 Bioprocess 生物过程, 2012, 2, 58-64 http://dx.doi.org/10.12677/bp.2012.22010 Published Online June 2012 (http://www.hanspub.org/journal/bp) The Study of Nutrient Consumption on Catharanthus roseus Cell in the Alkaloid Biosynthesis Culture Nan Zhao, Wen Chen, Zhigang Guo Department of Chemical Engineering, Tsinghua University, Beijing Email: guozhig@mail.tsinghua.edu.cn Received: May 9th, 2012; revised: May 24th, 2012; accepted: Jun. 2nd, 2012 Abstract: Catharanthus roseus cells were cultured using the EPC-500 PE bio-fermenter, and a stirring speed of 150 r/min and gas supply of 0.5 mL/min were determined to be the best cultivating conditions. During the cultivation proc- ess, the cells’ sucrose consumption was about 50% of the initial addition (30 g/L), while nitrate consumption, ammonia nitrogen consumption and phosphorus consumption were 90%, 50% and 70% of the initial addition, respectively. The content of vinblastine peaked at the fifth day. Keywords: Catharanthus roseus; Cell Culture; Bio-Fermenter; Terpene Indole Alkanoid; Vinblastine 长春花细胞生物碱合成培养过程的营养消耗研究 赵 楠,陈 文,郭志刚 清华大学化学工程系,北京 Email: guozhig@mail.tsinghua.edu.cn 收稿日期:2012 年5月9日;修回日期:2012年5月24 日;录用日期:2012 年6月2日 摘 要:本文利用 EPC-500PE 型植物细胞反应器培养了长春花细胞,筛选到较优的参数组合为150 rpm 搅拌速 度和 0.5 mL/min 供气量。在合成培养过程中,长春花细胞的蔗糖消耗量约为初始添加浓度(30 g/L)的50%,硝 态 氮的消耗量为初始添加浓度的90%,氨态氮的消耗量为初始添加浓度的 50%,磷的消耗量为初始添加浓度的 70%。长春碱含量在第 5天达到峰值。 关键词:长春花;细胞培养;生物反应器;萜类吲哚生物碱;长春碱 1. 引言 在上世纪70 年代,Zenk M.H.等[1]就利用长春花 细胞培养法生产其生物碱。但是从长春花培养细胞中 只能分离到阿玛碱或蛇根碱。在近 40 年的研究历程 中,人们进行了大量的培养基优化实验,认为氮源和 磷源可以促进细胞生长,但抑制生物碱合成。矿质元 素对长春花培养细胞的生长和生物碱积累也有一定 影响。MacCarthy 等[2]研究了在长春花细胞培养过程 中对 Mg、K、Na、Ca、S等元素的动态吸收,发现K 和Na 的吸收与细胞的生长成正比。Knobloch和Berlin [3]研究发现 Ca和Mg 与其它微量元素不影响长春花细 胞生物碱的合成。Ten Hoopen等[4]研究发现在 LS培 养基中,只有Zn 和Mo 被完全吸收,SO4、Ca、K、 Mg、Mn、Na、Fe 等只有 50%被吸收。Knobloch 和 Berlin[3]曾将培养2周的长春花细胞悬浮培养物转入 10 倍体积的 8%蔗糖溶液中,发现 2天后阿玛碱开始 积累,到 10 天达到最大。Smith等[5]发现以乳糖替代 蔗糖可明显提高生物量和长春质碱的产量。Rho 和 Andre[6]的研究表明,低磷、硝酸根和氨的培养基有利 于生物碱积累。在培养基中糖的种类和浓度极大地影 Copyright © 2012 Hanspub 58  长春花细胞生物碱合成培养过程的营养消耗研究 响生物碱的积累以及细胞的生物量。Toivonen 等[7]和 Schlatmenn[8]发现长春花细胞最适合的培养基是低氮 (氨态氮、硝态氮),低磷的培养基。并详细研究了培 养时间、磷、氮(氨态氮、硝态氮)和葡萄糖对阿玛碱 生产的影响,发现培养时间并不是影响阿玛碱生产的 重要因子,葡萄糖(最适 500 mM)才是最重要的影响因 子。阿玛碱的合成需要无氨和无磷酸根环境,硝态氮 的最佳浓度为12 mM。而色胺的最适合成条件要求低 糖,低硝酸和高磷,高氨[9]。 尽管在长春花细胞生长过程中对于营养物质的 需求的相关研究报告很多,但是很少有关于长春花细 胞在合成长春碱过程中的营养物质消耗研究报告。特 别是自学者开发出两段培养工艺之后,把植物细胞的 生长与产物的合成分成两个不同阶段,甚至在两个不 同的培养阶段使用不同的培养基。近年我们采用阻断 阿玛碱合成途径的方法有效地促进了长春质碱的合 成[10]。在此基础上进而通过对长春碱合成途径的综合 调控,能够促使长春花细胞稳定地合成长春碱。本研 究的目的是在长春花细胞两步培养过程中,了解细胞 在长春碱合成培养阶段的营养物质消耗规律,为科学 地调整或设计在长春碱合成培养阶段培养液的碳源、 氮源和磷等大量营养物质的添加浓度。为在长春花细 胞的工业化生产过程中,减少物质排放以及节约成本 提供必要的理论参数。 2. 材料与方法 2.1. 细胞培养方法 2.1.1. 细胞扩增培养 长春花细胞系是本实验室诱导并驯化培养 2年的 C03 细胞系,实验采用固–液两步法。继代或细胞扩 增培养基为改良的Murashige & Skoog(MS)固体培养 基,添加 0.5 mg/L BA(6-苄氨基嘌呤),0.5 mg/L NAA(萘乙酸),30 g/L 蔗糖,5.5 g/L 琼脂,pH 调节为 5.6~5.8。继代培养采用50 mL 培养基/200 mL 三角瓶, 在121℃高压灭菌 18 分钟后冷却备用,每瓶接种6~7 块大小均一且处于生长旺盛期的细胞团,接种量为鲜 重1.5 ± 0.1 g/瓶。在25℃,避光条件下培养,每 3周 继代培养一次。 2.1.2 生物碱合成培养 长春碱合成培养采取细胞悬浮培养方式,合成培 养基采用1/2 改良Murashige & Skoog(MS)培养基,添 加0.5 mg/L BA,0.5 mg/L NAA,30 g/L 蔗糖,100 mg/L 色氨酸、50 mg/L丙酮酸钠、50 mg/L马钱子苷和 30 mg/L 氯化铈等代谢前体和诱导子,pH 调节为5.8 ± 0.02。并装入 EYELA 日本东京理化公司5 L 搅拌式植 物细胞反应器(型号 EPC-500PE),反应器系统包括反 应器主体(Φ = 18 cm;h = 36 cm)、搅拌装置等(筐式搅 拌桨 Φ = 9 cm,h = 5 cm),系统接入了 pH 电极、DO(溶 氧)电极和 EC(电导率)电极,可以在线监测培养基的 pH 值、DO 以及EC 值。将装有培养液的反应器放入 高压灭菌锅中,121℃灭菌 40 分钟,冷却后取出放置 在超净工作台中,在无菌条件下利用0.42 μm孔径的 无菌过滤膜再过滤加入0.6 mg/L 维生素 C、5 μg/L 乙 酰辅酶 A、20 μg/L 过氧化氢、0.5 μM/L 苯丙三氮唑 和8.4 mg/L 萘普生等代谢调控因子。然后按照 3升培 养液重量比 20%的接重量接种 600 克长春花细胞。设 定培养温度为25℃,搅拌转速 110 或150 rpm,通气 量为 0.5 L/min。在培养过程中,利用计算机联机记录 监控反应罐温度、溶氧、pH、搅拌速度等参数,每天 按时采取50 mL 培养液和细胞样品,用4000 转/分的 离心机离心 5分钟,抽出上清液用于分析营养物质含 量,做出消耗曲线。将沉淀细胞称重,冷冻干燥后称 重,备用于检测生物碱含量。生物碱合成培养实验重 复三次。 2.1.3. 反应器参数优化实验 实验分两组进行,第一组实验将供气量设定在 0.5 L/min,反应器搅拌速度设定在 110 rpm和150 rpm 两 个组合,培养液体积为3 L,细胞接种量为 20%,初 始pH 5.8。 第二组实验取优化搅拌速度150 rpm,供气量分 为0.25 L/min,0.5 L/min和1 L/min三个组合,培养 液体积、初始pH 和细胞接种量与第一组实验相同。 培养温度设定在25℃,培养周期为 5天,5天后 收获细胞,冷冻干燥后称重,备用于生物碱检测。反 应器参数优化实验重复三次。 2.2. 营养物质的分析检测方法 收集培养液,用 3,5-硝基水杨酸显色法测定培养 液中的总糖含量;采用 GLU 血糖试剂盒检测葡萄糖浓 度(北京利德曼生物技术公司产);用酚二磺酸显色法 Copyright © 2012 Hanspub 59  长春花细胞生物碱合成培养过程的营养消耗研究 检测硝态氮含量;用靛定酚蓝比色法测定培养液中的 氨态氮含量,用磷钼蓝比色法分析培养液中的磷含量。 2.3. 长春花生物碱提取 采用超声萃取与摇床萃取相结合的方式对长春 花吲哚生物碱进行提取,先将收获的细胞冷冻干燥至 恒重称重,然后充分研磨再从中精确称取 200 mg样 品置于 25 mL 磨口锥形瓶内,加入 5 mL 甲醇,摇匀, 超声提取 30 min后,移到摇床上放置抽提 12 h(100 rpm,25℃)后取出,用 4000 转/分离心机离心获得上 清液。将细胞残渣再次加入5 mL甲醇,重复上述抽 提过程。最后合并二次萃取液,在旋转蒸发仪上蒸干 获得浸膏,再用 5 mL 色谱级甲醇定容。并用 0.45 μm 微孔滤膜过滤到分析用小瓶中,待测。 2.4. 长春花生物碱检测 本实验采用 HPLC方法对长春花悬浮培养细胞中 吲哚生物碱进行定量分析。 HPLC 分析仪器:SHIMADZU-CLASS-10Avp 型 高效液相色谱,色谱柱:岛津公司Shim-pack C18反 相柱 PREP-ODS(H)KIT(4.6 × 250 mm,5 μm),流动相: A:3.1 g乙酸铵、5 mL甲酸、1 L水。B:甲醇,等 梯度洗脱:A:B = 1:1,流动相流速:0.8 mL/min,紫 外检测器检测波长:260 nm,柱箱温度:40℃。 长春质碱、文多灵、长春碱、它波宁标准品购于 上海康爱生物制品有限公司,样品纯度大于98%。 3. 结果与讨论 3.1. 反应器搅拌速度和供气量优化 3.1.1. 搅拌速度优化 搅拌速度和供气量是反应器培养过程中最重要 的两个参数,调整供气量和搅拌速度能够改善细胞的 悬浮状态,并直接影响到细胞的生长和次生代谢产物 合成。图 1表示了不同搅拌速度下长春花细胞的各种 生物碱合成量。可以看到搅拌速度的变化对吲哚类生 物碱的合成没有显著影响,但是在搅拌速度为 110 rpm 时所收获的平均细胞生物量(干重)为11 g,而搅 拌速度为150 rpm 时所收获的细胞(干重)为14.4 g。这 说明 150 rpm搅拌速度下的细胞生长状态较好。其原 因是由于在该条件下培养液的溶氧水平相对较高, 0.0 0.1 0.2 0.3 0.4 0.5 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 Catharanthine&Vindoline(mg/g dryweight) rotate speed/r.p.m Catharanthine Vindoline Vinblastine Tabersonie 110 150 Vinblastine&Tabersonie(mg/g dry weight) Figure 1. Effect of agitation speed on the indole alkaloid bi os yn t he - sis of C. roseus cells 图1. 搅拌速度对长春花细胞吲哚生物碱合成的影响 细胞能够获得更多溶氧的结果。在150 rpm 条件下,长 春质碱和长春碱的含量与 110 rpm相比没有显著差异, 它波宁的含量有所下降,但是药用价值更高的文多灵 的含量却有所上升。因此在使用 EPC-500PE 型反应器 培养长春花细胞时,如果选择生物量,150 rpm搅拌速 度较有利,如果选择生物碱合成 110 rpm 较有利。由于 在两种转速条件下目标产物长春碱的含量没有什么差 异,结合生物量综合考虑,选用 150 rpm 较为有利。 3.1.2. 供气量优化 在合成培养过程中,观察发现供气量为 0.25 L/min时,细胞在培养前期出现明显的褐变现象,这 可能是由于因供氧不足而造成部分细胞死亡。收获细 胞时其平均干重只有 10 g。当供气量为 0.5 L/min 时, 细胞生长状况良好,收获时的细胞平均干重达到 14 g。 当供气量提高到1 L/min时,细胞悬浮状态良好,生 长正常,收获细胞平均干重达到 15 g。图 2表示了反 应器转速为 150 rpm,供气量分别为0.25 L/min、0.5 L/min、1 L/min时各种生物碱的含量变化。从吲哚类 生物碱的合成情况来看,供气量为 0.25 L/min 时各种 吲哚生物碱的合成均受到了抑制。当供气量提高到 0.5 L/min时,长春质碱、文多灵以及长春碱的合成得到 了促进。而将供气量提高到1 L/min时,它波宁的含 量大幅度提高,而长春质碱、文多灵和长春碱的含量 却有所下降。它波宁是合成文多灵或长春碱的前体, 由于高通气量影响了它波宁转化成文多灵或长春碱, 所以它波宁被较多积累。实验结果表明,EPC-500PE 型植物细胞反应器的转速在 150 rpm与 供气量为 Copyright © 2012 Hanspub 60 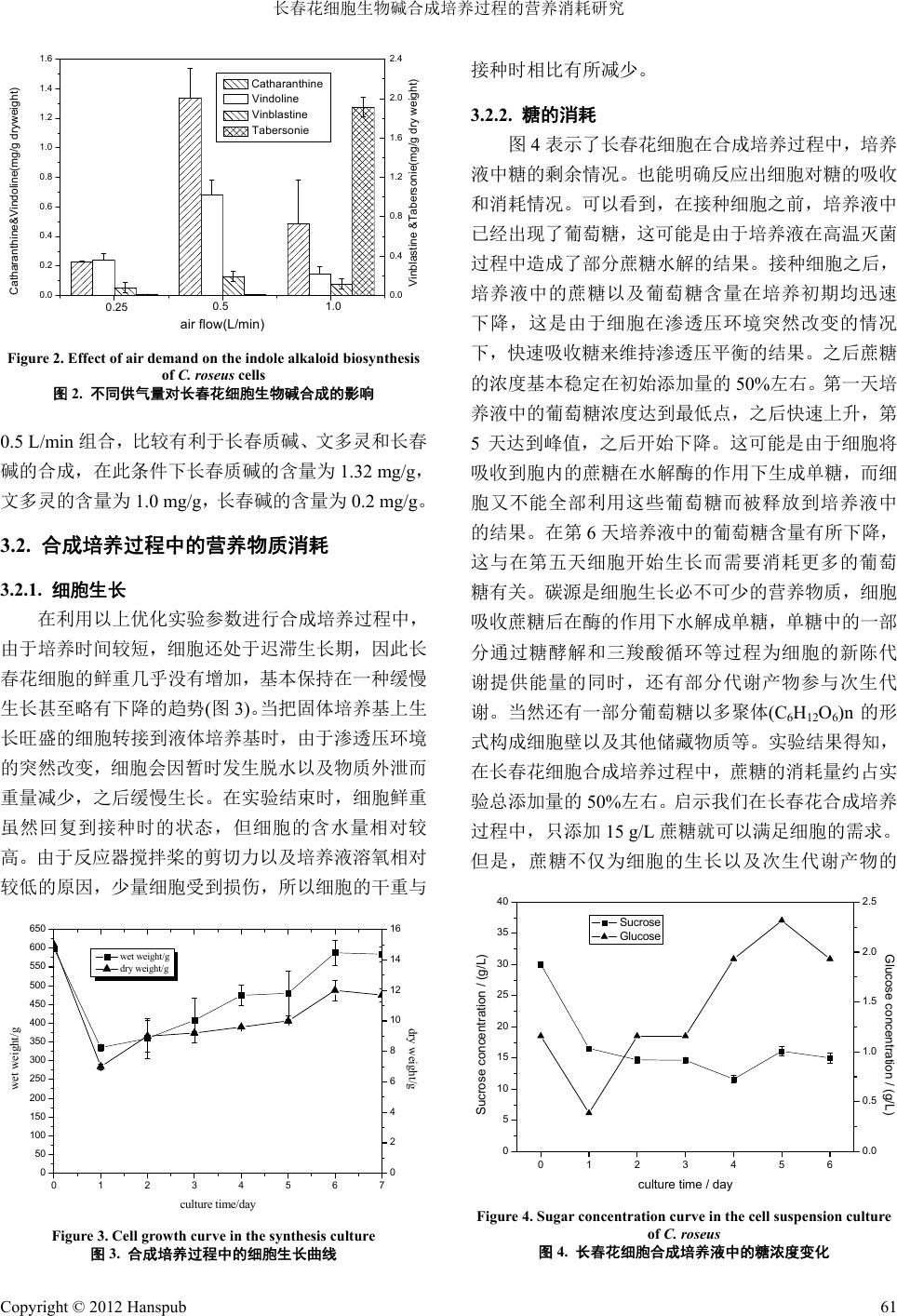 长春花细胞生物碱合成培养过程的营养消耗研究 0.0 0.4 0.8 1.2 1.6 2.0 2.4 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 Catharanthine&Vindoline(mg/g dryweigh t) air flow(L/min) 0.25 0.5 1.0 Vinblastine &Tabersonie(mg/g dry weight) Catharanthine Vindoline Vinblastine Tabersonie Figure 2. Effect of air demand on the indole alkaloid biosynthesis of C. roseus cells 图2. 不同供气量对长春花细胞生物碱合成的影响 0.5 L/min 组合,比较有利于长春质碱、文多灵和长春 碱的合成,在此条件下长春质碱的含量为1.32 mg/g, 文多灵的含量为1.0 mg/g,长春碱的含量为0.2 mg/g。 3.2. 合成培养过程中的营养物质消耗 3.2.1. 细胞生长 在利用以上优化实验参数进行合成培养过程中, 由于培养时间较短,细胞还处于迟滞生长期,因此长 春花细胞的鲜重几乎没有增加,基本保持在一种缓慢 生长甚至略有下降的趋势(图3)。当把固体培养基上生 长旺盛的细胞转接到液体培养基时,由于渗透压环境 的突然改变,细胞会因暂时发生脱水以及物质外泄而 重量减少,之后缓慢生长。在实验结束时,细胞鲜重 虽然回复到接种时的状态,但细胞的含水量相对较 高。由于反应器搅拌桨的剪切力以及培养液溶氧相对 较低的原因,少量细胞受到损伤,所以细胞的干重与 01234567 0 50 100 150 200 250 300 350 400 450 500 550 600 650 wet weight/g dry weight/g culture time/day wet weight/g 0 2 4 6 8 10 12 14 16 dry weight/g Figure 3. Cell growth curve in the synthesis culture 图3. 合成培养过程中的细胞生长曲线 接种时相比 3.2.2. 糖的消耗 春花细胞在合成培养过程中,培养 液中糖的 612 6 有所减少。 图4表示了长 剩余情况。也能明确反应出细胞对糖的吸收 和消耗情况。可以看到,在接种细胞之前,培养液中 已经出现了葡萄糖,这可能是由于培养液在高温灭菌 过程中造成了部分蔗糖水解的结果。接种细胞之后, 培养液中的蔗糖以及葡萄糖含量在培养初期均迅速 下降,这是由于细胞在渗透压环境突然改变的情况 下,快速吸收糖来维持渗透压平衡的结果。之后蔗糖 的浓度基本稳定在初始添加量的50%左右。第一天培 养液中的葡萄糖浓度达到最低点,之后快速上升,第 5天达到峰值,之后开始下降。这可能是由于细胞将 吸收到胞内的蔗糖在水解酶的作用下生成单糖,而细 胞又不能全部利用这些葡萄糖而被释放到培养液中 的结果。在第 6天培养液中的葡萄糖含量有所下降, 这与在第五天细胞开始生长而需要消耗更多的葡萄 糖有关。碳源是细胞生长必不可少的营养物质,细胞 吸收蔗糖后在酶的作用下水解成单糖,单糖中的一部 分通过糖酵解和三羧酸循环等过程为细胞的新陈代 谢提供能量的同时,还有部分代谢产物参与次生代 谢。当然还有一部分葡萄糖以多聚体(C HO )n 的形 式构成细胞壁以及其他储藏物质等。实验结果得知, 在长春花细胞合成培养过程中,蔗糖的消耗量约占实 验总添加量的50%左右。启示我们在长春花合成培养 过程中,只添加15 g/L 蔗糖就可以满足细胞的需求。 但是,蔗糖不仅为细胞的生长以及次生代谢产物的 0123456 0 5 10 15 20 25 30 35 40 Sucros e Glucose culture time / day Sucrose concentration / (g/L) 0.0 0.5 1.0 1.5 2.0 2.5 Glucose concentration / (g/L) Figure 4. Sugar concentration curve in the cell suspension culture of C. roseus 图4. 长春花细胞合成培养液中的糖浓度变化 Copyright © 2012 Hanspub 61 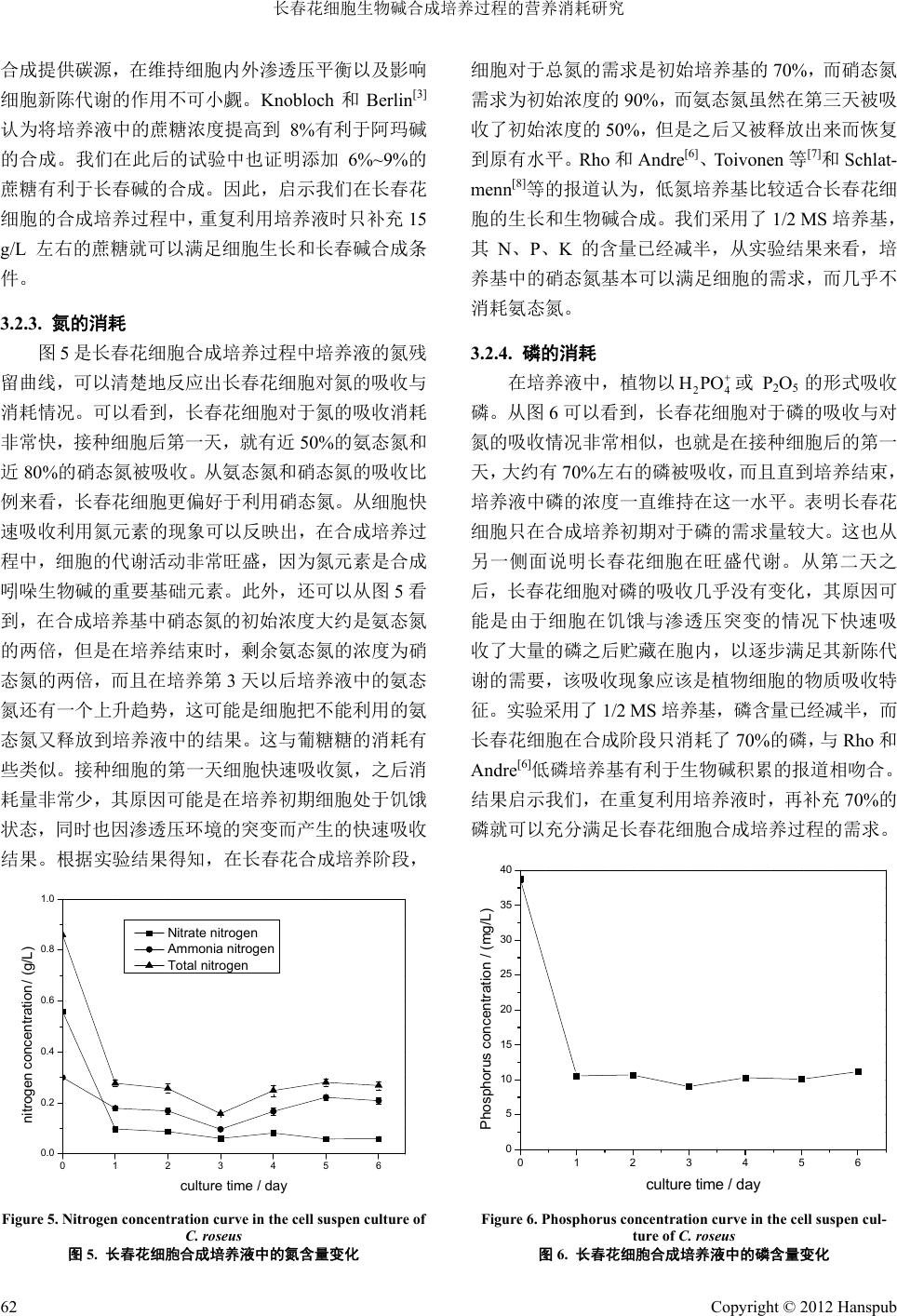 长春花细胞生物碱合成培养过程的营养消耗研究 合成提供 及影响 3.2.3. 氮的消耗 细胞合成培养过程中培养液的氮残 留曲线 碳源,在维持细胞内外渗透压平衡以 细胞新陈代谢的作用不可小觑。Knobloch 和Berlin[3] 认为将培养液中的蔗糖浓度提高到 8%有利于阿玛碱 的合成。我们在此后的试验中也证明添加6%~9%的 蔗糖有利于长春碱的合成。因此,启示我们在长春花 细胞的合成培养过程中,重复利用培养液时只补充15 g/L 左右的蔗糖就可以满足细胞生长和长春碱合成条 件。 图5是长春花 ,可以清楚地反应出长春花细胞对氮的吸收与 消耗情况。可以看到,长春花细胞对于氮的吸收消耗 非常快,接种细胞后第一天,就有近 50%的氨态氮和 近80%的硝态氮被吸收。从氨态氮和硝态氮的吸收比 例来看,长春花细胞更偏好于利用硝态氮。从细胞快 速吸收利用氮元素的现象可以反映出,在合成培养过 程中,细胞的代谢活动非常旺盛,因为氮元素是合成 吲哚生物碱的重要基础元素。此外,还可以从图 5看 到,在合成培养基中硝态氮的初始浓度大约是氨态氮 的两倍,但是在培养结束时,剩余氨态氮的浓度为硝 态氮的两倍,而且在培养第 3天以后培养液中的氨态 氮还有一个上升趋势,这可能是细胞把不能利用的氨 态氮又释放到培养液中的结果。这与葡糖糖的消耗有 些类似。接种细胞的第一天细胞快速吸收氮,之后消 耗量非常少,其原因可能是在培养初期细胞处于饥饿 状态,同时也因渗透压环境的突变而产生的快速吸收 结果。根据实验结果得知,在长春花合成培养阶段, 0123456 0.0 0.2 0.4 0.6 0.8 1.0 nitrogen concertration / (g/L) culture time / day Nitrate nit rogen Ammonia nit rogen Total nitrogen concent r ation Figure 5. Nitrogen concentration curve in the cell suspen cultur 图5. 长春花细胞合成培养液中的氮含量变化 细胞对于 硝 态 氮 植物以或 P2O5的形式吸收 磷。 花细 e of C. roseus 初始培养总氮的需求是基的70%,而 需求为初始浓度的 90%,而氨态氮虽然在第三天被吸 收了初始浓度的50%,但是之后又被释放出来而恢复 到原有水平。Rho和Andre[6]、Toivonen 等[7]和Schlat- menn[8]等的报道认为,低氮培养基比较适合长春花细 胞的生长和生物碱合成。我们采用了 1/2 MS 培养基, 其N、P、K的含量已经减半,从实验结果来看,培 养基中的硝态氮基本可以满足细胞的需求,而几乎不 消耗氨态氮。 3.2.4. 磷的消耗 在培养液中, 24 HPO 从图 6可以看到,长春胞对于磷的吸收与对 氮的吸收情况非常相似,也就是在接种细胞后的第一 天,大约有 70%左右的磷被吸收,而且直到培养结束, 培养液中磷的浓度一直维持在这一水平。表明长春花 细胞只在合成培养初期对于磷的需求量较大。这也从 另一侧面说明长春花细胞在旺盛代谢。从第二天之 后,长春花细胞对磷的吸收几乎没有变化,其原因可 能是由于细胞在饥饿与渗透压突变的情况下快速吸 收了大量的磷之后贮藏在胞内,以逐步满足其新陈代 谢的需要,该吸收现象应该是植物细胞的物质吸收特 征。实验采用了 1/2 MS 培养基,磷含量已经减半,而 长春花细胞在合成阶段只消耗了70%的磷,与Rho 和 Andre[6]低磷培养基有利于生物碱积累的报道相吻合。 结果启示我们,在重复利用培养液时,再补充 70%的 磷就可以充分满足长春花细胞合成培养过程的需求。 40 0123456 0 5 10 15 20 25 30 35 Phosphorus concentration / (mg/L) culture time / day Figure 6. Phos p horus c o n c entration curve in the cell suspen cul- ture of C. roseus 图6. 长春花细胞合成培养液中的磷含量变化 Copyright © 2012 Hanspub 62  长春花细胞生物碱合成培养过程的营养消耗研究 3.3. 生物 我们对细胞中吲哚类生物碱 的含量也进行了检测。如图 7所示,长春质碱和文多 灵的 另外一个分支途径 合成 先优化了生物反应器培养长春花细胞的 搅拌速度和供气量参数。在此基础上又研究了长春花 细胞 碱含量检测 在以上实验的同时 合成趋势相近,从培养的第一天含量开始升高, 在第三天有一个下降的过程,之后再继续升高,长春 质碱在第 6天达到峰值,文多灵在第 5天达到峰值, 之后其含量开始下降。它波宁的含量在第一天就达到 了峰值,之后快速下降并一直维持在较低水平。这可 能与它波宁是文多灵的合成前体被消耗有关。长春碱 的合成量从第一天开始缓慢上升,在第五天达到峰 值。此外,从实验结果还发现,在接种细胞时,细胞 中的各种生物碱含量几乎为零。 长春质碱和文多灵是合成长春碱的前体化合物。 其中长春质碱是由异胡豆苷通过 。在细胞合成培养初期,首先是两个上游产物它 波宁和长春质碱快速增加,与这两种物质的代谢过程 相吻合。在它波宁被合成之后,文多灵才能开始合成。 并且再与长春质碱通过过氧化酶的催化而合成长春 碱。在第五天文多灵和长春碱的含量均达到最大值, 而长春质碱的最大值延迟到第六天。这是由于长春质 碱的合成量比较大,与文多灵的产量不相匹配,所以 也影响了长春碱的合成。到了第 7天,各种吲哚生物 碱的含量开始下降,这可能与长春花细胞开始进入生 长阶段有关。根据以上实验结果,考虑药用价值最高 的生物碱是长春碱,所以采用固液两步法进行合成培 养时于第 5天收获细胞最有利。 4. 结论 本文首 在合成培养阶段的营养物质消耗规律以及各种 吲哚生物碱的合成过程。实验结果表明,用 EPC-500PE型植物细胞反应器培养长春花细胞的较 优参数组合为搅拌速度150 rpm/0.5 mL/min供气量。 长春花细胞在合成培养过程中蔗糖的消耗量约为初 始添加量(30 g/L)的50%;长春花细胞偏好硝态氮,1/2 MS 培养基的硝态氮基本可以满足细胞的需求。细胞 几乎不消耗氨态氮。细胞对磷的消耗量是 1/2 MS培 养基中初始添加量的 70%。长春碱的含量在第 5天达 到峰值,是收获细胞的有利时机。 1.6 -1012345678 -0.2 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 -0.1 0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 Catharanthine & Vindoline / (m culture time / day g/g DW) Catharanthine Vindoline Vinblastine ) Tabersonine Vinblastine & Tabersonine / (mg/g DW Figure 7. Biosynthesis curve of TIAs 图7. 吲哚生物碱的合成曲线 5. 致谢 十一五”国家科技支撑计划项目–植物细胞 模培养及生物转化技术研究课题(2008BAI63B04) 的经 ) [1] M. H. Zenk, H. El-Shagi, H. Arens, J. Stockigt, E. W. Weiler and alkaloids serpentine and ajma- of Catharanthus roseus. In: W. 感谢“ 大规 费支持。 参考文献 (References B. Deus. Formation of the indole licine in cell suspension cultures Bare, E. Reinhard, M. H. Zenk, Eds., Plant Tissue Culture and Its Bio-Technological Application, Berlin: Springer-Verlag, 1977: 27-44. [2] J. J. MacCarthy, P. K. Stumpf. Fatty-acid composition and bio- synthesis in cell suspension cultures of Glycine max (L.) Merr., Catharanthus roseus G. Don and Nicotiana tabacum L. Planta, 1980, 147(5): 384-388. [3] K. H. Knobloch, J. Berlin. Influence of medium composition on the formation of secondary compounds in cell suspension cultures of Catharanthus roseus (L.) G. Don. Zeitschrift für Naturforschung, 1980, 35c: 551-556. [4] P. R. H. Moreno, J. E. Schlatmann, R. Van der Heijden, W. M. Van Gulik, H. J G. Hoopen, R. Verpoorte and J. J. Heijnen. Induction of ajmalicine formation and related enzyme activities in Catharanthus roseus cells: Effect of inoculum density. Applied Microbiology and Biotechnology, 1993, 39(1): 42-47. [5] J. I. Smith, N. J. Smart, W. G. W. Kurz and M. Misawa. Stimulation of indole alkaloid production in cell suspension cultures of Catharanthus roseus by abscisic acid. Planta Medica, 1987, 53(5): 470-474. [6] D. Rho, G. André. Growth and stoichiometry of a Catharantus ro seus cell suspension culture under nitrogen limiting conditions. Biotechnology and Bioengineering, 1991, 38(6): 579-587. [7] L. Toivonen, M. Ojala and V. Kauppinen. Studies on the optimization of growth and indole alkaloid production by hairy root cultures of Catharanthus roseus. Biotechnology and Bio- engineering, 1991, 37(7): 673-680. [8] J. E. Schlatmann, H. J. G. Hoopen and J. J. Heijnen. Optimization of the medium composition for alkaloid production of Catharanthus roseus using statistical experimental designs. Mededelingen van de Faculteit Landbouwwetenschappen Rijk- suniversiteit Gent, 1992, 57(4a): 1567-1569. Copyright © 2012 Hanspub 63  长春花细胞生物碱合成培养过程的营养消耗研究 Copyright © 2012 Hanspub 64 catharanthine bio- clooxygenase in the cell sus-[9] A. Pietrosiuk, M. Furmanowa and B. Łata. Catharanthus roseus: Micropropagation and in vitro techniques. Phytochemistry Reviews, 2007, 6(2-3): 459-473. [10] Z. G. Guo, Y. Liu and X. H. Xing. Enhanced synthesis through regulation of cy pension culture of Catharanthus roseus (L.) G. Don. Process Biochemistry, 2011, 46(3): 783-787. |