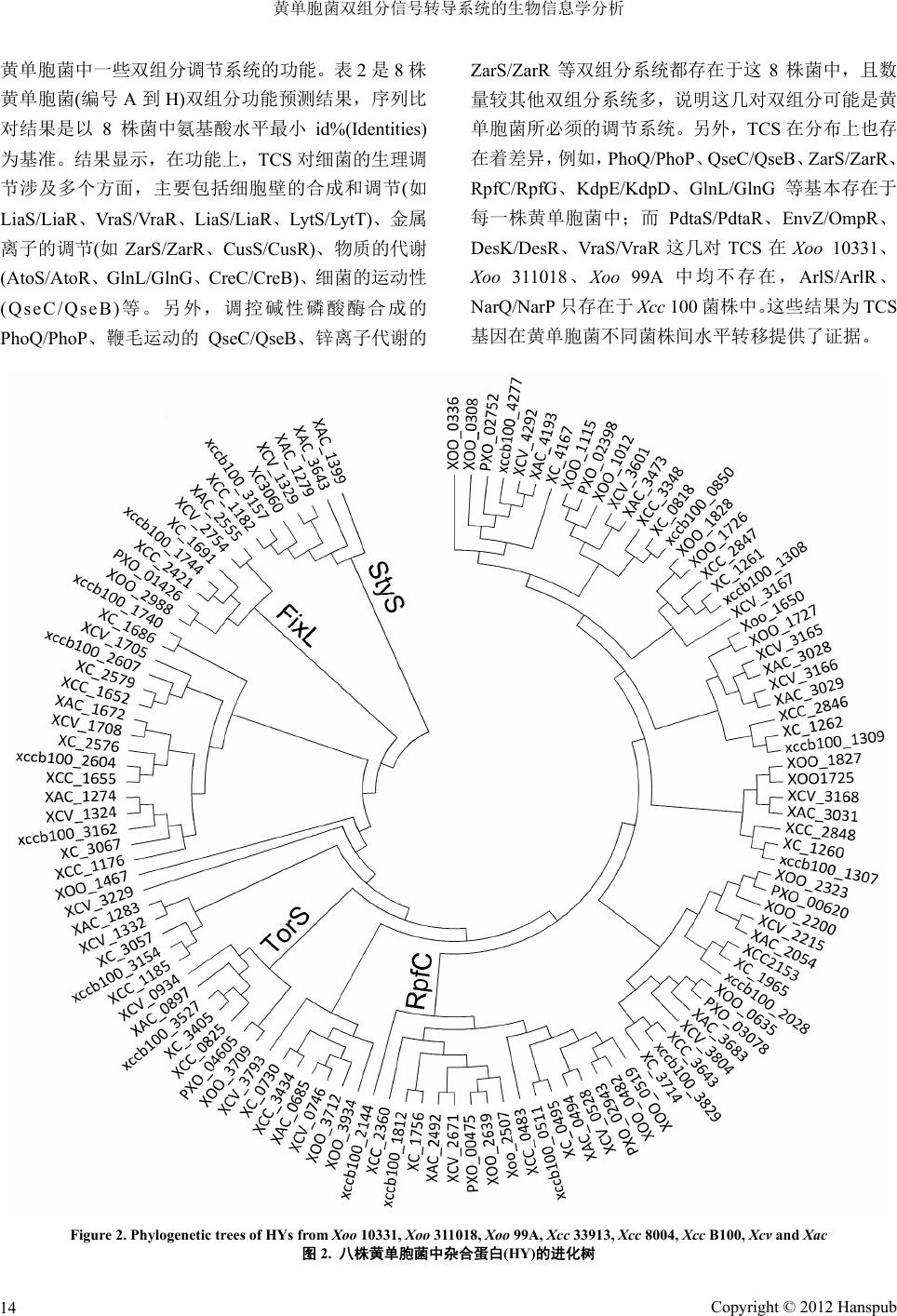
HY
进化树主要包含
4
个类群:
St yS
、
FixL
、
TorS
、
RpfC
。其中,
RpfC
主要参与对细胞毒力的和第二信
使的调控,部分是经过
HY
蛋白多步磷酸传递发挥作
用的,此调节方式为信号转导调节提供了多个调控位
点,提高了对信号感应的精确性,是细菌不断适应环
境变化的体现。
3.3.
黄单胞菌属双组分系统功能预测
根据黄单胞菌
TCS
各组分的保守结构域和在基
因组中的相对位置,我们发现了
36
对不同的
TCS
。
根据序列相似性分析和相关文献的报道,我们预测了
Cop
yright © 2012 Hanspub
13
黄单胞菌双组分信号转导系统的生物信息学分析
Copyright © 2012 Hanspub
14
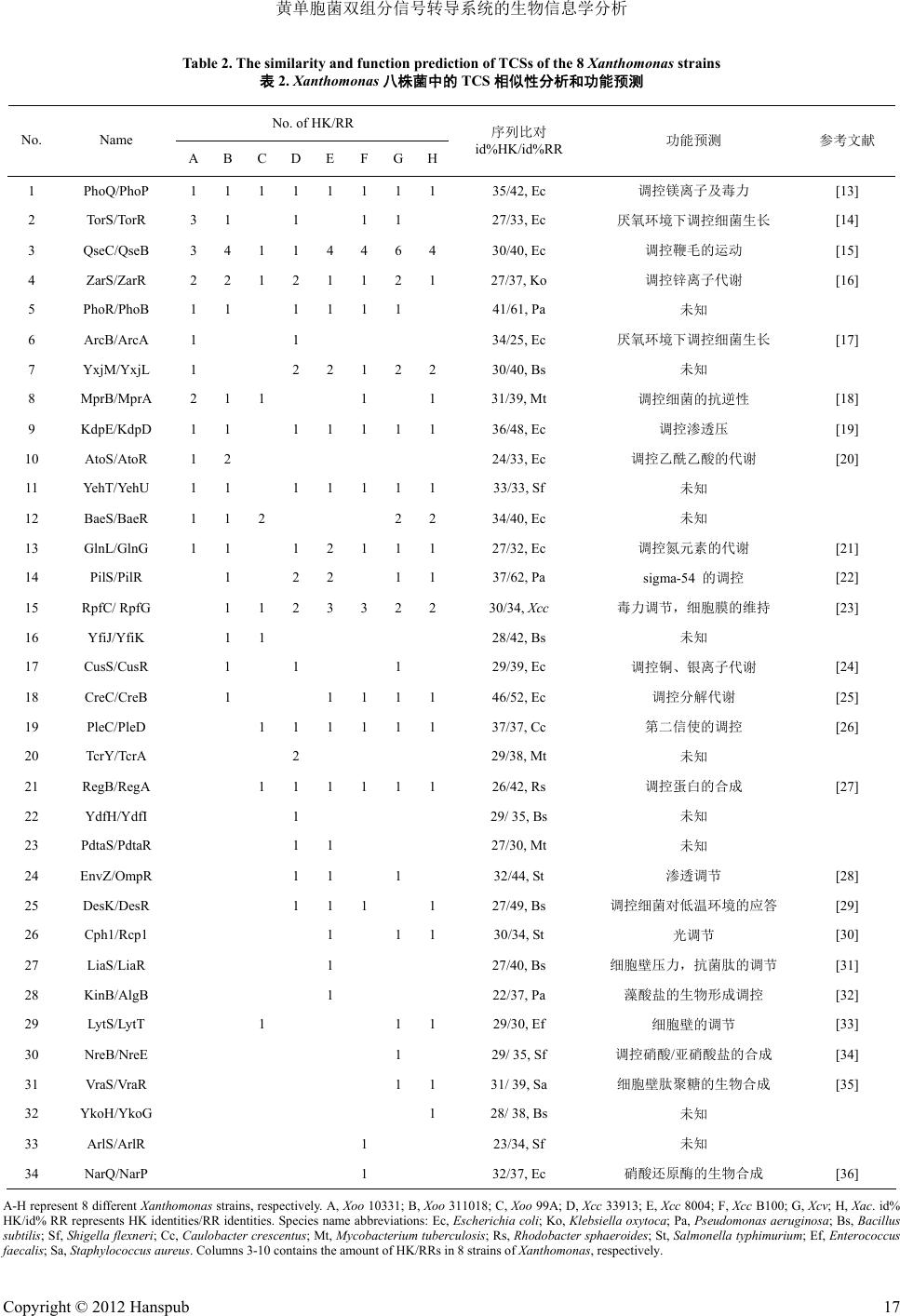
黄单胞菌中一些双组分调节系统的功能。表
2
是
8
株
ZarS/ZarR
等双组分系统都存在于这
8
株菌中,且数
量较其他双组分系统多,说明这几对双组分可能是黄
单胞菌所必须的调节系统。另外,
TCS
在分布上也存
在着差异,
例如,
PhoQ/PhoP
、
QseC/QseB
、
ZarS/ZarR
、
RpfC/RpfG
、
K dp E /KdpD
、
GlnL/GlnG
等基本存在于
每一株黄单胞菌中;而
PdtaS/PdtaR
、
EnvZ/OmpR
、
DesK/DesR
、
VraS/VraR
这几对
TCS
在
Xoo
10331
、
Xoo
311018
、
Xoo
99A
中均不存在,
ArlS/ArlR
、
NarQ/NarP
只存在于
Xcc
100
菌株中。
这些结果为
TCS
基因在黄单胞菌不同菌株间水平转移提供了证据。
黄单胞菌
(
编号
A
到
H)
双组分功能预测结果,序列比
对结果是以
8
株菌中氨基酸水平最小
id%(Identities)
为基准。结果显示,在功能上,
TCS
对细菌的生理调
节涉及多个方面,主要包括细胞壁的合成和调节
(
如
LiaS/LiaR
、
VraS/Vr aR
、
LiaS/LiaR
、
LytS/LytT)
、金属
离子的调节
(
如
ZarS/ZarR
、
CusS/CusR)
、物质的代谢
(AtoS/AtoR
、
GlnL/GlnG
、
CreC/CreB)
、细菌的运动性
(QseC/QseB)
等。另外,调控碱性磷酸酶合成的
PhoQ/PhoP
、鞭毛运动的
QseC/QseB
、锌离子代谢的
Figure 2. Phylogenetic trees of HY s from
Xoo
10331,
Xoo
311018,
Xoo
99A,
Xcc
33913,
Xcc
8004,
Xcc
B100,
Xcv
and
Xac
图
2.
八株黄单胞菌中杂合蛋白
(HY)
的进化树
黄单胞菌双组分信号转导系统的生物信息学分析
Figure 3. Phylogenetic trees of HKs from
Xoo
10331,
Xoo
311018,
Xoo
99A,
Xcc
33913,
Xcc
8004,
Xcc
B100,
Xcv
and
Xac
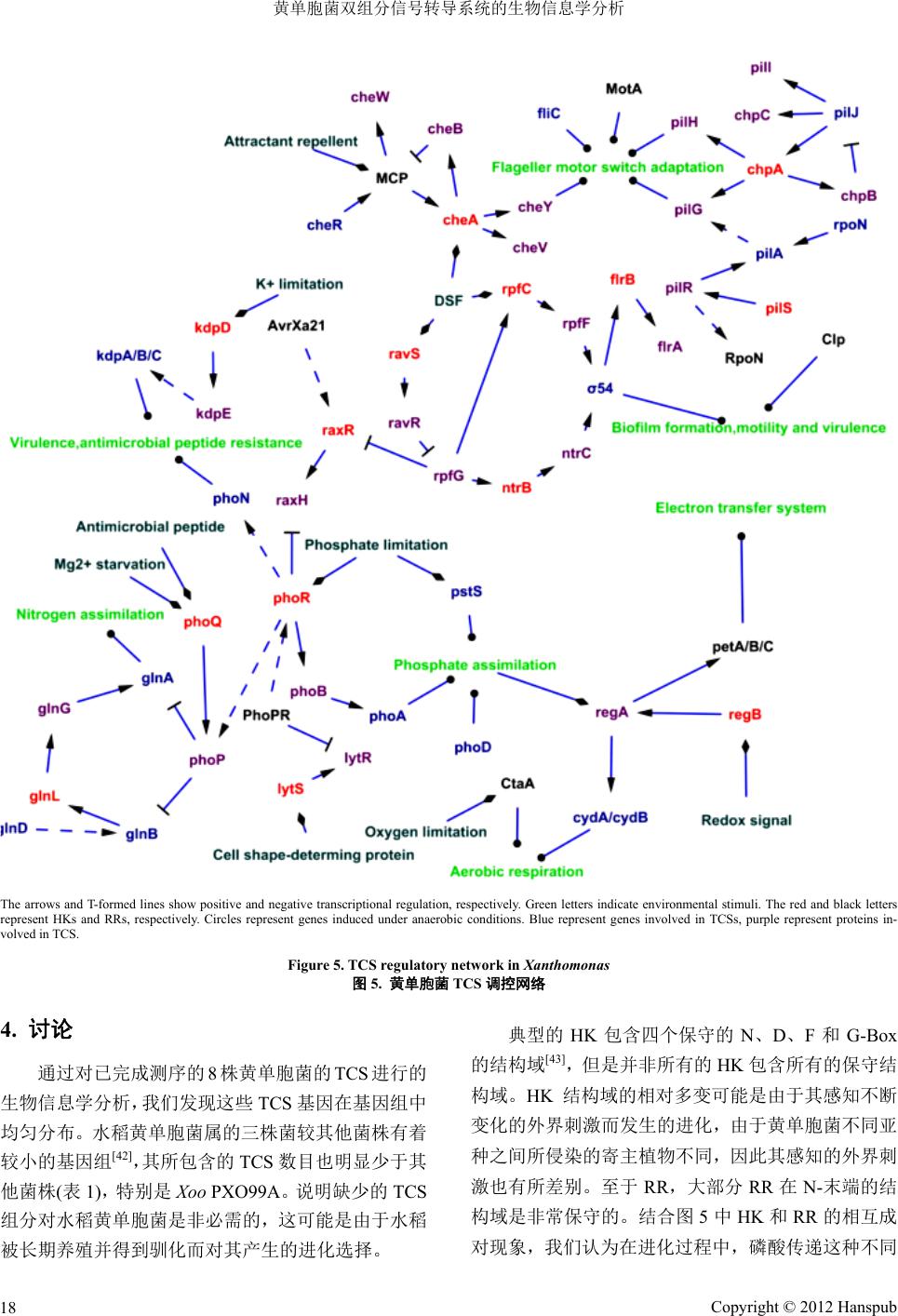
黄单胞菌属双组分系统信号调控网络
我们在预测
8
株
Xanthomonas
菌
TCS
功能的基础
上,结合相应
从网络
一方面,
一
HK
可以使多个
RR
发生磷酸化;
个
HK
磷酸 。例如,当黄
单胞菌在低
PhoP
的表达
游基因
phoPR
的表
Rax
使双组分
RpfC/Rpf F
调控下游基因的表达,而
RpfC
受另一双组分
RpfG
图
3.
八株黄单胞菌中组氨酸蛋白激酶
(HK)
的进化树
3.4.
的数据库和软件
[37,38]
,构建出黄单胞菌
中部分
TCS
的调控网络
(
图
5)
。 图中可以看到,
当
HK
和
RR
相互作用时,存在“一对多,多对一”
的信号交谈
(cross-talk)
现象,形成了更复杂的
TCS
调
控网络,从而能快速、准确地对细胞内外的各种信号
刺激做出响应。
个
同时,一个
RR
也可被多 化
Ca
2+
和
Mg
2+
离子的条件下,
PhoR
诱导
,从而调控下 达;而
PhoP
又可以被
H/RaxR
这对双组分负调控,从而
响应外界低金属离子浓度
[39]
。又如,黄单胞菌在感受
到群体感应信号
(DSF)
时,会促
Cop
yright © 2012 Hanspub
15
黄单胞菌双组分信号转导系统的生物信息学分析
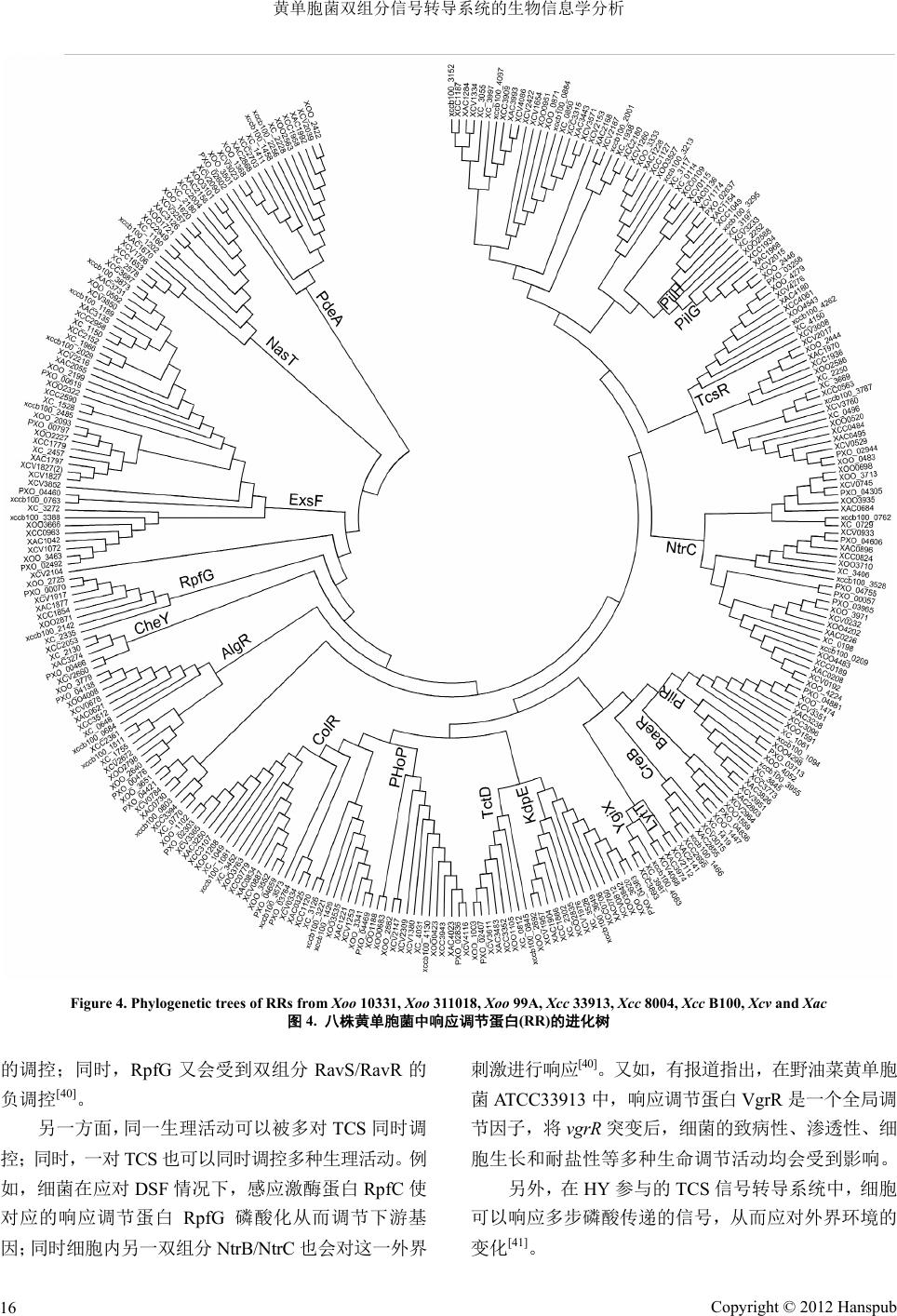
Figure 4. Phylogenetic trees of RRs from
Xoo
10331,
Xoo
311018,
Xoo
9 9A,
Xcc
33913,
Xcc
8004,
Xcc
B100,
Xcv
and
Xac
图
4.
八株黄单胞菌中响应调节蛋白
(RR)
的进化树
的调控;同时,
RpfG
又会受到双组分
RavS/RavR
的
负调控
[40]
。
另一方面,同一生理活动可以被多对
TCS
同时调
控;同时,一对
TCS
也可以同时调控多种生理活动。例
如,细菌在应对
DSF
情况下,感应激酶蛋白
RpfC
使
对应的响应调节蛋白
RpfG
磷酸化从而调节下游基
因;同时细胞内另一双组分
NtrB/NtrC
也会对这一外界
刺激进行响应
[40]
。又如,有报道指出,在野油菜黄单胞
菌
ATCC33913
中,响应调节蛋白
VgrR
是一个全 局调
节因子,将
vgrR
突变后,细菌的致病性、渗透性、细
胞生长和耐盐性等多种生命调节活动均会受到影响。
另外,在
HY
参与的
TCS
信号转导系统中,细胞
可以响应多步磷酸传递的信号,从而应对外界环境的
变化
[41]
。
Cop
yright © 2012 Hanspub
16
黄单胞菌双组分信号转导系统的生物信息学分析
Table 2. The similarity and function prediction of TCSs of the 8
Xanthomonas
strains
表
2.
Xanthomonas
八株菌中的
TCS
相似性分析和功能预测
No. of HK/RR
No. Name
A
B
C D E
FG H
序列比对
功能预测
参考文献
id%HK/id%RR
1 PhoQ/PhoP 1 11 1 1 11135/42, Ec
调控镁离子及毒力
[13]
2 TorS/TorR 3 1 1 1 1 27/33, Ec
厌氧环境下调控细菌生长
[14]
3 QseC/QseB 3 41 1 4 46430/40, Ec
调控鞭毛的运动
[15]
4 ZarS/ZarR 2 21 2 1 12127/37, Ko
调控锌离子代谢
[16]
5 PhoR/PhoB 1 1 1 1 11 41/61, Pa
未知
6 ArcB/ArcA 1 1 34/25, Ec
厌氧环境下调控细菌生长
[17]
7 YxjM/YxjL 1 2 2 12230/40, Bs
未知
8 MprB/MprA 2 1 1 1 131/39, Mt
调控细菌的抗逆性
[18]
9 KdpE/KdpD 1 1 1 1 11136/48, Ec
调控渗透压
[19]
10 AtoS/AtoR 1 2 24/33, Ec
调控乙酰乙酸的代谢
[20]
11 YehT/YehU 1 1 1 1 11133/33, Sf
未知
12 BaeS/BaeR 1 1 2 2 234/40, Ec
未知
13 GlnL/GlnG 1 1 1 2 11127/32, Ec
调控氮元素的代谢
[21]
14 PilS/PilR 1 2 2 1 137/62, Pa
sigma-54
的调控
[22]
15 RpfC/ RpfG 11 2 3 32230/34,
Xcc
毒力调节,细胞膜的维持
[23]
16 YfiJ/YfiK 11 28/42, Bs
未知
17 CusS/CusR 1 1 1 29/39, Ec
调控铜、银离子代谢
[24]
18 CreC/CreB 1 1 11146/52, Ec
调控分解代谢
[25]
19 PleC/PleD 1 1 1 11137/37, Cc
第二信使的调控
[26]
20 TcrY/TcrA 2 29/38, Mt
未知
21 RegB/RegA 1 1 1 11126/42, Rs
调控蛋白的合成
[27]
22 YdfH/YdfI 1 29/ 35, Bs
未知
23 PdtaS/PdtaR 1 1 27/30, Mt
未知
24 EnvZ/OmpR 1 1 1 32/44, St
渗透调节
[28]
25 DesK/DesR 1 1 1 127/49, Bs
调控细菌对低温环境的应答
[29]
26 Cph1/Rcp1 1 1 130/34, St
光调节
[30]
27 LiaS/LiaR 1 27/40, Bs
细胞壁压力,抗菌肽的调节
[31]
28
29 LytS/LytT 1
细胞壁的调节
[33]
S/Vra 1
32 1
32/
KinB/AlgB 1 22/37, Pa
藻酸盐的生物形成调控
[32]
1 129/30, Ef
30 NreB/NreE 1
31 VraR 1
29/ 35, Sf
调控硝酸
/
亚硝酸盐的合成
[34]
31/ 39, Sa
细胞壁肽聚糖的生物合成
[35]
YkoH/YkoG 28/ 38, Bs
未知
33 ArlS/ArlR 1 23/34, Sf
未知
34 NarQ/NarP 1 37, Ec
硝酸还原酶的生物合成
[36]
A-H represent 8 di fferent
Xantho onas
strains, respectively. A,
Xoo
10331; B,
Xo
HK/id% RR represents HK identities/RR identities. Species name abbreviations: E
subtilis
; Sf,
Shigella flexner i
; Cc,
Caulobacter crescen tus
; Mt,
Mycobacterium tu
faecalis
; Sa,
Staphylococcus aureus
. Columns 3-10 conta
ins the amount of HK/RR
m
o
31101
c,
Esc
berculos
cter sphaer oides
; St,
Salmonella typh imurium
; Ef,
Enterococcus
s in 8
onas
, respectively .
8
; C,
Xoo
99A
; D,
Xcc
33913
; E,
Xcc
8004
; F,
Xcc
B100
; G,
Xcv
;
H,
Xac
. id%
herichia coli
; Ko,
Klebsiella oxytoca
; Pa,
Pseudomonas aer uginosa
; Bs,
Bacillus
is
; Rs,
Rhodoba
strains of
Xanthom
Cop
yright © 2012 Hanspub
17
黄单胞菌双组分信号转导系统的生物信息学分析
The arrows anes shoosinneg
rial gulpe
n letters in
dletters
repret HKs apectively. Circles resennendu
under anic condite represent genes involved
in TCSs, purple represent ins in-
volvn TCS.
gur. TCS relatwo
homona
图
5.
黄单胞菌
TCS
调
4.
论
测序的
8
株黄的
TCS
进行的
生物信息学分们发现这些
S
基因在基因组中
均匀分布。 菌属的三株菌较其他菌株有着
较小的基因组
所包含的
TCS
数目也明显少于其
HK
包含四个
N
、
D
、
F
和
Box
的结构域 ,但是并非
有的保守结
构域。 结构域的相其感知不断
变化的外激而发生的进 由于黄单胞菌不同亚
种之间所侵染的寄主植物不同,因此其感知的外界刺
激也有所。至于
RR
,大部分
RR
在
N-
末端的结
d T-formed lin
nd RRs, res
w ptive ad
pre
ative tr
t ge
ansc
s i
ption
ced
reation, res
aerob
ctively
. Gree
ions. Blu
dicate envir
onmental stimuli. The red an black
protesen
ed i
Fie 5guory netrk in
Xant s
控网络
讨
通过对已完成 单胞菌
析,我
TC
水稻黄单胞
[42]
,其
他菌株
(
表
1)
,特 别 是
Xoo
PXO99A
。说明缺少的
TCS
组分对水稻黄单胞菌是非必需的,这可能是由于水稻
被长期养殖并得到驯化而对其产生的进化选择。
构域是非
常保守的。结合图
5
中
HK
和
RR
的相互成
对现象,我们认为在进化过程中,磷酸传递这种不同
典型的
保守的
G-
[43]
所有的
HK
包含所
HK
对多变可能是由于
界刺 化,
差别
Cop
yright © 2012 Hanspub
18
黄单胞菌双组分信号转导系统的生物信息学分析
基因之间的作用形式被保留下来了。
研究表明,
TCS
也涉及一些致病菌毒力因子的表
达调控
[44]
,但是大多数
TCS
的生理功能并没有被完全
揭示。作用机制相似的
TCS
广泛存在于各类微生物中
并调控多种重要生物学功能,因此被认为是潜在的药
物靶标。有研究者已经针对
TCS
找到一类与
ATP
竞
争以阻断
HK
激酶活性的抑制物
[45,46]
。
本文全局性地概括了黄单胞菌
TCS
的种类、功
能、及其在基因组的分布,并构建了其调控网络,为
深入研究
TCS
对黄单胞菌的生长、代谢以及毒力因子
的表达调控奠定了基础;同时揭示了黄单胞菌致病性
与
TCS
之间的联系,并初步阐明了黄单胞菌种间的进
化关系。为寻找药物靶标以及防治黄单胞菌引起的病
害提供了新的研究思路。
5.
致谢
本文由
973
项目
(2010CB126105)
、
国家自然科学基
金
(31070065)
和湖北省自然科学基金
(2010CDB1000 3)
资助。感谢王阶平博士对本文提出的建设性意见,硕
士研究生王燕也对本文给予帮助。
参考文献
(References)
[1]
郭亚辉
.
黄单胞菌属的分类研究进展
[J].
微生物学杂志
,
1997, 17(4): 50-51.
[2]
龙海
,
李一农
,
李芳荣
.
四种黄单胞菌的基因芯片检测方法
的建立
[J].
生物技术通报
, 2011, 1: 186-190.
[3]
A. H. West, A. M. Stock. Histidine kinases and response regula-
tor proteins in two-component signaling systems. Trends in Bio-
chemical Sciences, 2001, 26(6): 369-376.
[4]
程钢
,
喻子牛
.
阿维链霉菌双组分信号系统的生物信息学分
析
[J].
华中农业大学学报
, 2006, 25(6): 630-635.
[6]
H. Ochiai, Y. Inoue, M. Take
ya, et al
Xanthomonas oryzae
pv.
oryzae
suggests contr
numbers of effector genes and insertion sequence t
versity. Japan Agricultural Research Quarterly, 2005, 39: 275-
. J. Park, D. S. Park, et al. The geno me sequence of
C
r the
reconstruction of metabolic pathways involved in Xanthan bio-
synthesis. Journal of Biotechnology, 2008, 134(1-2): 33-45.
[11]
A. C. da Silva, J. A. Ferro, F.
C. Reinach, et al. Comparison of
the genome of two
Xanthomonas
pathogens with differing host
specificities. Nature, 2002, 417(688
7): 459-463.
[12]
F. Thieme, R. Koebnik, T. Bekel, et al. Insights into genome
plasticity and pathogenicity of the plant pathogenic bacterium
Xanthomonas campestris
pv.
vesicatoria
revealed by the com-
plete genome sequence. Journal of Bacteriology, 2005, 187(21):
7254-7266.
[13]
A. Kato, H. Tanabe and R. Utsumi. Molecular characterization
of the PhoP-PhoQ two-component system in
Escherichia coli
K-12: Identification of extracellular Mg
2+
-responsive promoters.
Journal of Bacteriology, 1999, 181(17): 5516-5520.
[14]
C. Jourlin, A. Bengrine, M. Chippaux, et al. An unorthodox
sensor protein (TorS) mediates the induction of the
tor
structural
genes in response to trimethylamine N-oxide in
Escherichia
coli
.
Molecular Microbiology, 1996, 20(6): 1297-1306.
[15]
V. Sperandio, A. G. Torres and J. B. Kaper. Quorum sensing
Escherichia
coli
regulators B and C (QseBC): A novel two-
component regulatory system involved in the regulation of fla-
gella and motility by quorum sensing in
E. coli
. Molecular Mi-
crobiology, 2002, 43(3): 809-821.
[16]
S. Leonhartsberger, A. Huber, F. Lottspeich, et al. The
hydH/G
genes from
Escherichia
coli
code for a zinc and lead responsive
two-component regulatory system.
Journal of Molecular Biology,
2001, 307(1): 93-105.
[17]
S. Iuchi, Z. Matsuda, T. Fujiwara, et al. The
arcB
gene of
Es-
cherichia
coli
encodes a sensor-regulator protein for anaerobic
repression of the
arc
modulon. Molecular Microbiology, 1990,
4(5): 715-727.
[18]
S. Gallien, E. Perrodou, C. Carapito, et al. Ort
ho-proteogenom-
ics: Multiple proteomes investigation through orthology and a
new MS-based protocol. Genome Research, 2009, 19(1): 128-
135.
[19]
M. O. Walderhaug, J. W. Polarek, P. Voelkner, et al. KdpD and
KdpE, proteins that control expression of the
kdpABC
operon,
are members of the two-component sensor-effector class of
regulators. Journal of Bacteriology, 1992, 174(7): 2152-2159.
[20]
E. S. Canellakis, A. A. Paterakis, S. C. Huang, et al. Identifica-
tion, cloning, and nucleotide sequencing of the ornithine decar-
boxylase antizyme gene of
Escherichia coli
. Proceedings of the
National Academy of Sciences USA, 1993, 90(15): 7129-7133.
[21]
S. Ueno-Nishio, S. Mango, L. J. Reitzer, et al. Identification and
regulation of the
glnL
operator-promoter of the complex
glnALG
operon of
Escheric hia
coli
. Journal of Bacteriolog y, 1984, 160(1):
379-384.
[22]
M. Hobbs, E. S. Collie, P. D. Free, et al. PilS and PilR, a
two-component transcriptional regulatory system controlling
[23]
H. Sla
ter, A. Alvarez-Morales, C. E. Barber, et al. A two-com-
GYP domain protein links cell-
thogenic
ity gene expression in
Xanthomonas
. Molecular Microbiology, 2000, 38(5): 986-1003.
[24]
R. A. Welch, V. Burland, G. Plunkett 3rd, et al. Extensive mosaic
mam
[5]
M
. T. Laub, M. Goulian. Specificity in two-components signal
transduction pathways. Annual Review of Genetics, 2007, 41:
121-145.
expression of type 4 fimbriae in
P
seudomonas
aeruginosa
. Mo-
lecular Microbiology, 1993, 7(5): 669-682.
. Geno
me sequence of
ibution
ponent sy
stem involving an HD-
nalling to pa
of large
o its race di-
cell sig
campestris
287.
[7]
B. M. Lee, Y
Xanthomonas oryzae
pathovar
oryzae
KACC10331, the bacterial
blight pathogen of rice. Nucleic Acids Research, 2005, 33(2):
577-586.
[8]
S. L. Salzberg, D. D. Sommer, M. C. Schatz, et al. Genome
sequence and rapid evolution of the rice pathogen
Xanthomonas
oryzae
pv.
Oryzae
PXO99A. BM Genomics, 2008, 9: 204.
[9]
W. Qian, Y. Jia, S. X. Ren, et al. Comparative and functional
genomic analyses of the pathogenicity of phytopathogen
Xan-
thomonas campestris
pv.
campestris
. Genome Research, 2005,
15(6): 757-767.
[10]
F. J. Vorhölter, S. Schneiker, A. Goesmann, et al. The genome of
Xanthomonas campestris
pv.
campestris
B100 and its use fo
str
ucture revealed by the complete genome sequence of uropa-
thogenic
Escherichia
coli
. Proceedings of the National Academy
of Sciences USA, 2002, 99(26): 17020-17024.
[25]
K. Yaoto, K. Hirao, T. Oshi ma, et al. Functional characteri-
zation
in vitro
of all two-component signal transduction systems
from
Escherichia
coli
. The Journal of Biological Chemistry,
2005, 280(2): 1448-1456.
[26]
S. P. Wang, P. L. Sharma, P. V. Schoenlein, et al. A histidine
protein kinase is involved in
polar organelle development in
Caulobacter
crescentus
. Proceedings of the National Academy
of Sciences USA, 1993, 90(2): 630-634.
[27]
M. K. Phillips-Jones, C. N. Hunter. Cloning and nucleotide
sequence of
re gA
, a putative response regulator gene of
Rhodo-
bacter
sphaeroides
. FEMS Microbiology Letters, 1994, 116(3):
Cop
yright © 2012 Hanspub
19
黄单胞菌双组分信号转导系统的生物信息学分析
Copyright © 2012 Hanspub
20
. Structure and ex-
[31]
iele, et al. Response of the oxy
S-containing NreB and apo-NreB
anaerobically growing
Staphylococcus
car-
nal of Bacteriology, 2010, 192(1): 86-93.
[35]
T. Baba, F. Takeuchi, M. Kuroda, et al. Genome and viru
11): 3259-3268.
integration and
interpretation of large-scale molecular datasets. Nucleic
Research, 2012, 40: D109-D114.
oin-
is required for
egulatory system RavS/RavR. Molecular Microbi-
51-161.
pv.
citri
.
ical Chemistry, 2006, 281(44): 33414-
269-275.
[28]
P
. Liljeström, I. Laamanen and E. T. Palva
pression of the
ompB
operon, the regulatory locus for the outer
membrane porin regulon in
Salmonella
typhimurium
LT-2. T he
Journal of Biological Chemistry, 1988, 201(4): 663-673.
[29]
P. S. Aguilar, A. M. Hernandez-Arriaga and L. E. Cybulski,
Molecular basis of thermosensing: A two-component signal
transduction thermometer in
Bacillus
subtilis
. The EMBO Jour-
nal, 2001, 20(7): 1681-1691.
[30]
K. C. Yeh, S. H. Wu, J. T. Murphy, et al. A cyanobacterial phy-
tochrome two-component light sensory system. Science, 1997,
277(5331): 1505-1508.
A
vrX
olog
T. Mascher, S. L. Zimmer, T. A. Smith,et al. Antibiotic-induc-
ible promoter regulated by the cell envelope stress-sensing two-
component system LiaRS of
Bacillus
subtilis
. Antimicrobial
Agents and Chemotherapy, 2004, 48(8): 2888-2896.
[32]
S. Ma, D. J. Wozniak and D. E. Ohman. Identification of the
histidine protein kinase KinB in
Pseudomonas
aeruginosa
and
its phosphorylation of the alginate regulator
algB
. The Journal of
Biological Chemistry, 1997, 272(29): 17952-17960.
[33]
I. T. Paulsen, L. Banerjei, G. S. Myers, et al. Role of mobile
DNA in the evolution of vancomycin-resistant
Enterococcus
faecalis
. Science, 2003, 299(5615): 2071-2074.
[34]
F. Reinhart, A. Huber, R. Th
sensor NreB to air
in vivo
: Fe-
gen [
in aerobically and
nosus
. Jour
lence
3342
1.
[45]
K. Stephenson, Y. Yamaguchi and J. A. Hoch. The mechanism of
action of inhibitors of bacterial two-component signal transduc-
tion systems. The Journal of Biological Che mistry, 2000 , 275(49):
determ
inants of high virulence community-acquired MRSA.
Lancet, 2002, 359(9320): 1819-1827.
[36]
R. S. Rabin, V. Stewart. Du
al response regulators (NarL and
NarP) interact with dual sensors (NarX and NarQ) to control ni-
trate- and nitrite-regulated gene expression in
Escherichia
coli
K-12. Journal of Bacteriology, 1993, 175(
[37]
M. Kanehisa, S. Goto, Y. Sato, et
al. KEGG for
Acids
and Chem
otherapy, 1999, 43(7): 1693-1699.
[38]
M
. E. Smoot, K. Ono, J. Ruscheinski, et al. Cytoscape 2.8: New
features for data integration and network visualization. Bi
formatics, 2011, 27(3): 431-432.
[39]
S.
W. Lee, K. S. Jeong, S. W. Han, et al. The
Xanthomonas
oryzae
pv.
oryzae
PhoPQ two-component system
A21 activity,
hrpG
expression, and virulence. Journal of
Bacteriology, 2008, 190(6): 2183-2197.
[40]
Y. W. He, C. Boon, L. Zhou, et al. Co-regulation of
Xanthomo-
nas campestris
virulence by quorum sensing and a novel two-
component r
y, 2009, 71(6): 1464-1476.
[41]
W. Qian, Z. J. Han and C. Z. He. Two-component signal trans-
duction systems of
Xanthomonas
spp: A lesson from genomics.
Molecular Plant-Microbe Interactions, 2008, 21(2): 1
[42]
M. O. Andrade, M. C. Alegria, C. R. Guzzo, et al. The HD-GYP
domain of RpfG mediates a direct linkage between the Rpf quo-
rum-sensing pathway and a subset of diguanylate cyclase pro-
teins in the phytopathogen
Xanthomonas axonopodis
Molecular Microbiology, 2006, 62(2): 537-551.
[43]
G. M. Pao, M. H. Saier Jr. Response regulators of bacterial sig-
nal transduction systems: Selective domain shuffling during
evolution. Journal of Molecular Evolution, 1995, 40(2): 136-
154.
44]
Y. W. He, C. Wang, L. Zhou, et al. Dual signaling functions of
the hybrid sensor kinase RpfC of
Xanthomonas campestris
in-
volve either phosphorelay or receiver domain-protein interaction.
The Journal of Biolog
38900-38904.
[46]
J. J. Hilliard, R. M. Goldschmidt, L. Licata, et al. Multiple
mechanisms of action for inhibitors of histidine protein kinases
from bacterial two-component systems. Antimicrobial Agents
黄单胞菌双组分信号转导系统的生物信息学分析
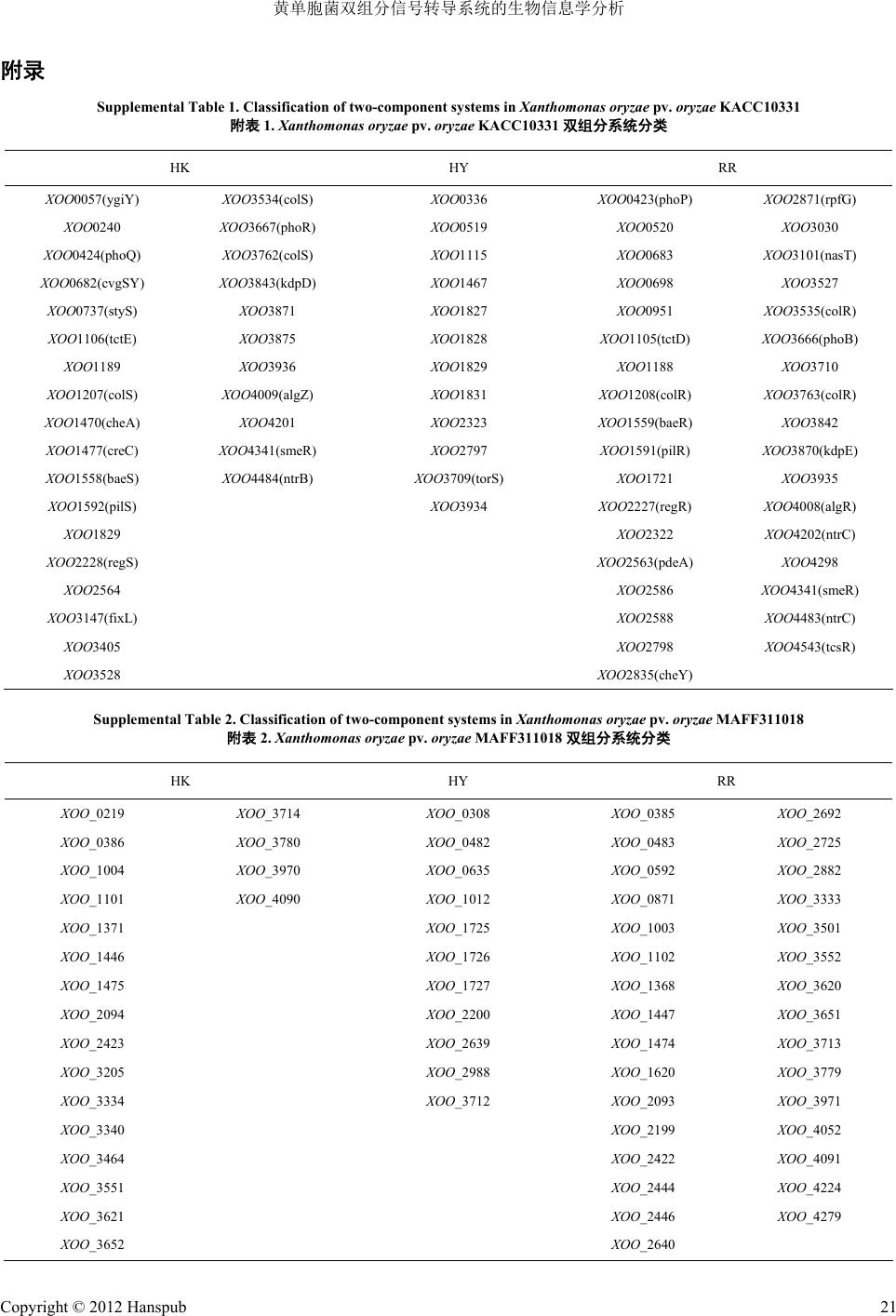
附录
Supplemental Table 1. Classification of two-componen
附表
1.
Xanthomonas
oryzae
pv.
t systems in
Xanthomonas
oryzae
pv.
oryzae
KACC10331
oryzae
KACC10331
双组分系统分类
HY RR HK
XOO
0057(ygiY)
XOO
3534(colS)
XOO
0336
XOO
0423(phoP)
XOO
2871(rpfG)
XOO
0240
XOO
3667(phoR)
XOO
XOO
0424(phoQ)
0519
XOO
0520
XOO
3030
XOO
3762(colS)
XOO
0682(cvgSY)
XOO
3843(kdpD)
XOO
XOO
0737(styS)
XOO
3871
XOO
XOO
1115
XOO
0683
XOO
3101(nasT)
1467
XOO
0698
XOO
3527
1827
XOO
0951
XOO
3535(colR)
1828
XOO
1105(tctD)
XOO
3666(phoB)
1829
XOO
1188
XOO
XOO
1106(tctE)
XOO
3875
XOO
XOO
1189
XOO
3936
XOO
3710
XOO
2586
XOO
4341(smeR)
XOO
1207(colS)
XOO
4009(algZ)
XOO
XOO
1470(cheA)
XOO
4201
XOO
1831
XOO
1208(colR)
XOO
3763(colR)
2323
XOO
1559(baeR)
XOO
3842
XOO
1477(creC)
XOO
4341(smeR)
XOO
XOO
1558(baeS)
XOO
4484(ntrB)
XOO
XOO
1592(pilS)
2797
XOO
1591(pilR)
XOO
3870(kdpE)
3709(torS)
XOO
1721
XOO
3935
XOO
3934
XOO
2227(regR)
XOO
4008(algR)
XOO
1829
XOO
2228(regS)
XOO
2322
XOO
4202(ntrC)
XOO
2563(pdeA)
XOO
4298
XOO
2564
XOO
3147(fixL)
XOO
2588
XOO
4483(ntrC)
XOO
2798
XOO
4543(tcsR)
XOO
2835(cheY)
XOO
3405
XOO
3528
Supplemental Table 2. Classification of two-component systs in
Xanthomonas
oryzae
pv.
oryzae
MAFF311018
附表
2.
Xanthomonas
oryzae
pv.
oryzae
AFF311018
双组分系统分类
HK HY RR
em
M
XOO
_0219
XOO
_3714
XOO
_0308
XOO
_0385
XOO
_2692
XOO
_038
6
XOO
_3780
XOO
_0482
XOO
_0483
XOO
_2725
XOO
_1004
XOO
_3970
XOO
_0635
XOO
_0592
XOO
_2882
XOO
_1101
XOO
_4090
XOO
_1012
XOO
_0871
XOO
_3333
XOO
_1371
XOO
_1725
XOO
_1003
XOO
_3501
XOO
_1446
XOO
_1726
XOO
_1102
XOO
_3552
XOO
_1475
XOO
_1727
XOO
_1368
XOO
_3620
XOO
_2094
XOO
_2200
XOO
_1447
XOO
_3651
XOO
_2423
XOO
_2639
XOO
_1474
XOO
_3713
XOO
_3205
XOO
_2988
XOO
_1620
XOO
_3779
XOO
_3334
XOO
_3712
XOO
_2093
XOO
_3971
XOO
_3340
XOO
_2199
XOO
_4052
XOO
_3464
XOO
_2422
XOO
_4091
XOO
_3551
XOO
_2444
XOO
_4224
XOO
_3621
XOO
_2446
XOO
_4279
XOO
_3652
XOO
_2640
Cop
yright © 2012 Hanspub
21
黄单胞菌双组分信号转导系统的生物信息学分析
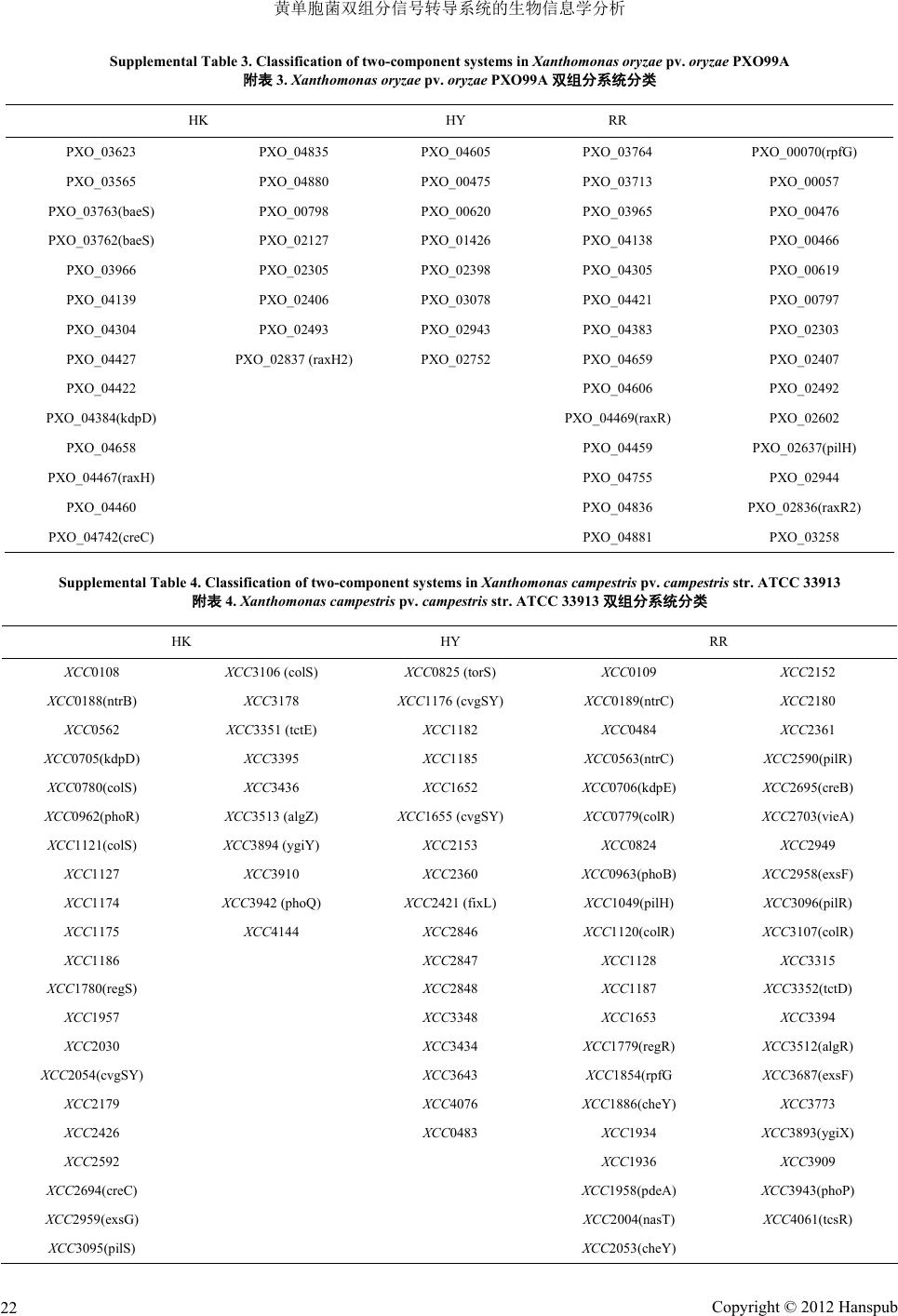
Supplemental Table 3. Classification of two-component systems in
Xanthomonas
oryzae
pv.
oryzae
PXO99A
附表
3.
Xanthomonas
oryzae
pv.
oryzae
PX O99A
双组分系统分类
HK HY RR
PXO_03623 PXO_04835 PXO_04605 PXO_03764 PXO_00070(rpfG)
PXO_03565 PXO_04880 PXO_00475 PXO_03713 PXO_00057
) PX8 PXPX
PXeS) PX7 PXPXO_04PX
5 PXPXO_04PX
406 PXPXO_04
493 PPX
7 (raxH2) PPXO_P
PX
) PXOP
PX P
) PXO_P
PXPX
PXreC) PXO_
PXO_03763(baeSO_0079O_00620 O_03965 PXO_00476
O_03762(baO_0212O_01426 138 O_00466
PXO_03966 PXO_0230O_02398 305 O_00619
PXO_04139 PXO_02O_03078 421 PXO_00797
PXO_04304 PXO_02XO_02943 O_04383 PXO_02303
PXO_0442PXO_02837 XO_02752 04659 XO_02407
PXO_04422 O_04606 PXO_02492
PXO_04384(kdpD _04469(raxR) XO_02602
PXO_04658 O_04459 XO_02637(pilH)
PXO_04467(raxH 04755 XO_02944
PXO_04460 O_04836 O_02836(raxR2)
O_04742(c 04881 PXO_03258
tal Table 4. Classificaon of two-component systemn
Xanthomonas
campes
pestris
str.
附表
4.
Xanthomonas
campestris
pv.
campestris
str. ATCC 33913
双组分系统分类
HK HY RR
Supplemen
tis i
tris
pv.
cam
ATCC 33913
XCC
0108
XCC
3106 (colS)
XCC
0825 (torS)
XCC
52
XCC
0109 21
XCC
0188(ntrB)
XCC
3178
XCC
1176 (cvgSY)
XCC
0189(ntrC)
XCC
2180
XCC
361
XCC
0705(kdpD)
XCC
3395
XCC
1185
XCC
0563(ntrC)
XCC
2590(pilR)
XCC
0780(colS)
XCC
3436
XCCXCC
0706(kdpE)
XCC
2695(creB)
XC
)
X
)
XCC
)
XCCXC
)
X
)
X
)
X
XCCXC
)
XC
)
XCXCC XC
XCCXCCXC
)
X
XC
)
X X
X
XCCXC
)
XC
)
XCCXC
)
XCC
XCC
0483
XXC
)
X
XC
)
XCCXC
)
XC
)
XCC XC
X
XCC
0562
XCC
2
XCC
3351 (tctE)
XCC
1182
XCC
0484
1652
C
0962(phoR
CC
3513 (algZ1655 (cvgSY0779(colR)
C
2703(vieA
CC
1121(colS
CC
3894 (ygiY
XCC
2153
CC
0824
XCC
2949
XCC
1127
XCC
3910
XCC
2360 0963(phoB)
C
2958(exsF
XCC
1174
C
3942 (phoQ
C
2421 (fixL) 1049(pilH)
C
3096(pilR)
XCC
1175 4144
XCC
2846 1120(colR)
C
3107(colR
XCC
1186
XCC
2847
CC
1128
XCC
3315
C
1780(regS
XCC
2848
CC
1187
CC
3352(tctD)
XCC
1957
XCC
3348
CC
1653
XCC
3394
XCC
2030
XCC
3434 1779(regR)
C
3512(algR
C
2054(cvgSY
XCC
3643 1854(rpfG
C
3687(exsF
XCC
2179
XCC
4076 1886(cheY)
XCC
3773
XCC
2426
CC
1934
C
3893(ygiX
XCC
2592
CC
1936
XCC
3909
C
2694(creC
1958(pdeA)
C
3943(phoP
C
2959(exsG
2004(nasT)
C
4061(tcsR)
CC
3095(pilS)
2053(cheY)
Cop
yright © 2012 Hanspub
22
黄单胞菌双组分信号转导系统的生物信息学分析
Sup
04
附表 类
HK HY RR
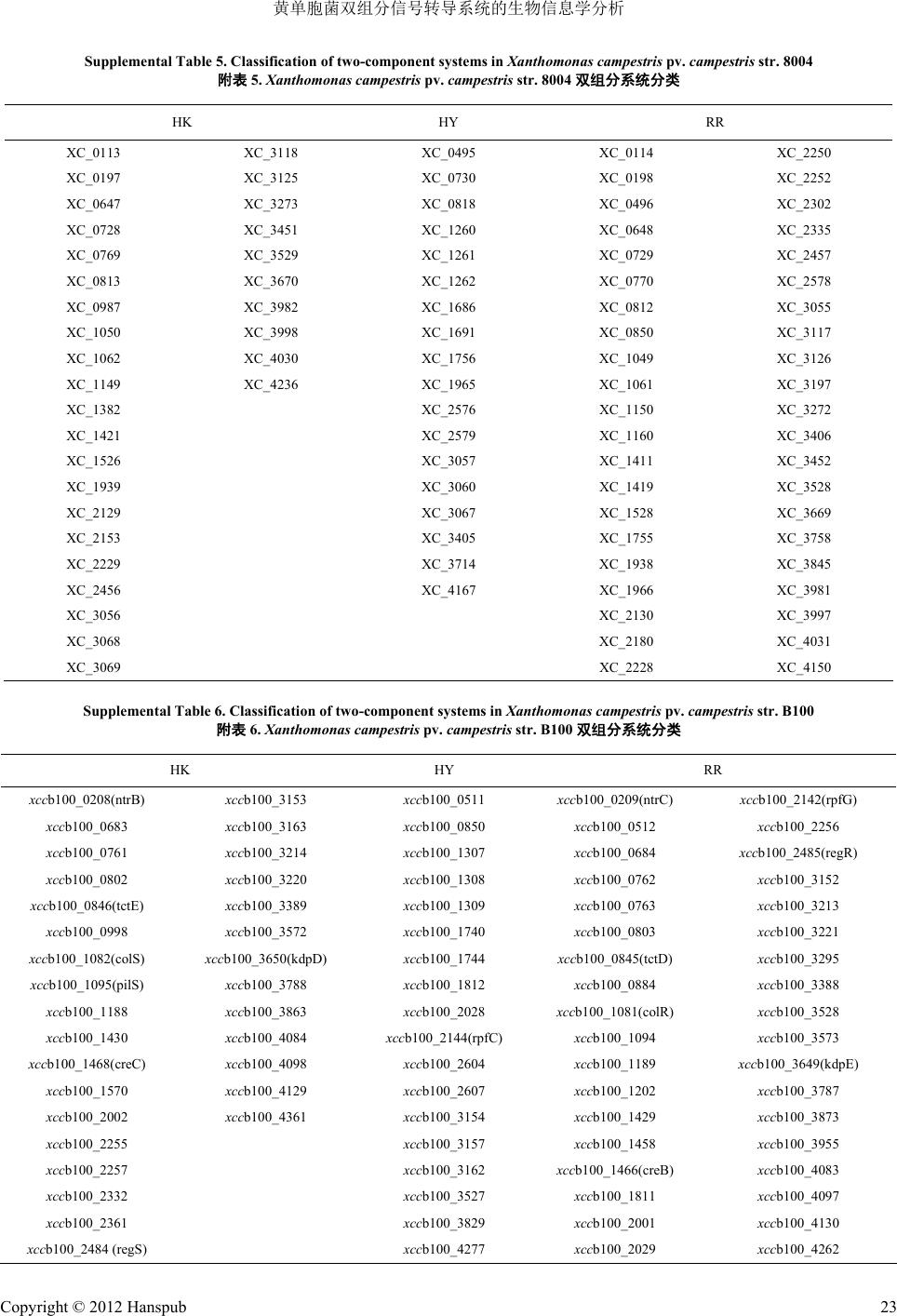
plemental Table 5. Classification of two-component systems in
Xanthomonas
campestris
pv.
campestris
str. 80
5.
Xanthomonas
campestris
pv.
campestris
str. 8004
双组分系统分
XC_0113 XC_3118 XC_0495 XC_0114 XC_2250
XC_0197 X
X
XC
XC_0769 XC_3529 XC_1261 XC_0729 XC_2457
XC
X
X
XC_423XC_1965
XC_2576
C_2579
C_3057
XC_3060
XC_3067
XC_3405
XC_2229 XC_3714 XC_1938 XC_3845
XC_3056 XC_2130 XC_3997
XC_3068 XC_2180 XC_4031
C_3125 XC_0730 XC_0198 XC_2252
XC_0647 C_3273 XC_0818 XC_0496 XC_2302
XC_0728 _3451 XC_1260 XC_0648 XC_2335
XC_0813 _3670 XC_1262 XC_0770 XC_2578
XC_0987 C_3982 XC_1686 XC_0812 XC_3055
XC_1050 C_3998 XC_1691 XC_0850 XC_3117
XC_1062 XC_4030 XC_1756 XC_1049 XC_3126
XC_1149 6 XC_1061 XC_3197
XC_1382 XC_1150 XC_3272
XC_1421 X
XC_1526 X
XC_1160
XC_1411
XC_3406
XC_3452
XC_1939 XC_1419 XC_3528
XC_2129 XC_1528 XC_3669
XC_2153 XC_1755 XC_3758
XC_2456 XC_1966 XC_3981 XC_4167
XC_3069 XC_2228 XC_4150
onent systems in Supplemental Table 6. Classif two-comp
Xanthomonas
.
campestris
str.
附
nas
campestris stris
str. B100
双组分系统分类
HK HY RR
ication of
campestris
pv
B100
表
6.
Xanthomo
pv.
campe
xcc
)
xccxcc xcc
b100_
xcc
b100_0208(ntrB
b100_3153 b100_0511 0209(ntrC) b100_2142(rpfG)
xcc
b100_0683
xcc
b100_3163
xcc
b100_0850
x
xcc
xcc
b
xcc
b10)
xcc
xcc
xcc
xcc
b1tE)
x
xcc
xcc
xcc
x
xcc
b1lS)
xcc
b100_3kdpD)
xcc
xcc
b10)
xcc
xcc xcc
b100_3788
xcc
xcc
b
xcc
xcc
b100_3863
xcc
xcc
b10)
xcc
xcc
xcc
b100_4084
xcc
b1fC)
x
xcc xcc
b100_4098
xccxcc xcc
b100_
xcc xcc
b100_4129
xccxccxcc
b100_
xcc xcc
b100_4361
xccxccxcc
b100_
xccxcc xcc
b100_
xcc
xcc
xcc
b100_3162
xcc
b10)
xcc
xcc
b100_3527
xcc
b100_3829
x
xcc
)
xcc
b100_4277
xcc
b10062
cc
b100_0512
xcc
b100_2256
xcc
b100_0761
xcc
b100_3214 b100_1307100_0684 0_2485(regR
b100_0802b100_3220b100_1308
xcc
b100_0762
xcc
b100_3152
00_0846(tc
xcc
b100_3389
xcc
b100_1309
cc
b100_0763
xcc
b100_3213
b100_0998b100_3572b100_1740
cc
b100_0803
xcc
b100_3221
00_1082(co
650(
b100_1744
0_0845(tctD
b100_3295
b100_1095(pilS) b100_1812100_0884
xcc
b100_3388
b100_1188b100_2028
0_1081(colR
b100_3528
b100_1430
00_2144(rp
cc
b100_1094
xcc
b100_3573
b100_1468(creC) b100_2604 b100_1189 3649(kdpE)
b100_1570
b100_2002
b100_2607
b100_3154
b100_1202
b100_1429
3787
3873
b100_2255
b100_3157 1458 b100_3955
b100_2257
0_1466(creB
b100_4083
xcc
b100_2332
xcc
b100_1811
xcc
b100_4097
xcc
b100_2361
cc
b100_2001
xcc
b100_4130
b100_2484 (re gS
xcc
b100_2029 _42
Cop
yright © 2012 Hanspub
23
黄单胞菌双组分信号转导系统的生物信息学分析
Supplemental Table 7. Classification of two-component systems in
Xanthomonas
campestris
pv. vesicatoria str. 85-10
附表
7.
Xanthomonas
campestris
pv. vesicatoria str. 85-10
双组分系统分类
HK HY RR
XCV
0114
XCXC
)
V
3364(colS)
XCV
0528
XCV
0115
V
2112(lytT
X
)
XC
)
XC
)
X
)
XC
)
XC
)
XC
)
XC
Q)
XCV
Y)
XC
E)
XC
)
XC
)
XC
B)
XC
)
XC
)
XC
)
XC
)
XC
)
XC
)
XCV
804
XC
)
XC
)
XCV
292
XC
)
XC
)
XCV
XCV
2155
XCV
3852
XCV
2188
XCV
1976(cheY)
XCV
3951
XCV
2310(qseC)
XCV
2015
XCV
4068
)
CV
019(ntrB
XCV
3453
XCV
0746
V
0192(ntrC
V
2147(virG
XCV
0231
CV
3610(tctE
XCV
0934
XCV
0232
XCV
2153
XCV
0335
XCV
3761
XCV
1324
XCV
0334
XCV
2187
XCV
0677
XCV
3840
XCV
1329
XCV
0529
XCV
2216
XCV
0744
XCV
4069
XCV
1332
V
0678(algR
V
2309(qseB
XCV
0783
XCV
4087
XCV
1705
XCV
0745
XCV
2422
V
0811(kdpD
V
4115(pho1708(cvgS
XCV
0784
XCV
2660
XCV
0888
XCV
4386
XCV
2215
V
0812(kdp
XCV
2672
V
1071(phoR
XCV
2671
XCV
0887
XCV
2964
XCV
1079
XCV
2754
XCV
0933
V
3015(creB
XCV
1254
XCV
3165
V
1072(pho
XCV
3023
XCV
1260
XCV
3166
XCV
1174
V
3233(pilG
XCV
1323
XCV
3167
XCV
1253
XCV
3257
XCV
1333
XCV
3168
XCV
1261
V
3351(pilR
V
1828(regS
V
3229(pilL
XCV
1334
V
3365(colR
XCV
2038
XCV
3601
XCV
1380
XCV
3571
XCV
2040
XCV
3793
V
1827(regR
XCV
3608
XCV
2105
3
XCV
1654
V
3611(tctD
V
2111(lytS
4
XCV
1706
XCV
3760
V
2145(virA
V
1827(regR
XCV
3839
2152
XCV
1917(rpfG)
XCV
3850
XCV
1957(cheY)
XCV
2965
XCV
2017
XCV
4086
XCV
3014(creC)
XCV
2039
XCV
4116(phoP)
XCV
3350(pilS)
XCV
2090(nasT
XCV
4276
Supplemental Tabion of two-cos in
Xantho
odis
pv. citri st
axonop
tr. 306
双组分系统分类
HK HY RR
le 8. Classificatmponent system
monas
axonop
r. 306
附表
8.
Xanthomonas
odis
pv. citri s
XAC
0135
XAC
3237(pilS)
XAC
0494
XAC
0136
XAC
1992
XAC
0207(ntrB)
XAC
3249(colS)
XAC
0685(torS)
XAC
0208(ntrC)
XAC
2055
XAC
0225
XAC
3273
XAC
0897
XAC
0226(ntrC)
)
XAC
)
XAC
) R)
XAC
3482(tctE) )
XAC
3975(ygiY) )
XA
) )
XAC
4022(phoQ) E) )
)
XAC
2141(lytT)
XAC
0326(smeS
3292(cvgSY1274(cvgSY
XAC
0325(sme
XAC
2158
XAC
0620(algZ)
XAC
3335
XAC
1279
XAC
0495
XAC
2168
XAC
0644
XAC
1283
XAC
0621(algR
XAC
2493
XAC
0683
XAC
1399
XAC
0684
XAC
2803(baeR
XAC
0729
C
3994
XAC
1672(cvgSY
XAC
0730
XAC
2855(creB
XAC
0759(kdpD)
XAC
2054
XAC
0760(kdp
XAC
2868(vieA
XAC
0835(colS)
XAC
2492
XAC
0834(colR
XAC
3126
Cop
yright © 2012 Hanspub
24
黄单胞菌双组分信号转导系统的生物信息学分析
Copyright © 2012 Hanspub
25
Continued
XAC
1041(phoR)
XAC
0896
XAC
3135(exsF)
XAC
2555(fixL)
XAC
1074
XA
8
XAC
1042(phoB)
XAC
3238(pilR)
XA
)
XA
)
XA
)
X
)
XA
)
X
)
XA
)
XA
)
X
)
XA
Y)
XA
)
XA
)
X
)
C
302
C
1222(colS
XAC
3029
C
1154(pilH
XAC
3250(colR)
XAC
1228
XAC
3031
XAC
1221(colR)
XAC
3274(cheY)
XAC
1273
XAC
3473
XAC
1229
XAC
3443
XAC
1282
C
3643(styS
XAC
1284
AC
3483(tctD
C
1798(regS
XAC
3683
XAC
1670
AC
3731(exsF
C
2142(lytS
XAC
4193
XAC
1797(regR)
XAC
3826
XAC
4283
C
1877(rpfG
AC
3974(ygiX
XAC
2167
C
1904(che
XAC
3993
C
2804(baeS
XAC
1968
C
4023(phoP
XAC
2854(creC)
XAC
1970
AC
4180(tcsR