Hans Journal of Agricultural Sciences
Vol.07 No.02(2017), Article ID:20417,8
pages
10.12677/HJAS.2017.72015
Physiological and Biochemical Responses of Suaeda salsa to Pb2+, Cd2+ in Single and Combined Stress at Different Growth Stages
Xiaolei Yu, Luyao Wang, Chenyi Wang*, Xinyuan Zhao, Lihua Teng, Jia Yang, Jie Hu
College of Biological & Environmental Science s, Zhejiang Wanli University, Ningbo Zhejiang

Received: Apr. 13th, 2017; accepted: Apr. 25th, 2017; published: Apr. 30th, 2017

ABSTRACT
Due to the rapid development of human activities and industrialization, tidal wetlands are suffering from unprecedented pollution and destruction b which lead and cadmium pollution is one of the heavy metal pollution. While Suaeda salsa has strong heavy metal tolerance, in the removal of pollution at the same time also has high economic value. In order to study the growth and physiological and biochemical responses of Suaeda salsa under different Pb2+ and Cd2+ concentration, the plant was cultured by vermiculite outdoors, single and combined pollution of Pb2+ and Cd2+ was carried out. The results showed that the plant height and contents of SOD, Pro and MDA in seedling stage and flowering stage of Suaeda salsa were significantly different. When the Suaeda salsas were responded to the Pb2+, Cd2+ single and combined stress, SOD activities responded quickly at first, Pro’s osmotic adjustment would be activated after SOD, the disorderly MDA contents real- time indicated the damaged conditions of cell membrane. In conclusion, Suaeda salsa had strong self-adjustable mechanism when under the Pb2+, Cd2+ stress treatment and which have potential for restore the Pb2+, Cd2+ pollution, which provide a theoretical basis for injury mechanism in which halophytes Suaeda salsa restore high saline areas, such as coastal wetlands and resist the heavy metals contamination.
Keywords:Suaeda salsa, Lead, Cadmium, Stress, Physiology and Biochemistry
不同生长时期碱蓬在Pb2+、Cd2+单一及 复合胁迫下的生理生化响应
于晓磊,王路遥,赵欣园,王趁义*,滕丽华,杨佳,胡杰
浙江万里学院生物与环境学院,浙江 宁波

收稿日期:2017年4月13日;录用日期:2017年4月25日;发布日期:2017年4月30日

摘 要
由于人类活动及工业化的迅速发展,沿海滩涂湿地正在遭受前所未有的污染和破坏,其中铅、镉污染是主要的污染之一,而碱蓬有较强的重金属耐受性和耐盐性,在修复污染的同时也具有较高的经济价值。为研究不同生长时期碱蓬在Pb2+、Cd2+胁迫下的生理生化响应,采用蛭石室外培养,对碱蓬进行Pb2+、Cd2+单一及复合污染处理,结果表明:Pb2+、Cd2+单一及复合胁迫下,碱蓬幼苗期与花期的株高、超氧化物歧化酶(SOD)活性、脯氨酸(Pro)含量、丙二醛(MDA)含量均呈显著差异(P < 0.05)。碱蓬在抵御外界Pb2+、Cd2+胁迫时,以SOD活性的快速响应为先,Pro的渗透调节抵御为后,MDA的无序变化实时表征植物的受损状况。总之,碱蓬对Pb2+、Cd2+的胁迫有较强的自我调节能力,具有修复Pb2+、Cd2+污染的潜力,这为进一步利用盐生植物碱蓬修复滨海滩涂等高盐碱土壤,以及抵御Pb2+、Cd2+等重金属复合污染的损伤机制研究提供了理论依据。
关键词 :碱蓬,Pb2+,Cd2+,胁迫,生理生化

Copyright © 2017 by authors and Hans Publishers Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


1. 引言
工业企业的临海布局、人类活动的增加,远超海洋环境容量,造成海洋滩涂重金属含量超标,破坏海洋湿地生态系统,影响海产品品质,严重制约当地的渔业经济,也不利于滨海经济的可持续发展 [1] [2] 。Pb2+、Cd2+是动植物生长的非必需元素 [3] ,镉会导致细胞死亡,植物黄化枯萎 [4] 。在铅胁迫时,即使很低的浓度,植物组织也会受到损害,如微核,DNA破坏 [5] ,膜通透性的改变 [6] 和部分酶活性降低 [7] 等。此外,铅对镉在植物体内的积累有增强效果 [8] 。植物对铅和镉累积,也会刺激超氧自由基产生,造成植物氧化应激 [9] ,使膜脂过氧化,引起超氧化物歧化酶(Superoxide Dismutase, SOD)活性、丙二醛(Malondialdehyde, MDA)含量、脯氨酸(Proline, Pro)含量等生理生化指标发生变化 [10] [11] [12] 。
碱蓬(Suaeda salsa)属藜科,是本土一年草本真盐生植物,常见于盐碱地及沿海滩涂,在新疆、蒙、甘、陕、鲁、浙、晋、辽等省区均有分布。碱蓬3~6月出苗,7~8月为花期,9~10月为结实期,11月为种子枯熟期 [13] 。由于碱蓬耐盐性强,又为土著植物,从而受到盐碱湿地修复领域的广泛关注,对其重金属累积能力 [14] 、有机物吸收能力 [15] 以及对氮磷的吸收与再分配作用 [16] 等都有相关的研究报道,在一些沿海滩涂地区盐生植物碱蓬作为优势植物,可明显吸收盐渍土壤中的Cd、Pb等重金属 [17] ,被誉为盐碱地修复的先锋植物 [18] 。本文研究并比较了不同生长时期的碱蓬(幼苗期和花期),在Pb2+、Cd2+单一及复合胁迫下的生理生化响应,旨在为碱蓬的生长代谢过程中长期适应和抵御Pb2+、Cd2+胁迫的损伤机制,及其修复重金属污染的高盐碱滩涂应用等提供理论依据。
2. 材料与方法
2.1. 试剂与材料
盐地碱蓬(采集于杭州湾庵东湿地);塑料花盆(直径16 cm,高16 cm);蛭石(φ = 2~4 mm,填充花盆至2/3体积)、海水(取自象山港)。蛭石和海水中的Pb2+、Cd2+背景值如表1。
2.2. 试验方法
根据《土壤环境质量标准》 (GB15618-2008)、土壤背景值及工业区附近土壤中的Pb2+、Cd2+浓度,进行完全交互实验设计,共16个处理组,每个处理4盆重复,如表2所示。
实验分别于当年5月(幼苗期),7月(花期)开始,时间均为60天,Pb2+和Cd2+以Pb(NO3)2∙6H2O、Cd(NO3)2∙4H2O配成溶液形式加入,于种植前2周平衡固定、装盆,其中,不添加任何重金属的为对照组。当年5月,采集株高相近的幼苗,洗净根部泥沙,种于上述盆中,每盆2棵,将蛭石填入花盆中,占盆体积2/3,以Hoagland营养液:海水 = 1: 1 (盐度约为1.6%)定期浇灌,室外自然光照下培养,以Hoagland营养液:海水 = 1: 1 (盐度约为1.6%)定期浇灌,既能保持基质湿润,还能满足碱蓬对盐分的需求,并逐步适应滨海环境中的土壤盐度。参照文献 [19] ,在幼苗期、花期分别检测株高,以盆为单位,采集每株主干上相同高度、成熟度一致且生长状况相近的绿色叶片,测量其超氧化物歧化酶(SOD)活性(氮蓝四唑法) [20] 、丙二醛(MDA)含量(硫代巴比妥酸法) [21] 、脯氨酸(Pro)含量(酸性茚三酮比色法) [22] ,每组重复测量3次。数据采用SPSS 20组间方差分析、Graph Pad Prism 5作图,结果以均值±标准差表示。
3. 结果与分析
3.1. Pb2+、Cd2+胁迫对碱蓬株高的影响
由图1可知,幼苗期,随单一Cd2+浓度增加,促进碱蓬生长,使株高显著高于对照组(P < 0.05),浓度继续增大,抑制碱蓬生长,使株高显著小于对照组(P < 0.05),且浓度越大,抑制作用越强。单一Pb2+浓度增加对株高起促进-抑制-促进作用,加入5 mg∙kg−1的Cd2+使其逆转为抑制-促进-抑制作用,分别以20, 50 mg∙kg−1的Cd2+与Pb2+ (除200 mg∙kg−1外)复合,均抑制碱蓬生长。生长至花期,对照组株高最大

Table 1. Background values of Pb2+, Cd2+ in the vermiculite and sea water
表1. 蛭石和海水中的Pb2+、Cd2+背景值
Table 2. Experimental factor-level table
表2. 实验因素-水平表

Figure 1. Effects of Pb2+, Cd2+ single and combined stress on the height of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图1. Pb2+、Cd2+单一及复合胁迫对碱蓬株高的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1
(P < 0.05),单一Pb2+、Cd2+胁迫使株高随浓度增加而减小,但500 mg∙kg−1 Pb2+对株高的抑制作用小于200 mg∙kg−1的Pb2+胁迫。幼苗期与花期,不同浓度Cd2+与200 mg∙kg−1的Pb2+复合胁迫,株高均大于相应的单一Pb2+胁迫,表现为拮抗作用,使毒性减小;与500, 20 mg∙kg−1的Pb2+复合胁迫,株高均小于相应的单一Pb2+胁迫,表现为协同作用,其中,500 mg∙kg−1的Pb2+与50 mg·kg−1的Cd2+复合胁迫下株高最小,协同作用显著(P < 0.05),毒性加强,抑制生长,这表明碱蓬的两个生长时期下,Pb2+、Cd2+的相互作用属性不变。重金属影响植物生长,低浓度促进植物生长,高浓度抑制生长,甚至引起植物死亡 [23] [24] 。其中,Pb2+对植物的毒害表现为矮化植物,影响矿质元素的吸收;Cd2+则能与蛋白质相互作用,对细胞产生毒性 [2] 。试验结果中花期对照组株高最高,表明随Pb2+、Cd2+胁迫时间的延长,重金属对碱蓬主要表现为毒害作用,抑制生长。
3.2. Pb2+、Cd2+胁迫对碱蓬SOD活性的影响
SOD活性是植物抗氧化保护系统的第一道防线,主要是清除活性氧,使细胞膜免受氧自由基的伤害,当受到一定范围内的胁迫时,活性会有所提高,增加植物的抗逆能力,其水平高低是衰老与死亡的直观指标 [25] 。由图2可知,幼苗期,单一Cd2+ (5 mg∙kg−1, 50 mg·kg−1)胁迫使SOD活性显著高于对照组,说明一定浓度的Cd2+能诱导SOD合成保护机体;单一Pb2+胁迫以及与低浓度的Cd2+ (5 mg·kg−1, 20 mg·kg−1)复合胁迫使SOD活性随Pb2+浓度增加先升高后降低,峰值分别出现在20 mg·kg−1、200 mg·kg−1的Pb2+处;高浓度的Cd2+ (50 mg·kg−1)胁迫下,SOD活性随Pb2+浓度的增加一直下降。Pb2+与Cd2+复合胁迫,SOD活性均显著小于对应的单一Pb2+胁迫(P < 0.05),说明在复合胁迫中,Cd2+对碱蓬SOD活性影响较大,会阻碍SOD的合成。
花期碱蓬SOD活性约300 U·g−1,极显著低于幼苗期(P < 0.01),表明碱蓬已开始进入衰老期,符合生长周期规律。对照组SOD活性最大(P < 0.05),Pb2+、Cd2+单一及复合胁迫的SOD活性均显著小于对照组(P < 0.05);且SOD活性变化较小;SOD活性在500 mg∙kg−1 Pb2+ + 20 mg·kg−1 Cd2+胁迫下最小。这可能是由于胁迫时间较长,植物吸收累积了一定重金属,阻碍SOD合成;也可能是与植物进入衰老期相关,说明碱蓬在花期自我调节SOD活性来适应外界胁迫的能力有限。
3.3. Pb2+、Cd2+胁迫对碱蓬Pro含量的影响
Pro是植物体内普遍存在的一种逆境保护性物质,有助于抵御逆境胁迫,植物能通过提高其含量调节渗透平衡,保护细胞结构和功能的完整性,其含量在一定程度上反映了植物的抗逆能力 [10] [26] 。由图3
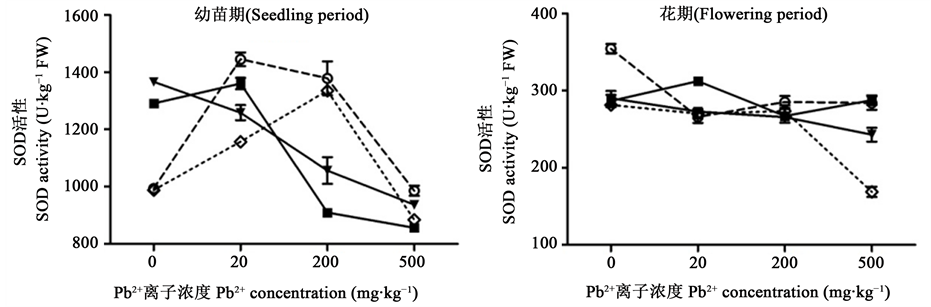
Figure 2. Effects of Pb2+, Cd2+ single and combined stress on the SOD activity of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图2. Pb2+、Cd2+单一及复合胁迫对碱蓬SOD活性的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1

Figure 3. Effects of Pb2+, Cd2+ single and combined stress on the Pro content of Suaeda salsa. Note: Cd2+ 0 mg∙kg−1; Cd2+ 5 mg∙kg−1; Cd2+ 20 mg∙kg−1; Cd2+ 50 mg∙kg−1
图3. Pb2+、Cd2+单一及复合胁迫对碱蓬Pro含量的影响。注:Cd2+ 0 mg∙kg−1;Cd2+ 5 mg∙kg−1;Cd2+ 20 mg∙kg−1;Cd2+ 50 mg∙kg−1
可知,幼苗期,单一Pb2+、Cd2+胁迫使Pro含量随重金属浓度增加先升高后降低;复合胁迫时,Pro含量随Pb2+浓度增加而下降;Pro含量均比对应的单一Pb2+胁迫低(P < 0.05),说明两种重金属复合作用对碱蓬Pro含量影响较大,为协同毒害作用。除200 mg∙kg−1 Pb2+ + 50 mg∙kg−1 Cd2+、500 mg∙kg−1 Pb2+ + 5 mg∙kg−1 Cd2+外,其余浓度复合胁迫时,花期碱蓬Pro含量显著高于对照组(P < 0.05),说明碱蓬能通过提高渗透平衡保护细胞结构和功能完整,有助于抵御逆境胁迫,但在高浓度Pb2+、Cd2+复合胁迫下,Pro的自我调节能力会受到限制。中高浓度Cd2+ (20, 50 mg∙kg−1)胁迫使Pro含量随Pb2+浓度增加而下降;单一Pb2+及Pb2+与低Cd2+ (5 mg∙kg−1)复合胁迫,则使Pro含量随Pb2+浓度增加先升高后下降,峰值均出现在200 mg∙kg−1的Pb2+处,表明碱蓬对外界胁迫响应强烈,这可能与200 mg∙kg−1的单一Pb2+胁迫抑制碱蓬生长相关。
3.4. Pb2+、Cd2+胁迫对碱蓬MDA含量的影响
MDA是植物细胞膜脂过氧化产物之一,能与细胞内各种成分发生反应,引起各种酶和膜的严重损伤,其含量高低可以反应膜脂过氧化作用强弱及质膜损坏程度 [27] 。由图4可知,幼苗期,加入低浓度的Cd2+ (≤20 mg∙kg−1)使碱蓬MDA含量随Pb2+浓度增加而下降,加入高浓度的Cd2+ (50 mg∙kg−1)使碱蓬MDA
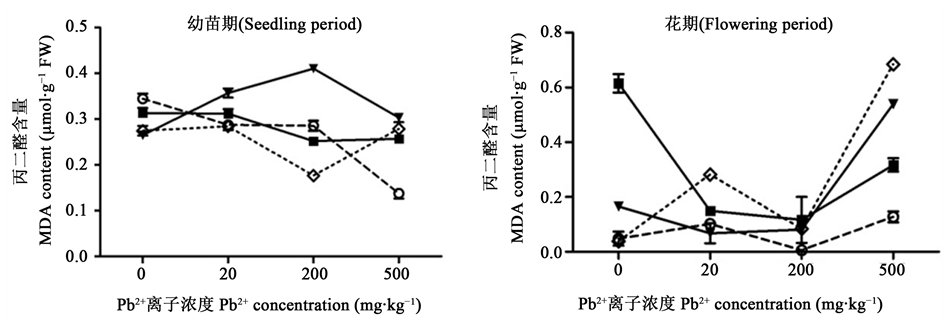
Figure 4. Effects of Pb2+, Cd2+ single and combined stress on the MDA content of Suaeda salsa
图4. Pb2+、Cd2+单一及复合胁迫对碱蓬MDA含量的影响
含量随Pb2+浓度增加先升高后下降。其中,高浓度的Pb2+ (500 mg∙kg−1)与任意浓度的Cd2+复合,MDA含量显著大于单一的Pb2+胁迫(P < 0.05),表明高Pb2+胁迫下,加入不同浓度的Cd2+均能加剧碱蓬质膜损伤;高浓度Cd2+(50 mg∙kg−1)与Pb2+复合的MDA含量高于低Cd2+ (5 mg∙kg−1)与Pb2+复合,说明幼苗期在高浓度的Cd2+胁迫下时,质膜损伤最为严重,在200 mg∙kg−1 Pb2+ + 50 mg∙kg−1 Cd2+时,MDA含量达到最大。花期,碱蓬MDA含量基本高于对照组,500 mg∙kg−1 Pb2+与20 mg∙kg−1 Cd2+复合胁迫下的MDA含量最大;200 mg∙kg−1、500 mg∙kg−1的Pb2+与任意浓度的Cd2+复合,均显著大于对应的单一Pb2+胁迫(P < 0.05),表明在单一和复合胁迫下,质膜都受损严重,加入Cd2+使其损伤更大,表现为协同毒害作用。
4. 讨论
大部分研究表明 [23] [25] [27] (周玉卿等,2012;徐学华等,2009;郭天荣等,2015),低浓度的重金属对植物有促进作用,高浓度则为抑制作用,甚至导致死亡。本试验中,幼苗期碱蓬在单一Cd2+胁迫下符合这一规律;在单一Pb2+胁迫下,低浓度(20 mg·kg−1)的Pb2+促进生长,较高浓度(200 mg·kg−1)的Pb2+抑制生长,更高浓度(500 mg·kg−1)的Pb2+反而促进碱蓬生长,出现异常,这可能是由于碱蓬对高Pb2+存在一种规避机制,与何洁 [28] 研究翅碱蓬对单一铅的累积吸收结果类似;但随碱蓬继续生长,胁迫时间延长,花期碱蓬在各个浓度胁迫下的株高均显著小于对照组(P < 0.05),抑制作用显著,这可能是由于碱蓬体内累积了一定量的重金属,阻碍碱蓬生长。单一胁迫与复合胁迫相较,任意浓度的Cd2+与200 mg·kg−1的Pb2+复合胁迫,株高均显著大于相应的单一Pb2+胁迫(P < 0.05),为拮抗作用;与500、20 mg·kg−1的Pb2+复合胁迫,株高均显著小于相应的单一Pb2+胁迫(P < 0.05),为协同作用,并非两种重金属的复合毒害作用一定强于单一重金属的毒害,这与刘大林 [10] 以Pb2+、Cd2+胁迫饲用高粱、郭天荣 [27] 以Pb2+、Cd2+胁迫空心菜时,复合胁迫较单一胁迫会加剧对植物的毒害作用的结果不同,可能原因是与植物本身的调节方式有关,或许又与重金属在植物体内存在的部位、形式、累积量有一定联系,有待于进一步研究。
逆境胁迫对植物的损伤常常是由于产生过多的体内活性氧(ROS),从而导致膜脂受损,细胞功能丧失。植物往往能有限地在体内合成一些物质来进行自我调节,以适应外界环境 [25] 。SOD活性在重金属胁迫下,往往有3种表现形式 [24] :一是随浓度的增加而增加;二是随浓度增加先升高后下降;三是随浓度的增加而下降,本试验的SOD活性符合后两种情况。单一Pb2+胁迫以及与20 mg∙kg−1的Cd2+复合胁迫,SOD活性均随Pb2+增加先升高后降低,而与5, 50 mg∙kg−1的Cd2+复合胁迫,SOD活性随Pb2+浓度增加而降低,这可能是由于Cd2+浓度的增加使SOD调节机制受损,但其具体受损机理需进一步研究。
幼苗期,碱蓬在单一Pb2+、Cd2+胁迫下,Pro含量随重金属浓度增加先升高后降低;与Cd2+复合胁迫时,Pro含量随Pb2+浓度增加而降低;花期碱蓬的Pro含量基本高于对照组,说明幼苗期碱蓬的Pro调节能力较弱,而在花期Pro调节作用变得较为明显。在单一重金属胁迫时,Pro与SOD相比较,Pro在遇到重金属胁迫时,其响应较慢,在重金属浓度升高后才逐渐产生且产生量较SOD少,因此其调节存在先后,SOD为先,Pro为后。
幼苗期的MDA含量高于花期,随Pb2+浓度增加呈无序变化,与孙永林 [12] 在研究Pb2+对黄瓜的生理影响中,随Pb2+浓度增加MDA含量也增加的结果不同,这可能是碱蓬对Pb2+存在独特的调节机制。
综上所述,幼苗期,SOD活性能迅速对重金属胁迫响应,Pro在花期其渗透调节作用变得较为明显能抵御外界重金属胁迫,MDA含量呈无序变化,代表机体膜质受损状况,其无序变化随外界重金属胁迫及碱蓬体内Pro、SOD活性等组成的调节系统的动态变化而变化。因此,碱蓬在抵御外界重金属胁迫的整个过程中,会启动SOD、Pro两种物质协同抵抗,各自的调节能力有限,且存在先后之分,并能通过MDA含量实时表征植物的受损情况。
5. 结论
(1) Pb2+、Cd2+单一及复合胁迫均对碱蓬有毒害作用,且时间延长毒害作用增加,表现为抑制碱蓬生长;(2) SOD和Pro对重金属调控起重要作用,SOD响应为先,Pro为后;(3) 无论在幼苗期还是花期,当200 mg∙kg−1 Pb2+与20、50 mg∙kg−1 Cd2+复合胁迫时,重金属之间表现为拮抗作用,其他Pb2+浓度与Cd2+复合时均为协同作用。总之,碱蓬对重金属胁迫有独特的调节机制和较强的耐受性,且应对重金属复合胁迫的耐受性更强,因此碱蓬对于滨海滩涂铅、镉重金属污染修复有极大的应用潜力。
基金项目
浙江省自然科学基金项目[The Project Supported by Zhejiang Provincial Natural Science Foundation of China] (LY16C150002);浙江省公益性技术应用研究计划项目[The Research Project on nonprofit technology of Zhejiang Province] (2015C32035);浙江省重中之重学科自设课题[Zhejiang Provincial Top Key Discipling Under Grant] (ZS2016011);浙江万里学院引进人才重点项目[Key project of Zhejiang Wanli University talents] (1741000530);国家级大学生创新创业训练计划项目[National Training Program of Innovation and Entrepreneurship for Undergraduates](201610876028)资助。
文章引用
于晓磊,王路遥,赵欣园,王趁义,滕丽华,杨 佳,胡 杰. 不同生长时期碱蓬在Pb2+、Cd2+单一及复合胁迫下的生理生化响应
Physiological and Biochemical Responses of Suaeda salsa to Pb2+,Cd2+ in Single and Combined Stress at Different Growth Stages[J]. 农业科学, 2017, 07(02): 120-127. http://dx.doi.org/10.12677/HJAS.2017.72015
参考文献 (References)
- 1. 张韵, 蒲新明, 黄丽丽, 等. 我国滨海湿地现状及修复进展[C]//中国环境科学学会. 2013中国环境科学学会学术年会论文集(第六卷), 2013: 5743-4746.
- 2. 刘俊祥, 孙振元, 巨关升, 等. 结缕草对重金属镉的生理响应[J]. 生态学报, 2011, 31(20): 6149-6156.
- 3. 贺靖舒, 丁继军, 潘远智, 等. 石竹(Dianthus chinensis)对土壤镉胁迫的生理生化响应[J]. 四川农业大学学报, 2013, 31(3): 290-295.
- 4. Finger-Teixeira, A., Ferrarese, M.D.L.L., Soares, A.R., et al. (2010) Cadmium-Induced Lignification reStricts Soybean Root Growth. Ecotoxicology & Environmental Safety, 73, 1959-1964. https://doi.org/10.1016/j.ecoenv.2010.08.021
- 5. Gichner, T., Žnidar, I., Száková, J. (2008) Evaluation of DNA Damage and Mutagenicity Induced by Lead in Tobacco Plants. Mutation Research/Genetic Toxicology & Environmental Mutagenesis, 652, 186-190. https://doi.org/10.1016/j.mrgentox.2008.02.009
- 6. Sharma, P., Dubey, R.S. (2005) Lead Toxicity in Plant. Brazilian Journal of Plant Physiology, 17, 35-52. https://doi.org/10.1590/S1677-04202005000100004
- 7. Reddy, A.M., Kumar, S.G., Jyothsnakumari, G., et al. (2005) Lead Induced Changes in Antioxidant Metabolism of Horsegram (Macrotyloma uniflorum, (Lam.) Verdc.) and Bengalgram (Cicer arietinum, L.). Chemosphere, 60, 97-104. https://doi.org/10.1016/j.chemosphere.2004.11.092
- 8. Chen, H.L., Lai, H.Y., Wang, S.M., et al. (2010) Effect of Biosolids and Cd/Pb Interaction on the Growth and Cd Accumulation of Brassica rapa, Grown in Cd-Contaminated Soils. Water Air & Soil Pollution, 206, 385-394. https://doi.org/10.1007/s11270-009-0114-9
- 9. 熊春晖,许晓光,卢永恩, 等. 铅镉复合胁迫下莲藕对铅镉的富集及其生理变化[J]. 园艺学报, 2012, 39(12): 2385- 2394.
- 10. 刘大林, 杨俊俏, 刘兆明, 等. 镉、铅胁迫对草地早熟禾幼苗生理的影响[J]. 草业科学, 2015, 32(2): 224-230.
- 11. 梁烜赫, 曹铁华. 重金属对玉米生长发育及产量的影响[J]. 玉米科学, 2010, 18(4): 86-88.
- 12. 孙永林. 铅对黄瓜叶片丙二醛、脯氨酸和可溶性总糖含量的影响[J]. 长江蔬菜(学术版), 2008, 1(24): 28-30.
- 13. 张爱琴, 庞秋颖, 阎秀峰. 碱蓬属植物耐盐机理研究进展[J]. 生态学报, 2013, 33(12): 3575-3583.
- 14. 李超峰, 葛宝明, 姜森颢, 等. 碱蓬对盐碱及污染土壤生物修复的研究进展[J]. 土壤通报, 2014, 45(4): 1014- 1019.
- 15. 高世珍, 赵兴茹, 崔世茂, 等. 典型持久性有机污染物在翅碱蓬中的分布特征[J]. 环境科学, 2010, 31(10): 2456- 2461.
- 16. 郑春芳, 陈琛, 彭益全, 等. 海水养殖废水灌溉对碱蓬和海蓬子生长和品质的影响[J]. 浙江农业学报, 2012, 24(4): 663-669.
- 17. 李学刚, 袁华茂, 许思思, 等. 胶州湾滨海湿地盐渍土壤中重金属的聚集与分散特性研究[J]. 海洋科学, 2011, 35(7): 88-95.
- 18. 李杏, 项学敏, 周集体, 等. 盐生植物碱蓬在土壤修复及废水处理中的研究现状[J].江苏环境科技, 2007, 20(1): 53-54.
- 19. 张海波, 崔继哲, 曹甜甜, 等. 大豆出苗期和苗期对盐胁迫的响应及耐盐指标评价[J]. 生态学报, 2011, 31(10): 2805-2812.
- 20. Min, Q., Qin, L.N., Liu, Y.J., et al. (2014) The Comparison of Two Methods of Testing Superoxide Dismutase Activity. Journal of Food Safety & Quality, 1, 3318-3323.
- 21. Kusvuran, A. (2015) The Effects of Salt Stress on the Germination and Antioxidative Enzyme Activity of Hungarian vetch (Vicia pannonica Crantz.) Varieties. Legume Research, 38, 51-59. https://doi.org/10.5958/0976-0571.2015.00009.0
- 22. Rady, M.M., Mohamed, G.F., Abdalla, A.M., et al. (2015) Integrated Application of Salicylic Acid and Moringa oleifera Leaf Extract Alleviates the Salt-Induced Adverse Effects in Common Bean Plants/ International Journal of Agricultural Technology, 11, 1595-1614.
- 23. 周玉卿, 赵九洲, 韩玉林. 铅、镉及其复合胁迫对黄菖蒲幼苗生长和生理抗性的影响[J]. 湿地科学, 2012, 10(4): 487-491.
- 24. 温瑀, 穆立蔷. 土壤铅、镉胁迫对4种绿化植物生长、生理及积累特性的影响[J]. 水土保持学报, 2013, 27(5): 234-239.
- 25. 徐学华, 黄大庄, 王连芳, 等. 土壤铅、镉胁迫对红瑞木生长及生理生化特性的影响[J]. 水土保持学报, 2009, 23(1): 213-221.
- 26. 陆海燕, 刘志辉, 吕光辉. 镉污染下芦苇叶片丙二醛、脯氨酸及SOD保护酶反应研究[J]. 干旱区资源与环境, 2013, 27(8): 171-175.
- 27. 郭天荣, 陈丽萍, 冯其芳, 等. 铝、镉胁迫对空心菜生长及抗氧化特性的影响[J]. 核农学报, 2015, 29(3): 571-576.
- 28. 何洁, 陈旭, 王晓庆, 等. 翅碱蓬对滩涂湿地沉积物中重金属Cu, Pb的累积吸收[J]. 大连海洋大学学报, 2012, 27(6): 539-545.
NOTES
*通讯作者。